The Carboniferous timescale: an introduction - Geological Society, London ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
The Carboniferous timescale: an introduction
Spencer G. Lucas1*, Joerg W. Schneider2,3, Svetlana Nikolaeva3,4,5 and
Xiangdong Wang6,7
1
New Mexico Museum of Natural History, 1801 Mountain Road NW, Albuquerque,
NM 87104, USA
2
TU Bergakademie Freiberg, Geological Institute, Bernhard-von-Cotta-Straße 2, 09599
Freiberg, Germany
3
Kazan Federal University, Kremlyovskaya 18, 420 008 Kazan, Republic of Tatarstan,
Russia
4
Natural History Museum, Cromwell Road, London SW7 5BD, UK
5
Borissiak Palaeontological Institute, Profsoyuznaya 123, 117 647 Moscow, Russia
6
Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing
210008, China
7
School of Earth Sciences and Engineering, Nanjing University, Nanjing 210023, China
SGL, 0000-0002-4594-3024
*Correspondence: spencer.lucas@state.nm.us
Abstract: The Carboniferous chronostratigraphic scale consists of two subsystems, six series and seven stages.
Precise numerical age control within the Carboniferous is uneven, and a global magnetic polarity timescale for
the Carboniferous is far from established. Isotope stratigraphy based on Sr, C and O isotopes is at an early stage
but has already identified a few Sr and C isotope events of use to global correlation. Cyclostratigraphy has cre-
ated a workable astrochronology for part of Pennsylvanian time that needs better calibration. Chronostrati-
graphic definitions of most of the seven Carboniferous stages remain unfinished. Future research on the
Carboniferous timescale should focus on Global Stratotype Section and Point (GSSP) selection for the remain-
ing, undefined stage bases, definition and characterization of substages, and further development and integration
of the Carboniferous chronostratigraphic scale with radioisotopic, magnetostratigraphic, chemostratigraphic
and cyclostratigraphic tools for calibration and correlation, and the cross-correlation of non-marine and
marine chronologies.
Today, the Subcommission on Carboniferous definitions of most of the substages used by some
Stratigraphy (SCCS), part of the International workers to subdivide the Carboniferous stages
Commission on Stratigraphy (IUGS), advocates a remain unfinished. For the non-marine Carbonifer-
Carboniferous chronostratigraphic scale of two sub- ous strata, correlations based on palynomorphs,
systems, six series and seven stages (Fig. 1) (Heckel megafossil plants, conchostracans, insects, bivalves
and Clayton 2006). The boundaries of the Carbonif- and tetrapods (amphibians and reptiles) have been
erous System and the bases of three of its seven proposed, but many problems of correlation remain,
stages are defined by global stratotype sections and especially the cross-correlation of Carboniferous
points (GSSPs). The numerical ages of most of non-marine and marine chronologies.
these boundaries appear to have been determined This Special Publication reviews the state of the
with a precision of about 0.3–0.4 myr, but precise art of the Carboniferous timescale, and this introduc-
numerical age control within the Carboniferous is tory chapter provides an overview of this volume. It
generally sparse and uneven (Aretz et al. 2020). A also presents the current Carboniferous timescale of
global polarity timescale for the Carboniferous is the SCCS (Figs 1 & 2).
being developed and is not yet complete. Isotope
stratigraphy based on Sr, C and O isotopes is under
development but has already identified some Sr The Carboniferous Earth, biota and the
and C isotope events of use to global correlation. timescale
Cyclostratigraphy has created an astrochronology
for at least part of Pennsylvanian time that still It is important to understand that the Carboniferous
needs better calibration. Chronostratigraphic timescale is constrained in different ways by the
From: Lucas, S. G., Schneider, J. W., Wang, X. and Nikolaeva, S. (eds) The Carboniferous Timescale.
Geological Society, London, Special Publications, 512,
https://doi.org/10.1144/SP512-2021-160
© 2022 The Author(s). Published by The Geological Society of London. All rights reserved.
For permissions: http://www.geolsoc.org.uk/permissions. Publishing disclaimer: www.geolsoc.org.uk/pub_ethics
Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
from Mississippian volcanic rocks are relatively
few, and little cyclostratigraphy is available to
develop an astrochronology. Nevertheless, the mag-
netic field was active during the Mississippian,
which resulted in a relatively well-understood mag-
netostratigraphy. Thus, Mississippian chronostratig-
raphy based on marine biostratigraphy is relatively
robust and non-provincial, and finds good support
from magnetostratigraphy but little support from
numerical ages and cyclostratigraphy.
The Pennsylvanian amalgamation of Pangaea
closed the Rheic Ocean (Fig. 5). The result was pro-
vincialization of the marine biota, which decreases
the ability to effect biostratigraphic correlations of
broad scope. The tectonics of the amalgamation pro-
duced much volcanism, particularly in the megasu-
tural zone between the supercontinents, so that
more numerical ages are available for Pennsylvanian
time than are available for the Mississippian. How-
ever, the magnetic field activity was much reduced,
when the almost totally reversed polarity Kiaman
superchron effectively eliminated a Pennsylvanian
magnetostratigraphy. The Pennsylvanian world,
however, was a time of great glacial events, so that
cyclostratigraphic data to produce an astrochronol-
ogy are available in the Pangaean tropics for much
of the subsystem.
Thus, in viewing the Carboniferous timescale,
it is fair to say that the global chronostratigraphy
is much stronger for the Mississippian than for
the Pennsylvanian, when correlations between
provinces (particularly between Euramerica, Gond-
wana and Angara) are challenging. The non-
biostratigraphic constraints also differ between the
Mississippian and Pennsylvanian, so that numerical
and other chronological constraints are better devel-
oped for the Pennsylvanian.
During the Mississippian, ammonoids, foramini-
fers, conodonts, corals, brachiopods and echinoderms
are important biostratigraphic guides. In many suc-
cessions, these marine fossils are abundant and
allow the finest subdivision. With a major regression
at the end of the Mississippian, the abundance and fre-
Fig. 1. The Carboniferous chronostratigraphic scale quency of marine groups decreases, even though
showing ratified GSSPs of stage bases. conodonts and foraminiferans remain very important.
Fossil plants are useful for Mississippian biostratigra-
disparate conditions of the Carboniferous Earth dur- phy, but in the Pennsylvanian their proliferation and
ing the Mississippian and the Pennsylvanian (Figs 3– abundance provide a remarkable stratigraphic tool
5). During the Mississippian, Pangaea had not yet for terrestrial sequences. Terrestrial invertebrates
fully assembled, so that a Rheic Ocean separated (especially conchostracans and insects) and tetrapods
Gondwana from the Laurussian supercontinent (footprints and bones) are also very important for
(Fig. 4). Glaciers were limited, so the Mississippian Pennsylvanian non-marine biostratigraphy.
was generally a time of global greenhouse climates
with a relative dearth of volcanism. The marine
biota, able to travel the shelves of the superconti- Carboniferous chronostratigraphy
nents via the Rheic Ocean, was relatively cosmopol-
itan, which results in far-reaching correlations using The Carboniferous chronostratigraphic scale is a
marine biostratigraphy. However, numerical ages hierarchy of two subsystems, six series and seven
Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
Carboniferous timescale
Conodonts Zones European European Main
AGE Epoch/Age
(Stage) Ammonoid Fusulinids& seq.
(Ma) S. China Europe N. America Zones Benthic Forams R T
298 Permian Streptognathodus Streptognathodus Streptognathodus Svetlanoceras
isolatus isolatus isolatus Daixina bosbytauensis -
289.9Ma S.wabaunsensis- S. binodosus Globifusulina robusta
S. wabaunsensis S. fissus S. farmeri
300 S. flexousus
S. tenuialveus S. simplex-S. bellus Daixina sokensis
S. bellus Shumardites /
Gzhelian S. virgilicus S.virgilicus S.virgilicus Vidrioceras Jigulites jigulensis
302 S. vitai
S.vitali
I. nashuiensis S.vitali Rauserites rossicus -
I. simulator I. simulator
I. naraoensis I. simulator S. zethus Rauserites stuckenbergi
304 303.68Ma I. eudoraensis S.firmus I. eudoraensis
I. guizhouensis I.toretzianus S.gracilis Rauserites quasiarcticus
I. magnificus Scancellosus I. confragus
Kasimovian I. cancellosus Dunbarites - Montiparus paramontiparus
I. heckeli / I. turbatus Isagittalis Parashumardites
306 I. turbatus
Sw. makhlinae Sw. makhlinae I. eccentricus Protriticites
Sw. subexcelsa Sw. subexcelsa I. sulciferus pseudomontiparus
307.02Ma Sw. nodocarinata Fusulina cylindrica -
Pennsylvanian
Neognathodus Eoschistoceras Protriticites ovatus
308 N. roundyi
Sw. neoshoensis
roundyi I delicatus Fusulinella bocki
I. acutus Pseudopara- Fusulinella colaniae -
310 I. podolskensis I. podolskensis I. rectus/ legoceras F. voshgalensis -
I. iowaensis Beedeina kamensis
Moscovian Fusulinella subpulchra
312 Mesogondolella Swadelina dissecta I. amplificus/ Paralegoceras /
clarki/ I. obliquus Priscoidella priscoidea
N. bothrops Eowellerites
M. donbassica “Streptognathodus”
314 Diplognathodus
transtivus N. colombiensis Diaboloceras - Aljutovella aljutovica
ellesmerensis Diplognathus Winslowoceras
315.15Ma ellesmerensis /
Declinognathodus Neognathodus Diaboloceras - Verella spicata -
316 donetzianus atokaensis Axinolobus Alj. tikhonovichi
“S.”expansusM2 N. atokaensis
Declinognathodus Branneroceras / Profusulinellarhombiformis
N. nataliae Profusulinella primitiva -
318 marginodosus Gastrioceras
I. klapperi Pseudostaffella gorskyi
Staffellaeformes
Bashkirian “S.”expansusM1 I. sinuosus I. sinuosus Bilinguites / staffellaeformis -
Pseudostaffella praegorskyi
320 Cancelloceras
I. primulus N. askynensis N. bassleri Pseudostaffella antiqua
N. symmetricus N. symmetricus Baschkortoceras /
Id. sinuatus Reticuloceras Semistaffella variabilis -
322 Id. sinuatus Id.sinuatus Semistaffella minuscilari
Homoceras /
D. noduliferuss.l. D. noduliferus D. noduliferus Hudsonoceras Plectostaffellabogdanovkensis
323.4Ma Monotaxinoides
324 G.postbilineatus G. postbilineatus Upper Rhachistognathus
muricatus
transitorius
Lower Rhachistognathus Eumophoceras /
muricatus
G.bollandensis G.bollandensis Cravenoceratoides Eostaffellina protvae
326 Adetognathus
Serpu- unicornis
khovian Cavusgnathus
328 Tumulies / Neoarchaediscus
naviculus
Lochrieaziegleri Cravenoceras postrugosus
Lochriea ziegleri
330 330.34Ma
Upper Gnathodus Lusitanoceras /Lyrogoniatites
Lochrieanodosa Lochrieanodosa bilineatus Arnbergites /Neoglyphoceras Janischewskina typica
332
Gnathodus Goniatites / Howchinia bradyana
334 bilineatus Eoglyphioceras
Gnathodus Lower Gnathodus
bilineatus bilineatus
Neoarchaediscus
336 “Hindeodus” Entogonites
scitulus/
Gnathodus “Apatognathus”
338 praebilineatus / scalenus Pojarkovella nibelis
Visean Lochriea Lochriea commutata
Mississippian
commutata
340
Bollandites /
Bollandoceras Uralodiscus rotundus
342
Gnathodus texanus
Gnathodus
344 Pseudognathodus homopunctatus Planoarchaediscus /
homopunctatus Ammarchaediscus
Eoparastaffella simplex
346
346.73Ma Ammonellipsites / Eoparastaffella morphotype 1
Scaliognathus Gnathodus bulbosus Fascipericyclus Darjella monilis
348 anchoralis / Scalliognathus Eotaphrus burlingtonensis
G. pseudosemiglaber anchoralis - Tetrataxis
Bactrognathus lanei
G. typicus / Doliognatus latus
Doliognathus latus
350 Protognathus Pseudopolygnathus Protocanites / Paraendothyra nalivkini
cordiformis Upper multistriatus Pericyclus
G. typicus / Gnathodus typicus Neopolygnathus carina
G. cuneiformis Spinochernella brencklei
352 G.punctatus Palaeospiroplectammina
G.punctatus
Tournaisian Si. isosticha - Protocanites / tchernyshinensis
Gnathodus
Si. isosticha Upper Si. crenulata delicats Gattendorfia
354
Septabrunsiina minuta
Si. crenulata
Si. crenulata Si. crenulata
356 Si. sandbergi Si. sandbergi Eocanites /
Si. sandbergi
Siphonodella jii Si. jii Si. cooperi Gattendorfia
unilocular interzone
358 Si. duplicata Si. duplicata Si. dupulicata
Si. bransoni Si. bransoni Acutimitoceras
Si. bransoni (Stockumites)
359.3Ma Si. sulcata Si. sulcata/ Protognathodus kockeli Postclymenia Tournayellina pseudobeata
Protognathodus kuehni
360 Devonian Si. praesulcata Si.praesulcata
S. = Streptognathodus, Si.=Siphonodella, Sw.=Swadelina, G.=Gnathodus, F.=Fusulinella, Alj.=Aljutovella, I.=Idiognathodus
Fig. 2. Carboniferous timescale (after Aretz et al. 2020).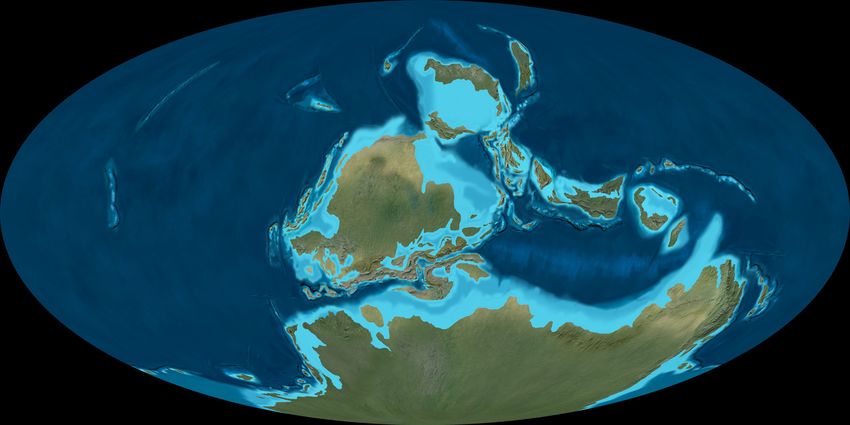
Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
Fig. 3. Major biotic and nonbiotic events of the Carboniferous. D/C, Devonian–Carboniferous; FAD, first
appearance datum.
stages developed over nearly two centuries of Bashkirian and the base of the Asselian. The GSSP
research since Conybeare and Phillips (1822) first of the Visean base has a foraminiferal event as its
used the term Carboniferous as a chronostratigraphic primary signal. Issues in the development of a Car-
construct (Fig. 1). Lucas et al. (2021a) review the boniferous chronostratigraphic scale include the
nearly two-century-long development of the Carbon- rank of chronostratigraphic units, provinciality,
iferous chronostratigraphic scale. Carboniferous conodont biostratigraphy, palaeobotanical biostra-
stage nomenclature developed with the proposal of tigraphy, and the development of astrochronology
numerous regional stages/substages based primarily and other methods of chronology and correlation
on palaeobotanical, foraminiferal and ammonoid (Lucas 2021a).
biostratigraphy, especially in western Europe, the Ratified GSSPs define boundaries of three of the
former Soviet Union, China and the USA. From seven Carboniferous stages recognized by the SPS,
the regional stages, seven ‘global stages’ have been and also define the boundaries of the two Carbonif-
identified (in ascending order): Tournaisian, Visean, erous subsystems and of the Carboniferous System
Serpukhovian, Bashkirian, Moscovian, Kasimovian (Fig. 1). The bases of most of the Carboniferous sub-
and Gzhelian. Three of the four ratified GSSPs rele- stages (Fig. 6) lack formal definition. They provide a
vant to the Carboniferous chronostratigraphy use more refined subdivision of Carboniferous time than
conodont evolutionary events as the primary signal do the stages and should be the focus of much future
for correlation – the bases of the Tournaisian and chronostratigraphic research.
Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
Fig. 4. Mississippian palaeogeography. © 2016 Colorado Plateau Geosystems Inc. Used by licence.
One of the most detailed schemes of Carboni- microfossils are possible between these regions,
ferous chronostratigraphy is that developed in the and all of the regional schemes are correlated to the
former Soviet Union (Fig. 6). Alekseev et al. official Russian General Stratigraphic scheme for
(2021) review this scheme by presenting the Carbon- the Carboniferous using zonations based on index
iferous stratigraphy and chronostratigraphy officially fossils. The Russian General Stratigraphic scheme
adopted in regions of the Russian Federation. These is correlated to the International Stratigraphic Scale
regions include the Moscow Basin/the Urals, North based on ammonoids, conodonts, foraminiferans
Timan, Siberia, Taimyr, the Kuznetsk Basin, the and palynomorphs. It has provided most of the stan-
Mongolo-Okhotsk Region, Omolon and the Ver- dard stages of the Carboniferous chronostratigraphic
khoyansk–Kolyma Region, and encompass different scale (Fig. 1).
geological histories and distinct depositional set- Perhaps more than any interval of the Phanero-
tings. Broad correlations based on macro- and zoic (except for the late Cenozoic), the
Fig. 5. Pennsylvanian palaeogeography. © 2016 Colorado Plateau Geosystems Inc. Used by licence.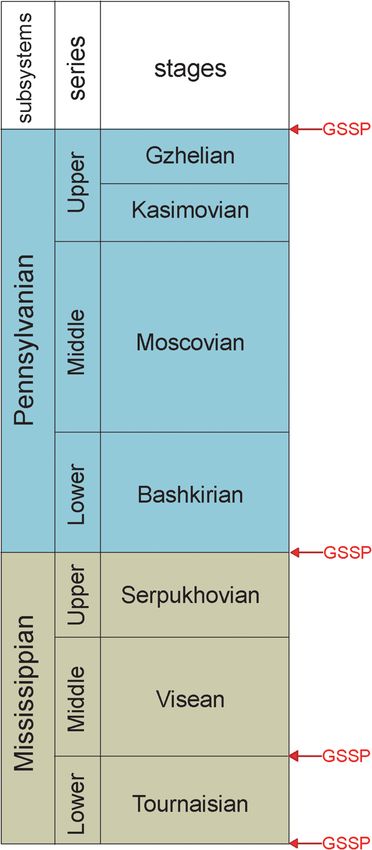
Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
Fig. 6. Carboniferous regional substages.
provincialization of the Carboniferous (especially is currently defined (also see González 2005).
Pennsylvanian) biota hinders global biostratigraphic Based on the abundant palaeontological record of
correlations and the development of a single, glob- the upper Paleozoic deposits of central-western
ally applicable chronostratigraphy. This problem is Argentina, central Patagonia and eastern Argentina,
especially evident in parts of Gondwana. González González and Díaz Saravia (2021) propose
and Díaz Saravia (2021) review the Carboniferous five successive regional stages: Malimanian (late
and earliest Permian rocks in the western Andean Tournaisian), Barrealian (mid-Carboniferous or Ser-
belt of Argentina, which contain a record so exten- pukhovian–Bashkirian), Aguanegrian (Late Penn-
sive that it allows a detailed reconstruction of the his- sylvanian), Uspallatian (Asselian–Tastubian?) and
tory of the late Paleozoic ice ages along the Bonetian (Sakmarian).
southwestern margin of South American Gondwana.
Severe endemism of the Gondwana biota during this
time interval makes it difficult to achieve a precise Radioisotopic ages
correlation of these glacially influenced deposits
with coeval strata of the palaeoequatorial belt, Aretz et al. (2020) recently presented a Carbonifer-
where the global Carboniferous chronostratigraphy ous numerical timescale based on the constraintsDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
provided by 47 radioisotopic ages to which they type palaeomagnetic studies, allowing the identifica-
assigned a 0.4 myr uncertainty. By adding Permian tion of polarity-bias chrons. Seven polarity bias
numerical ages to the analysis, a total of 84 high- chrons exist in the Mississippian (MI1nB–MI4nB),
resolution U/Pb ages are available, with the majority with an additional 33 conventional magnetochrons
having an uncertainty of 0.3–0.4 myr. This dataset is and submagnetochrons (MI4r–MI9r). The Kiaman
based mainly on GTS2012, Davydov et al. (2010, Superchron begins in the mid-Bashkirian, with data
2012), Schmitz and Davydov (2012) and Pointon indicating some brief normal polarity submagneto-
et al. (2012, 2014, 2018, 2019), as well as on Jirásek chrons within the Superchron. The Moscovian and
et al. (2013, 2018). Numerical age constraints are Gzhelian polarity is best resolved in magnetostrati-
weakest for the Mississippian stages, Tournaisian graphic studies from the Donets Basin and the south-
and Visean, which are the longest Carboniferous ern Urals. An assessment of supporting data from
stages. Exceptions are the radioisotopic ages from palaeopole-type studies suggests that the Ukrai-
the Central European Variscides presented by Poin- nian/Russian datasets currently provide the best
ton et al. in a series of articles from 2012 to 2019 (see magnetic polarity data through the Pennsylvanian.
above). Polarity bias assessment indicates a normal polarity
With the increased tectonism during the final clo- bias zone in the Kasimovian. In the Pennsylvanian
sure of the Rheic Ocean by continent–continent col- there are 27 conventional magnetochrons and sub-
lision, volcanism increased considerably, so the magnetochrons (PE1n–CI1r) and one normal polar-
chance of finding ash beds suitable for dating aug- ity bias chron (PE8nB).
mented. The dataset presented by Aretz et al.
(2020) presents representative ages for marine Car-
boniferous deposits. Further radioisotopic ages Isotope stratigraphy
from mixed marine–continental (paralic) and pure
continental deposits in relation to marine–non- The use of strontium, carbon and oxygen isotopes in
marine biostratigraphic correlations are discussed stratigraphic correlation (‘chemostratigraphy’) has
in detail by Schneider et al. (2020) and in this vol- grown dramatically during the last decade. We note
ume. A wealth of radioisotopic ages of non-marine that isotope curves that plot the composition of an
Carboniferous deposits has recently been produced element or changes in the ratio of isotopes of an ele-
for the Czech basins as a standard for central Europe ment have the potential to provide a means of corre-
by the team of Opluštil et al. (2016a, b) and in this lation essentially independent of other methods.
volume. A summary of non-marine–marine correla- However, like magnetostratigraphy, this record
tions based on biostratigraphy and radioisotopic needs calibration to a datum or to datums, either bio-
ages, as well as on magnetostratigraphy, for the stratigraphic or radioisotopic.
Pennsylvanian and Permian of Euramerica, includ- Chen et al. (2021) present an updated set of
ing the Russian Platform and the Karoo Basin of Carboniferous Sr, C and O isotope stratigraphies
Gondwana, is shown here in Figure 7 (an improved based on the existing literature. The Carboniferous
version of fig. 2 in Schneider et al. 2020). 87
Sr/ 86Sr record, constructed using brachiopods and
conodonts, identifies five phases beginning with a
rapid decline from a peak value of c. 0.70840 at
Magnetostratigraphy the Devonian–Carboniferous boundary to a trough
(0.70776–0.70771) in the Visean followed by a
The global polarity timescale for rocks of Late Juras- rise to a plateau (c. 0.70827) in the upper Bashkirian.
sic, Cretaceous and Cenozoic age provides a valu- A decline to c. 0.70804 extends from the earliest
able tool for evaluating and refining correlations Gzhelian to the end of the Carboniferous.
that are based primarily on radioisotopic ages or bio- Contemporaneous carbonate δ13C records show
stratigraphy. Carboniferous magnetostratigraphy has considerable variability between the materials
long been thought to consist of a mixed polarity analysed and between regions, although a few pro-
interval of Tournaisian–Bashkirian age, followed nounced excursions (e.g. a mid-Tournaisian positive
by the Kiaman Reversed Polarity Superchron, excursion and an end-Kasimovian negative excur-
which lasted from the mid-Bashkirian through the sion) can be identified in most records. Bulk carbon-
early part of the middle Permian (about 50 myr). ate δ13C records from Europe and South China are
However, there is no agreed geomagnetic polarity generally consistent with those of brachiopod calcite
timescale for the Carboniferous. from North America in terms of both absolute values
Hounslow (2021) notes that the geomagnetic and trends.
polarity pattern for the Carboniferous is incom- Both brachiopod calcite and conodont phosphate
pletely known and best resolved in the Serpukhovian δ18O have a large regional variability, so that Car-
and Bashkirian. In the Tournaisian–mid-Visean, boniferous δ18O records cannot be used for precise
interval polarity is mainly derived from palaeopole- stratigraphic correlation. Nevertheless, significantDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
Fig. 7. Multistratigraphic correlations of the basins discussed by Schneider et al. (2020). Positions of the radioisotopic ages are indicated by stars. For the data used for the
correlations, the dating methods, error ranges of the radioisotopic ages and for discussion, see the contributions by Schneider et al. (2020, 2021). Marine deposits are marked in
blue. Abbreviations: NA, North American regional scale; WE, West European regional scale; Miss., Missourian; Road., Roadian; Gr., Griesbachian; Di., Dienerian; Sm.,
Smithian; Sp., Spathian; Cant., Cantabrian; Graiss., Graissessac; Cgl., conglomerate; Kreuzn., Kreuznach.Downloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
positive δ18O shifts during some intervals (the mid- Non-fusuline foraminiferans
Tournaisian and the Mississippian–Pennsylvanian
transition) can be used for broad, global correlation. Non-fusuline foraminiferans are abundant in many
Carboniferous marine strata, and some taxa have
very broad distributions in shallow-marine carbonate
facies. This has led to the use of non-fusuline forami-
Cyclostratigraphy niferans in Carboniferous biostratigraphy, especially
in Europe, North America and Asia. Indeed, one of
The late Paleozoic ice ages began during the Late the few ratified Carboniferous GSSPs, the base of
Devonian and continued into the early Permian, the Visean, has a non-fusuline foraminiferan biotic
and were the longest-lived (c. 370–260 Ma) and event as its primary signal (Devuyst et al. 2003;
possibly most extensive of the icehouse periods of Richards and Aretz 2009).
the Phanerozoic. Montañez (2021) notes that mid- Vachard and Le Coze (2021) review the biostra-
to high-latitude glaciogenic deposits of the Carbonif- tigraphy of Carboniferous smaller foraminifers,
erous record a complex and dynamic history, with which include representatives of the classes Fusuli-
ice waxing and waning from multiple ice centres, nata, Miliolata and Nodosariata. The main biostrati-
and transcontinental ice sheets possibly present graphic markers belong to the superfamilies
during the apex of glaciation. New high-precision Archaediscoidea, Lasiodiscoidea and Bradyinoidea,
U–Pb ages confirm a previously hypothesized west and secondary biostratigraphic markers belong to
to east progression of glaciation during the late Lituotubelloidea (=‘Tournayelloidea’), Endothyroi-
Paleozoic icehouse and indicate that its demise dea and Loeblichioidea (the latter gave rise to the
occurred as a series of synchronous and widespread primitive Fusulinida). The Miliolata appeared during
deglaciations. The glaciation history of the Carbonif- the Visean–Serpukhovian boundary interval, and the
erous is also archived by far-field effects (primarily typical Carboniferous miliolates are primitive nube-
sea-level changes) in the low-latitude stratigraphic culariins and cornuspirinins. Tubiphytids might be
record, similar to the Cenozoic icehouse. However, miliolate and cyanobacterium consortia, derived
further evaluation of the phasing between climatic, from the nubeculariin Palaeonubecularia. The most
oceanographic and biotic changes during the ice- primitive nodosariates (syzraniids) appeared in the
house requires additional chronostratigraphic Moscovian, and in the latest Carboniferous they
constraints. gave rise to the Protonodosaria, Nodosinelloides,
The far-field effects of the late Paleozoic ice ages and possibly Polarisella, Paravervilleina and the
allow constructions of a ‘floating’ astrochronology oldest Geinitzinoidea. Palaeobiogeographical distri-
for much of the Pennsylvanian (Hinnov and Ogg butions of Pojarkovella, Janischewskina, Eosigmoi-
2007). This astrochronology provides a high tempo- lina, Brenckleina, Spireitlina, Hemigordius and
ral resolution of part of the late Paleozoic record, Syzrania document successive Carboniferous fora-
well demonstrated in both deep- and quiet-water miniferal migrations between the Palaeotethys,
deposits (e.g. Heckel 2013). Nevertheless, rigorous Ural and Panthalassan oceans.
testing of astronomical forcing in low-latitude cyclo-
themic successions, which have a direct link to
higher-latitude glaciogenic records through inferred Fusulines
glacioeustasy, will require a comprehensive
Fusuline foraminiferans have long played an impor-
approach that integrates new techniques with addi-
tant role in Pennsylvanian biostratigraphy and chro-
tional independent age constraints.
nostratigraphic definitions (e.g. Douglass 1977).
Indeed, the very first published fusuline-based corre-
lation was between Carboniferous fusulines in
Marine biostratigraphy Russia and the USA (Verneuil 1846). In general,
Pennsylvanian fusuline genera are of global distribu-
The distribution of fossils in marine strata (biostra- tion, whereas the species are restricted to Pennsylva-
tigraphy) has long provided the primary basis for nian provinces, so that fusuline correlations vary in
construction of the Carboniferous chronostrati- detail and precision.
graphic scale. The most important taxa in this regard Ueno (2021) reviews Carboniferous fusuline bio-
are non-fusulinid foraminifers, fusulines, ammo- stratigraphy by presenting a synthesis of the taxon-
noids and conodonts. Brachiopods and rugose corals omy, phylogeny, palaeogeographical distribution,
have also provided important biostratigraphy of regional biostratigraphy and palaeobiogeography.
Carboniferous marine strata. These groups are The Carboniferous fusulines are assigned to the fam-
reviewed by articles in this volume, as are the bio- ilies Ozawainellidae, Staffellidae, Schubertellidae,
stratigraphic utility of Carboniferous crinoids and Fusulinidae and Schwagerinidae, of which 95 genera
marine bivalves. are considered taxonomically valid. FusulinesDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
appeared in the latest Tournaisian, and during the characterized mostly by endemic taxa, which ham-
Mississippian they were of small size and morpho- pers long-distance correlation. An exception is the
logically conservative. rapid diversification of taxa of the Choristitinae,
During the Pennsylvanian, fusulines became which were widespread from the Bashkirian to the
larger and more diversified to become abundant in Moscovian, allowing long-distance correlation.
many Pennsylvanian microfossil assemblages. Ueno
(2021) reviews regional fusuline successions in 39 Crinoids
provinces, which provide a refined biostratigraphy
that enables zonation and correlation with substage- During the Carboniferous, crinoids were common
or higher-resolution precision in the Pennsylvanian. and often so locally abundant that their skeletal ossi-
Fusulines had a cosmopolitan palaeobiogeographi- cles formed limestones termed encrinites. Ausich
cal distribution in Mississippian time, suggesting et al. (2021), in their review of the Carboniferous
unrestricted faunal exchange through the palaeoe- record of crinoids, note that both the Mississippian
quatorial Rheic Ocean. However, during the Penn- and the Pennsylvanian started with high rates of cri-
sylvanian, after the formation of Pangaea, fusulines noid evolution and ended with low evolutionary
started to show provincialism. Their distributions rates associated with glaciation. Paleozoic crinoid
define the Ural–Arctic region in the Boreal realm, biodiversity reached its maximum during the Car-
the Paleotethys, Panthalassa and North American boniferous, from which there are numerous well-
Craton regions in the Palaeoequatorial realm, and documented localities with high biodiversity.
the Western Gondwana and the Eastern Peri- According to Ausich et al. (2021), crown-based cri-
Gondwana regions in the Gondwana realm. noid genera can be employed as biostratigraphic
indicators of Carboniferous stages. For Mississip-
Brachiopods pian crinoids, 37 genera are designated as biostrati-
graphically useful; and for the Pennsylvanian, 44
Brachiopoda is a phylum of marine animals with two genera are thus identified. Recognition of the utility
valves known from more than 12 000 fossil species of these genera for biostratigraphy is important for
in more than 5000 genera. Brachiopods were com- dating crinoidal deposits, which may be devoid of
mon shelly benthos during the Carboniferous, mostly other biostratigraphically useful fossils, and this
as seafloor filter feeders. adds to our overall ability to resolve Carboniferous
In their review of Carboniferous brachiopod marine biostratigraphy and correlation.
biostratigraphy, Angiolini et al. (2021) stress how
difficult it is to establish a biochronological scheme Rugose corals
for global correlation based on brachiopods because
of provincialism and endemism. However, numer- Rugose corals were one of the major fossil groups
ous new brachiopod assemblages have been in shallow-marine Carboniferous environments,
described during the last 40 years, making it possible and have long played an important role in the subdi-
to improve and update the brachiopod biostratigra- vision and correlation of Carboniferous strata. Thus,
phy in different regions. during the last century, biostratigraphic schemes
Thus, Angiolini et al. (2021) evaluate the bio- were established, and extensive taxonomic works
stratigraphic significance of the most important on rugose corals have been undertaken. Wang
brachiopod taxa to present correlations in seven geo- et al. (2021) review the Carboniferous record of
graphical regions. The Mississippian is characterized rugose corals to clarify their composition and
by rich brachiopod faunas that include widespread biostratigraphy.
taxa with a good potential for global correlation, This review documents two major evolutionary
such as Antiquatonia, Buxtonia, Delepinea, events in the Carboniferous rugose corals: (1) after
Fluctuaria, Lamellosathyris, Marginatia, Ovatia, the Devonian extinctions, the Tournaisian recovery
Rhipidomella, Rugosochonetes, Spinocarinifera, event, with abundant records of typical Carbonifer-
Syringothyris, Tylothyris and Unispirifer. From the ous rugose corals such as columellate taxa and a
mid-Visean to the late Serpukhovian, taxa of gigan- significant diversification of large, dissepimented
toproductidines are biostratigraphically significant, corals; and (2) the changeover of rugose coral com-
and are present everywhere except South America position at the mid-Carboniferous boundary, which
and Australia, which remain as distinct faunal prov- encompassed the disappearance of large dissepi-
inces for most of the Carboniferous. A major turn- mented taxa with complex axial structures and the
over in brachiopods took place at the beginning of appearance of typical Pennsylvanian compound
the Pennsylvanian, and ushered in a higher degree rugose taxa, the Petalaxidae, Geyerophyllidae and
of provincialism. Pennsylvanian brachiopod faunas Waagenophyllida. The Mississippian successions
are diverse in China, Russia and North America, of rugose corals have higher temporal resolution
but elsewhere they are less developed and are than the Pennsylvanian ones, which is likely to beDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
due to the late Paleozoic ice ages that resulted in the foraminiferal and conodont biostratigraphic scales
lack of continuous Pennsylvanian strata in many for most of the Carboniferous.
regions, especially in Europe and North America. The Carboniferous ammonoid genozones, with a
Rugose corals are totally missing in the Pennsylva- few gaps, are now recognized throughout the entire
nian of Gondwana. To achieve a high-resolution system in most successions worldwide. Thus, from
biostratigraphy of rugose corals, more detailed taxo- 10 to 11 ammonoid genozones are now identified
nomic work and precise correlations between differ- in the Mississippian, and eight to nine genozones
ent fossil groups are needed: for example, correlative are recognized in the Pennsylvanian. Based on these,
studies that more closely link the biostratigraphy of the lower boundaries of the Carboniferous subsys-
foraminifers to that of rugose corals. tems are reasonably well correlated with the ammo-
noid zonation, whereas correlations with the ratified
foraminiferan-based lower boundary of the Visean
Marine bivalves and other stage boundaries need additional research.
Future success in the application of ammonoids to
The bivalves were, like the brachiopods, common
Carboniferous biostratigraphy and chronostratigra-
denizens of Carboniferous seafloors. However,
phy will also depend on accurate identification and
they have only been used in a limited fashion in Car-
re-illustration of the type material of many taxa,
boniferous biostratigraphy. Amler and Silantiev
including material described by the pioneers of Car-
(2021) summarize research on the biostratigraphic
boniferous ammonoid biostratigraphy.
application of Carboniferous marine bivalves world-
wide. The role of marine bivalves in Carboniferous
stratigraphy, with a focus on the South Laurussian Conodonts
margin and the Palaeotethys, is outlined. Although
Conodonts are microscopic, tooth-like structures
marine bivalves have not received primary attention
composed of calcium phosphate that are abundant
for biostratigraphic purposes, a wealth of data exists
and widespread in Carboniferous marine strata.
nearly worldwide to complement the more favoured
Although the biological source of conodonts was
brachiopod, foraminiferan, ammonoid and conodont
long unknown, they are now clearly associated
biozonations.
with chordates. In the 1980s, conodonts began to
Pelagic, open-marine bivalves with a basin-wide
be used for defining Carboniferous chronostrati-
distribution are well suited for biostratigraphy,
graphic boundaries (e.g. Paproth et al. 1980). Thus,
whereas inner-shelf or nearshore groups are mostly
the GSSPs of three stages (bases of the Tournaisian,
rather restricted to specific facies types (substrates).
Bashkirian and Asselian) relevant to the Carbonifer-
According to Amler and Silantiev (2021), based
ous chronostratigraphic scale have conodont biotoic
on the current fossil record, the vertical distribution
events as primary signals, and it is likely that the
of marine bivalves in pelagic facies in the western
other Carboniferous stage bases awaiting GSSP def-
Palaeotethys provides useful biostratigraphy from
inition will also use conodont events as their primary
the Middle Famennian (Hembergian Stage) to the
correlation signals (Aretz et al. 2020; but see Lucas
Early Tournaisian (Balvian = Gattendorfian Stage)
2021a for a critique of the use of conodonts in Car-
and from the Visean (Aprathian = Goniatites Stage)
boniferous GSSP definitions).
to the Serpukhovian (Pendleian–Arnsbergian =
Barrick et al. (2021) review the state of the art of
Eumorphoceras Stage). The Middle and Late Tour-
Carboniferous conodont biostratigraphy, which con-
naisian (Erdbachian = Pericyclus Stage) are gener-
sists of regional zonations that reflect the palaeogeo-
ally poor in bivalve fossils.
graphical distribution of taxa and distinct shallow-
and deep-water conodont biofacies. Nevertheless,
Ammonoids some species have a global distribution and can be
used in high-resolution correlations. These taxa are
Ammonoids have long been important to Carbonif- incorporated into definitions of global Carboniferous
erous marine biostratigraphy, and Nikolaeva chronostratigraphic units, but a standard global Car-
(2021) reviews the application of ammonoids to Car- boniferous zonation has not been achieved.
boniferous chronostratigraphy. Considerable pro- The lowermost Mississippian is zoned by
gress has been made in refining the traditional Siphonodella species, except in shallow-water facies
ammonoid zonation that has been a cornerstone of where other polygnathids are used. Gnathodus spe-
Carboniferous biostratigraphy and chronostratigra- cies diversified during the Tournaisian and are used
phy. Thus, refined collecting and documentation of to define many Mississippian zones. A late Tournai-
occurrences in western Europe, North Africa, the sian maximum in diversity, characterized by short-
Urals, China and North America have established lived genera, was followed by lower-diversity faunas
the first evolutionary occurrences of many ammo- of Gnathodus species and carminate genera through
noid taxa, and facilitated their correlation with the Visean and Serpukhovian. By the late Visean andDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
Serpukhovian, species of Lochriea provide the best tetrapod (amphibian and reptile) footprints and
biostratigraphic resolution, and shallow-water zona- body fossils. Particularly significant is the role that
tions based on Cavusgnathus and Mestognathus spe- megafossil plants have played in Carboniferous bio-
cies are difficult to correlate. stratigraphy and chronostratigraphy. Studies of the
An extinction event near the base of the Pennsyl- fossil floras of the ‘coal forests’ began in Europe in
vanian was followed by the appearance of the new the early 1800s and soon thereafter in North America
gnathodid genera Declinognathodus, Idiognatho- (e.g. von Schlotheim 1804; Brongniart 1821; Stern-
dus, Idiognathoides, Neognathodus and Rhachistog- berg 1821). Extensive palaeobotanical biostrati-
nathus. By the middle of the Moscovian, a few graphic schemes developed, and many important
genera remained, namely Idiognathodus, Neogna- Carboniferous chronostratigraphic constructs, such
thodus and Swadelina. During the middle Kasimo- as Namurian, Westphalian and Stephanian, are
vian and Gzhelian, only Idiognathodus and rooted in palaeobotanical biostratigraphy.
Streptognathodus species were common. Near the
end of the Gzhelian, a rediversification of Streptog- Palynomorphs
nathodus species extended into the Cisuralian.
Spores and pollen are the microscopic reproductive
structures of vascular plants. Their organic walls
Chondrichthyans resist pressure, desiccation and microbial decompo-
Ginter (2021) notes that among the groups of marine sition, so they are often well preserved in sedimen-
fishes that existed during the Carboniferous, the tary rocks, and Carboniferous strata are no
Chondrichthyes appear to have the greatest biostrati- exception. Because of their abundance (one plant
graphic potential because most taxa are readily may produce thousands of palynomorphs), durabil-
identified by isolated teeth, which can be locally ity and easy dispersal (often by wind), palynomorphs
abundant. However, despite the long history of are found in both non-marine and marine strata.
study of Paleozoic sharks, and especially their Thereby, they provide an important means for cross-
teeth, our knowledge of their usefulness in biostra- correlation of non-marine and marine strata based on
tigraphy and palaeoecology is still at an early stage shared palynomorph taxa. However, most palyno-
of development. This is mainly because palaeoich- morphs are only dispersed within a few kilometres
thyologists have long focused on descriptions of or less of the plant that produced them, and any pro-
individual taxa, and not on the documentation of vincialization of the palaeoflora hinders their use in
whole assemblages. According to Ginter (2021), broad-scale correlation. Furthermore, plants are
the microscopic teeth of pelagic stem-group Chon- very environmentally sensitive, so palaeoenviron-
drichthyes, such as Thrinacodus (Phoebodonti- mental and facies restrictions of extinct plants can
formes), Denaea and Stethacanthulus (Falcatidae, affect the distribution of their palynomorphs.
Symmoriiformes) appear to be more useful than Eble (2021) presents a summary of palynological
macrofossils (e.g. tooth plates of Holocephali) data for Pennsylvanian-age coal beds in the Appala-
because of their wider geographical distribution chian Basin of the eastern USA, discussed primarily
and lesser facies dependence. from a biostratigraphic perspective. Coal-bed paly-
However, Ginter’s (2021) review fails to recog- nofloras of Early Pennsylvanian–early Permian age
nize some important Carboniferous selachian assem- are compared and correlated with miospore assem-
blages that are directly associated with fusulinid blage zones long established in western Europe
and/or conodont age control. A good example is (e.g. Clayton et al. 1977). Coal beds in the Appala-
the Kinney Brick Quarry in New Mexico, USA, chian Basin are Early–Late Pennsylvanian in age,
which has a diverse chondrichthyan fossil assem- with some early Permian coals located in the north-
blage (Hodnett and Lucas 2021; Hodnett et al. ern part of the basin. Palynological analyses of
2021), and direct age control as early Missourian these coals provide evidence of Pennsylvanian wet-
(Kasimovian) based on fusulinids and conodonts land palynofloras changing in composition through
(Lucas et al. 2011; Rosscoe and Barrick 2021). time. In addition, the occurrence and range of
Such assemblages are necessary to form the back- selected palynotaxa allow comparison and correla-
bone of a useful Carboniferous chondrichthyan tion with miospore assemblage zonations developed
biostratigraphy. for coal-forming basins in the interior region of the
USA, and also with western Europe.
Non-marine biostratigraphy Megafossil plants
Non-marine Carboniferous biostratigraphy has also As noted above, megafossil plants have been part of
been developed, based primarily on palynomorphs, Carboniferous chronostratigraphy and biostratigra-
megafossil plants, conchostracans and insects, and phy back to the seminal European works on theDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
Introduction: the Carboniferous timescale
flora of the ‘coal forests’ of the early 1800s. This is a potential (e.g. Scholze and Schneider 2015;
flora dominated by primitive conifers, lycopsids, pel- Schneider and Scholze 2018).
tasperms, true ferns, sphenopsids and cordaites. Mississippian occurrences of fossil insects are
Following the terrestrialization of the global biota very rare and not of biostratigraphic value. The
that began during the Devonian (Isozaki and Servais oldest known winged insect is Delitzschala Brauck-
2017), the Carboniferous terrestrial vegetation mann and Schneider (1996) from the Upper Missis-
became widespread, diverse and abundant. Accord- sippian (upper Serpukhovian, Arnsbergian) of
ing to the review by Opluštil et al. (2021), the result- Germany. The entomofauna of Guandacol, Argen-
ing fossil record has proved to be an effective tina, also has winged insects (Petrulevičius and
biostratigraphic tool for intra- and interbasinal corre- Gutiérrez 2016) and apparently is a bit younger but
lations in the palaeoequatorial Euramerican province not well dated. The advanced evolutionary stage of
that extended from the current locations of the North these oldest known winged insects indicates a
American Midcontinent basin to the Variscan basins much earlier origin, in the Devonian. Insects gain
of the Czech Republic. In addition to palaeogeogra- biostratigraphic importance through the sudden and
phy, Carboniferous plant biostratigraphy is strongly widespread appearance of cockroachoids (order
affected by a transition from greenhouse conditions Blattodea) during the Early Pennsylvanian (middle
during most of the Mississippian to an icehouse cli- Bashkirian), to become common in continental,
mate in the Pennsylvanian. mixed continental–marine and also in marine depos-
The Mississippian climate resulted in weak pro- its (e.g. Schneider 1983; Ricetti et al. 2018; Belah-
vincialism, with a cosmopolitan flora ranging from mira et al. 2019; Trümper et al. 2020).
the tropics to middle latitudes. The global cooling Schneider et al. (2021) present a biostratigraphy
around the Mississippian–Pennsylvanian boundary based on conchostracans and insects of mixed conti-
enhanced development of a latitudinal climatic zona- nental–marine and purely non-marine sections in the
tion and related floral provincialism. These changes palaeotropical belt of the Euramerican biotic prov-
are expressed in the recognition of distinct realms ince. They recognize nine insect and eight conchos-
or kingdoms, by which the tropical Amerosinian tracan zones that are either newly defined or
Realm (or Euramerican and Cathaysian realms) is improved. These zones encompass the time interval
surrounded by the Angaran and Gondwanan realms from the Early Pennsylvanian (middle Bashkirian)
occupying middle–high latitudes of the northern into the early Permian (early Asselian). They are
and southern hemispheres, respectively (Fig. 5). Flo- linked to the marine Standard Global Chronostrati-
ristic endemism in the Pennsylvanian thus precludes graphic Scale by common occurrences of insects
development of a global macrofloral biostratigraphy. and/or conchostracans with conodonts in mixed
Instead, each realm or area has its own biostrati- marine–continental sections, as well as by available
graphic scheme. The less rich and less diverse floras and reliable radioisotopic ages of associated volcanic
of the Gondwanan and Angaran realms only support rocks and ash beds. This insect and conchostracan
relatively low-resolution macrofloral biostratigra- zonation is an alternative tool to the well-established
phy. Higher-resolution macrofloral zonations exist megafossil plant biostratigraphy of the Pennsylva-
only in the tropical Amerosinian Realm due to diverse nian. In contrast to the latter, only single specimens
and abundant floras dominated by free-sporing and of insects or conchostracans allow biostratigraphic
early seed plants that lived in extensive wetlands. ages to be established with a similar high temporal
resolution.
Conchostracans and insects
Non-marine bivalves
Conchostracans are bivalved crustaceans that have
lived in freshwater lakes and ponds for the last few Non-marine bivalves are locally common in Carbon-
hundred million years. Their minute, drought- iferous strata, but they have only seen limited use in
resistant eggs can be dispersed by winds, and this Carboniferous biostratigraphy. Amler and Silantiev
guaranteed a broad geographical range to some (2021) summarize research on the biostratigraphic
conchostracan taxa across much of Carboniferous application of Carboniferous non-marine bivalves
Pangaea. Their habitats ranged from perennial worldwide.
lakes to seasonal playa lakes and temporary ponds Non-marine bivalves were of biostratigraphic
and puddles, where they could form mass death interest in the early decades of stratigraphic research,
assemblages. This, together with relatively high spe- in Europe, Asia and North America (i.e. mostly
ciation rates, make them ideal guide fossils, espe- Laurussia and Siberia/Angara), because a compara-
cially in otherwise fossil-poor wet and dry red tively large amount of material was collected due to
beds. However, the taxonomy of conchostracans coal mining during the nineteenth century. They thus
has long been oversplit, and vast taxonomic revi- received much attention in the paralic biofacies – the
sions are needed to realize fully their biostratigraphic change from fully marine faunas to somewhatDownloaded from http://sp.lyellcollection.org/ by guest on March 12, 2022
S. G. Lucas et al.
obscure brackish to limnic faunal assemblages, anthracosaurs (middle Tournaisian); (3) oldest amni-
sometimes accompanied by plants – and experienced otes (early Bashkirian); and (4) oldest high-fibre
a wide range of biostratigraphic studies leading to a herbivores (Bashkirian). Carboniferous tetrapod
zonation based on their assemblages that received footprints thus provide significant insight into some
constant refinement (e.g. Paproth et al. 1983). major events of the Carboniferous evolution of tetra-
Nevertheless, non-marine bivalves have a prob- pods, but only provide very coarse resolution for bio-
lematic taxonomy due to a general lack of morpho- stratigraphic and biochronological studies.
logical characters and a very wide range of
variation. Mostly relatively small and thin shelled, Tetrapods
their shell surface is predominantly smooth or has
only weakly developed ornamentation. The range Carboniferous tetrapod body fossils have not gener-
of variation in shape of the Carboniferous non- ally been used in non-marine biostratigraphy, and
marine bivalves is comparable to that of Recent Lucas (2021b) presents a Carboniferous, tetrapod-
limnic bivalves, and this is controlled by subtle dif- based biochronology. Tetrapod fossils of Carbonif-
ferences in substrate type, water depth, habitat erous age are known almost exclusively from the
type, and wave and current intensity, but also by soft- southern part of a palaeoequatorial Euramerican
body anatomy, sexual dimorphism or duration of the province, and their stratigraphic distribution is used
breeding period. Amler and Silantiev (2021) con- to identify five land-vertebrate faunachrons: (1) Hor-
clude that after a century of diverse research on tonbluffian (Givetian–early Visean), the time
Pennsylvanian non-marine bivalves, the concepts between the first appearance datum (FAD) of tetra-
used for species and genus definition are completely pods to the beginning of the Doran; (2) Doran (late
inadequate from a biological–systematic point. Fur- Visean–early Bashkirian), the time between the
thermore, because of recurrent convergent evolution FAD of the baphetid Loxomma and the beginning
in the non-marine bivalves, taxonomic identifica- of the Nyranyan; (3) Nyranyan (late Bashkirian–
tions depend on the stratigraphic framework pro- Moscovian), the time between the FAD of the eurep-
vided by marine horizons, particularly in the tile Hylonomus and the beginning of the Cobrean; (4)
Pennsylvanian in northwestern Europe. Cobrean (Kasimovian–late Gzhelian), the time
between the FAD of the eupelycosaur Ianthasaurus
Tetrapod footprints and the beginning of the Coyotean; and (5) Coyotean
(late Gzhelian–early Permian), the time between the
Fossil footprints of Carboniferous tetrapods, studied FAD of the eupelycosaur Sphenacodon and the
since the early 1800s, are common in some Carbon- beginning of the Seymouran.
iferous non-marine strata and had very broad palae- This biochronology provides insight into some
ogeographical distributions. Furthermore, some important evolutionary events in Carboniferous
Carboniferous non-marine strata that lack or nearly tetrapod evolution: (1) no data support tetrapod
lack a tetrapod bone record have an extensive foot- mass extinctions across the Devonian–Carbonifer-
print record. Therefore, various workers have used ous boundary; (2) Romer’s gap was mostly an arte-
Carboniferous tetrapod footprints in biostratigraphy. fact of the available fossil sample that is being
Lucas et al. (2021b) review the Carboniferous filled by discovery and by description of already
record of tetrapod footprints, which is mostly of known fossils; (3) the almost total restriction of Car-
Euramerican origin. Particularly significant is the boniferous tetrapod fossils to southern Euramerica is
Carboniferous tetrapod footprint record of the Mari- certainly in part due to a lack of sampling outside of
times Basin of eastern Canada (New Brunswick, that region but in part reflects the taphonomic mega-
Nova Scotia and Prince Edward Island), which bias introduced by coal mining in what was equato-
encompasses well-dated and stratigraphically super- rial Euramerica and the late Paleozoic ice ages,
posed footprint assemblages of Early Mississippian– which are likely to have made the poleward regions
early Permian age. A global footprint biostratigraphy (especially much of Gondwana) uninhabitable for
and biochronology of Carboniferous time identifies tetrapods during substantial intervals of Carbonifer-
four tetrapod-footprint biochrons: (1) stem-tetrapod ous time; (4) the oldest definite reptile body fossils
biochron of Middle Devonian–early Tournaisian (and footprints) are of Nyranyan age, and this has
age; (2) Hylopus biochron of middle Tournaisian– remained a remarkably stable biostratigraphic
early Bashkirian age; (3) Notalacerta–Dromopus datum for more than a century; (5) an important
interval biochron of early Bashkirian–Kasimovian increase in diversity of tetrapods is the Nyranyan
age; and (4) Dromopus biochron of Kasimovian– diversification event, a turning point in Carbonifer-
early Permian age. The Carboniferous tetrapod foot- ous tetrapod evolution; (6) there were important
print record provides these important biostrati- changes in the tetrapod biota during the Middle–
graphic datums: (1) oldest temnospondyls (middle Late Pennsylvanian transition, part of the ‘Kasimo-
Tournaisian); (2) oldest reptiliomorphs, likely to be vian revolution’; and (7) the Coyotean chronofaunaYou can also read

















































