SUBSTRATE AFFINITY AND SPECIFICITY OF THE SCSTH1P BROMODOMAIN ARE FINE-TUNED FOR VERSATILE HISTONE RECOGNITION - BIORXIV
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Substrate Affinity and Specificity of the ScSth1p Bromodomain
Are Fine-Tuned for Versatile Histone Recognition
Bartlomiej J. Blus,1,2,* Hideharu Hashimoto,1,3 Hyuk-Soo Seo,4 Aleksandra Krolak,2
and Erik W. Debler3,*
1
These authors contributed equally.
2
Laboratory of Cell Biology, Howard Hughes Medical Institute, The Rockefeller
University, New York, NY 10065, USA.
3
Department of Biochemistry & Molecular Biology, Thomas Jefferson University,
Philadelphia, PA 19107, USA.
4
Department of Cancer Biology, Dana-Farber Cancer Institute, Boston, MA 02215, USA.
*
Correspondence: bblus@rockefeller.edu (B.J.B.) and Erik.Debler@jefferson.edu
(E.W.D)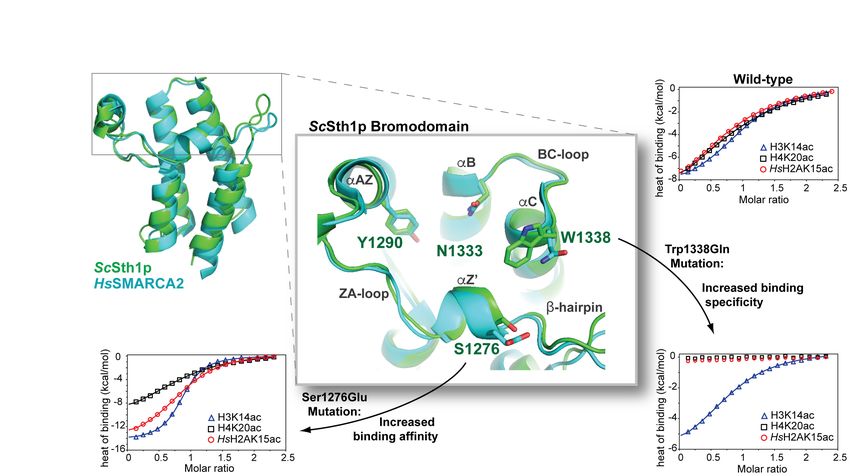
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Graphical Abstract
2
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Highlights
• The ScSth1p bromodomain preferentially recognizes H3K14ac and H4K20ac
peptides
• Ser1276 and Trp1338 are key determinants of substrate affinity and specificity
• Mutations of these residues drastically increase substrate affinity and specificity
• The ScSth1p bromodomain is fine-tuned for promiscuous histone tail recognition
3
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Summary
Bromodomains recognize a wide range of acetylated lysine residues in histones and other
nuclear proteins. Substrate specificity is critical for their biological function and arises
from unique acetyl-lysine binding sites formed by variable loop regions. Here, we
analyzed substrate affinity and specificity of the yeast ScSth1p bromodomain, an
essential component of the “Remodels the Structure of Chromatin” complex, and found
that the wild-type bromodomain preferentially recognizes H3K14ac and H4K20ac
peptides. Mutagenesis studies—guided by our crystal structure determined at 2.7 Å
resolution—revealed loop residues Ser1276 and Trp1338 as key determinants for such
interactions. Strikingly, point mutations of each of these residues substantially increased
peptide binding affinity and selectivity, respectively. Our data demonstrate that the
ScSth1p bromodomain is not optimized for binding to an individual acetylation mark, but
fine-tuned for interactions with several such modifications, consistent with the versatile
and multivalent nature of histone recognition by reader modules such as bromodomains.
Keywords: Chromatin, epigenetics, bromodomain, histone tails, lysine acetylation,
chromatin remodeling, substrate specificity, binding promiscuity, x-ray crystallography,
isothermal titration calorimetry
4
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Introduction
Protein acetylation is a ubiquitous post-translational modification primarily found in
eukaryotes. Site-specific acetylation of lysine residues in histone tails plays a major role
in regulating chromatin structure and gene expression (Garcia et al., 2007; Huang et al.,
2015; Latham and Dent, 2007). Acetylated histone tails are a hallmark of active genes
and multiple acetyl-lysine marks are often enriched at active promoter transcription start
sites (TSSs) (Garcia et al., 2007). Histone acetylation is a dynamic and reversible process
mediated by histone acetyltransferases and deacetylases (Weiner et al., 2015). The acetyl-
lysine is recognized primarily by bromodomains, but also by the YEATS and Tudor
domains (Filippakopoulos et al., 2012; Jurkowska et al., 2017; Zhao et al., 2017). Besides
acetylation, bromodomains have been shown to recognize propionylated and butyrylated
lysines (Vollmuth and Geyer, 2010). Consistent with their central role in chromatin
biology and gene transcription, many bromodomains become dysregulated in numerous
diseases including cancer and inflammatory disorders, and thus have emerged as
prominent epigenetic drug targets (Filippakopoulos and Knapp, 2014; Fujisawa and
Filippakopoulos, 2017).
Bromodomains consist of a left-handed four-helical bundle (αZ, αA, αB, αC) and
variable ZA and BC loops involved in substrate recognition (Dhalluin et al., 1999;
Filippakopoulos et al., 2012). They have conserved motifs in their helices and loops and
can be divided into eight families using a structure-based alignment (Filippakopoulos and
Knapp, 2012; Filippakopoulos et al., 2012). Most bromodomains contain PDY (Pro-Asp-
Tyr) and PϕD (ϕ denotes a hydrophobic residue) motifs in the ZA-loop. Other
5
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
bromodomain features are conserved to a lesser extent and are family-specific. For
instance, families I and II contain the WPF (Trp-Pro-Phe) “shelf” motif within the ZA
loop, which is equivalent to the E/DϕF motif in family VIII. The acetyl-lysine residue is
recognized by a conserved hydrophobic pocket located at the end of the four-helix bundle
in each bromodomain (Dhalluin et al., 1999). In particular, two highly conserved
residues—an Asn at the C-terminus of the αB helix and a Tyr in the ZA loop—form
canonical hydrogen bonds with the acetyl moiety in the target protein or peptide and the
conserved network of water molecules in the acetyl-lysine binding pocket, respectively.
In stark contrast to this conserved pocket, the surrounding surface area is formed by
highly variable regions which confer sequence-specific recognition of the residues
adjacent to the acetylated lysine (Vidler et al., 2012). Biochemical profiling of histone
peptide interactions has revealed that most bromodomains recognize several different
acetyl-lysine marks with micromolar to millimolar affinities (Filippakopoulos and Knapp,
2012). In addition to this binding promiscuity, multivalency of histone recognition by
more than one “reader” module is well established, generating avidity and rationalizing
why high affinity of recognition motifs to individual marks is often not observed
(Ruthenburg et al., 2007).
In order to examine the interplay between substrate binding affinity and specificity, we
set out to characterize these properties in the ScSth1p bromodomain from Saccharomyces
cerevisiae. This bromodomain is among the few yeast bromodomains that bind distinct
acetylated histone peptides with apparent high affinity (Zhang et al., 2010). The ScSth1p
protein is an ATP-dependent helicase associated with the “Remodels the Structure of
6
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Chromatin” (RSC) complex involved in transcription regulation and nucleosome
positioning (Cairns et al., 1996; Lorch et al., 2018; Rawal et al., 2018). The C-terminal
bromodomain in ScSth1p is essential and recruits the RSC complex to active TSSs (Du et
al., 1998). In this study, we determined the affinity of the ScSth1p bromodomain towards
several acetylated histone peptides by isothermal titration calorimetry (ITC) and report its
crystal structure at 2.7 Å resolution. Comparison with other bromodomains allowed us to
identify loop residues that are unique to ScSth1p. Strikingly, point mutations of these
residues substantially increase substrate specificity and binding affinity of the ScSth1p
bromodomain, corroborating the notion that bromodomains are fine-tuned for versatile
histone recognition.
Results
The ScSth1p Bromodomain Preferentially Recognizes H3K14ac and H4K20ac
Peptides
Using the SMART database (Letunic et al., 2015), we designed a ScSth1p bromodomain
construct encoding residues 1250-1359, which was expressed as a stable protein for
biophysical and structural characterization (Figures 1A and 1B). The ScSth1p
bromodomain was previously shown to preferentially bind to three acetylated peptides
derived from human histones: H3K14ac, H4K20ac, and HsH2AK15ac (Zhang et al.,
2010). Using isothermal titration calorimetry (ITC), we found that the ScSth1p
bromodomain binds these peptides with dissociation constants (Kds) of ~18 µM, 48 µM,
and 44 µM, respectively (Figure 2B). These interactions are acetyl-lysine specific, as the
unmodified histone peptides did not bind (Figures S1A and S1B). Likewise, alanine
7
bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
mutations of the two highly conserved acetyl-lysine recognizing residues, Tyr1290Ala
and Asn1333Ala, completely abolished the ScSth1p bromodomain interactions with these
acetylated peptides (Figures S1C and S1D). While the human and yeast H3 and H4
histone tails are identical (which is why we omit their species designations in this
manuscript), the histone H2A sequences vary significantly between the two species. Due
to these differences, we also tested two yeast peptides homologous to HsH2AK15ac:
ScH2AK13ac and ScHtz1K14ac (Figure 2A). Interestingly, these two peptides do not
interact with the ScSth1p bromodomain (Figure 2C), suggesting that histone residues
adjacent to the acetyl-lysine are critical for binding.
Inspection of the peptide sequences used in our binding assay reveals that the interacting
acetylated histones are enriched in positively charged residues compared to the non-
interacting peptides (Figure 2A and Table S1). The available structures of bromodomain
complexes with histone peptides indeed indicate that lysine and arginine residues
flanking the acetylated lysine participate in electrostatic interactions with the
bromodomain (Plotnikov et al., 2014; Zeng et al., 2008). However, when we varied the
ionic strength and pH of the buffer, the Kd values measured by ITC were not significantly
altered (up to fourfold) (Figure S2). These results imply that electrostatic forces do not
dominate the ScSth1p bromodomain interactions with the tested acetylated histone
peptides.
The Crystal Structure of the ScSth1p Bromodomain Reveals a Hydrophobic Acetyl-
Lysine Peptide Binding Site
8bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
In order to obtain structural insights into histone recognition by the ScSth1p
bromodomain, we determined its crystal structure at 2.7 Å resolution. Crystallographic
phases were determined by molecular replacement. The asymmetric unit contains six
bromodomains that are highly similar to each other with a root mean square deviation of
0.5 Å. The structure was refined to Rwork/Rfree of 23.8%/27.0% and features excellent
geometry as evaluated by MolProbity (Table 1) (Chen et al., 2010).
The ScSth1p bromodomain folds into a canonical left-handed four-helix bundle (αZ, αA,
αB, αC). In the variable ZA loop, the PDY motif contributes to the αAZ helix, while the
PMA motif, which deviates from the consensus PϕD sequence, marks the end of this loop
(Figures 1C and 3A) (Dhalluin et al., 1999; Filippakopoulos et al., 2012). Notably, the
surface surrounding the acetyl-lysine binding pocket of the bromodomain is relatively
hydrophobic, consistent with the lack of large effects on peptide binding affinity at
varying salt concentrations and buffer pH (Table S1 and Figure S2). This hydrophobic
area distinguishes the ScSth1p bromodomain from the polar site of its human homolog
HsSMARCA2 (Figure S4).
The ScSth1p Bromodomain Contains a β-Hairpin Characteristic of Family VIII
Members
The β-hairpin located between the αZ and αZ’ helices classifies the ScSth1p
bromodomain as a family VIII member (Figures 1C and 3B). A distance matrix
alignment (DALI) search (Holm and Rosenstrom, 2010) against known structures in the
Protein Data Bank (PDB) indeed confirms that the ScSth1p bromodomain is structurally
9bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
most similar to other family VIII members, including bromodomains of HsSMARCA2
and HsPB1 (Figures 1C and 3B) (Filippakopoulos et al., 2012; Singh et al., 2007)). The
β-hairpin in the ScSth1p bromodomain is stabilized by a salt bridge between His1272 and
Asp1342 in the αC helix (Figure 3C). Disruption of this salt bridge by the His1272Ala
mutation did not affect peptide specificity or binding affinity, suggesting that it does not
significantly contribute to recognizing acetylated histones (Figures 3D and S3A).
Two Single Point Mutations in the ScSth1p Bromodomain Dramatically Increase
Substrate Affinity and Specificity
Another common feature of the family VIII bromodomains is the E/DϕF sequence, which
substitutes for the “WPF shelf motif” found in many bromodomains (Figures 1C and 3A).
However, the corresponding “SIF” motif in ScSth1p deviates from this consensus
sequence. Like the WPF motif, the SIF sequence folds into a short 310-helix termed αZ’.
To examine the contribution of this non-conserved serine to acetylated histone peptide
binding (Figure 3E), we mutated it to the more common glutamate found in several
family VIII bromodomains (Figure 1C). Strikingly, the Ser1276Glu mutation increases
peptide binding affinity by up to sixfold with respect to the wild-type bromodomain,
resulting in a tenfold tighter binding to H3K14ac than to H4K20ac (Kd values of 3 and 31
µM, respectively; Figures 3F and S3B).
In addition to classifying bromodomains based on their entire structures (Filippakopoulos
et al., 2012), they can also be grouped according to the conservation of signature residues
in their variable acetyl-lysine binding sites (Vidler et al., 2012). According to the latter
10bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
classification, the ScSth1p bromodomain shares conserved elements with classes 5 and 6.
These include residues Ser1337, Val1339, Asp1342, and Tyr1287 that line the acetyl-
lysine binding pocket (Figures 3G and 1C). Notably, Trp1338 at the end of the BC loop is
not present in any other bromodomain and the equivalent position is most often occupied
by Ala, Glu, or Gln (Filippakopoulos et al., 2012; Zhang et al., 2010). When we mutated
Trp1338 to the more common glutamine found in family VIII members (Figure 1C), we
observed a dramatic effect on substrate specificity. While the Trp1338Glu mutant
bromodomain binding to H3K14ac was comparable to the wild-type protein, its
interactions with the H4K20ac and HsH2AK15ac peptides were completely abolished
(Figures 3H and S3C). Hence, our results show that two single point mutations of the two
non-conserved residues, Ser1272 and Trp1338, significantly increase ScSth1p
bromodomain binding affinity and specificity towards acetylated histone peptides.
Discussion
One of the major challenges in chromatin biology is to understand how distinct patterns
of post-translational modifications are recognized and “interpreted” or “read” by
chromatin interacting proteins and complexes. Bromodomains are ubiquitous protein
modules that bind site-specific acetylated lysines in histones, a key modification
implicated in transcriptional activity. Most bromodomains recognize multiple acetylated
substrates with a wide range of selectivity and modest binding affinity (Filippakopoulos
and Knapp, 2012). While it has been accepted that such binding behavior can be
understood in light of cooperative engagement with several histone modifications
(Ruthenburg et al., 2007), the question arises whether there are intrinsic, evolutionary
11bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
limitations to the bromodomain fold that preclude it from recognizing substrates with
high, i.e. nanomolar affinities. To address this issue, we focused on the ScSth1p
bromodomain, which binds several acetylated histone peptides with apparent high
affinity in a dot-blot assay (Zhang et al., 2010). We discovered that it was surprisingly
straightforward to substantially increase both substrate affinity and specificity by
generating two point mutations in the bromodomain, changing only one residue at a time.
The first of these two key residues is Ser1276 of the SIF motif within the variable ZA
loop, which deviates from the acidic residues in the consensus E/DϕF sequence of family
VIII bromodomains and from the tryptophan in the WPF motif present in several other
bromodomain families (Figure 1C) (Filippakopoulos et al., 2012). For example, Ser1276
corresponds to Glu206 in the second bromodomain of the human polybromodomain
protein PB1, the only family VIII member whose structure has been determined in
complex with a histone peptide to date (Charlop-Powers et al., 2010). In this NMR
structure, Glu206 plays a key role in recognizing the Kac +1 residue alanine (Ala15) of
the H3K14ac peptide, discriminating histone peptides with a sequence variation at this
position (Charlop-Powers et al., 2010). Consistent with this observation, the Ser1276Glu
mutation largely increased selectivity for H3K14ac over H4K20ac or HsH2AK15ac
peptides. We envision that the longer side chain of Glu1276 in the mutant bromodomain
is able to accommodate the Ala side chain at the +1 position in H3K14ac, but the bulkier
side chains of Ile and Thr in the H4K20ac or HsH2AK15ac peptides fit to a lesser degree.
We conclude that the exposed ZA loop residue at the position equivalent to Ser1276 is a
major determinant of substrate affinity in the family VIII bromodomains.
12bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
The second residue that we identified as a key determinant for the ScSth1p
bromodomain-peptide interactions corresponds to Trp1338 at the end of the BC loop,
proximal to Ser1276 (Figure 3E). Alignment of the 61 human and 14 yeast bromodomain
sequences illustrates that a tryptophan residue is unique at this position, which is
occupied by a glutamine in the majority of family VIII members (Filippakopoulos et al.,
2012). Strikingly, our Trp1338Gln mutation had an even larger impact on peptide
recognition than the Ser1276Glu mutation; it completely abolished the ScSth1p
bromodomain binding to the H4K20ac and HsH2AK15ac peptides, while the affinity for
H3K14ac essentially remained unchanged. These results suggest that the large aromatic
surface area of the exposed Trp1338 may contribute extensive contacts with the
interacting peptides.
Our proposed role for Trp1338 is corroborated by its prominent location at the
intersection of two roughly perpendicular grooves that are poised to accommodate bound
peptides. The first groove is hydrophobic and extends above the canonical N-acetyl-
lysine binding pocket between the ZA and BC loops, which corresponds to the preferred
peptide binding groove in experimentally determined bromodomain-peptide complexes
(Figure 3E) (Fujisawa and Filippakopoulos, 2017; Marchand and Caflisch, 2015; Zeng et
al., 2008). The second groove is slightly negatively charged and is formed between the β-
hairpin and a surface patch formed by Trp1338 and Asp1267. A binding mode of
peptides wrapping around Trp1339 would be consistent with the large effect that the
Trp1339Gln mutation has on peptide recognition. Precedence for such a binding mode is
found in the structure of the HsBrd4 bromodomain complex with a diacetylated histone
13bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
peptide (PDB code 3UVW), in which the peptide bends around Asp145 at the Trp1339-
equivalent position at a 90° angle (Filippakopoulos et al., 2012). In contrast to Trp1338,
the opposite edge of the second groove appears to be less important for peptide
recognition, as the His1272Ala mutation, which presumably destabilizes the β-hairpin,
has a negligible effect on peptide binding. Overall, the predominantly apolar character of
the two binding grooves agrees well with our finding that the ScSth1p bromodomain-
peptide interactions are dominated by hydrophobic contacts.
Our results on substrate specificity of the ScSth1p bromodomain have important
implications for the recruitment of its resident RSC chromatin remodeling complex. In S.
cerevisiae, the H3K14ac and Htz1K14ac marks are enriched at TSSs, but it is unknown
whether both are required for chromatin remodeling initiation (Pokholok et al., 2005;
Ramachandran et al., 2015). As the ScSth1p bromodomain recognizes H3K14ac, but not
the ScHtz1K14ac mark (Figure 2C), our data suggest that ScSth1p recruits the RSC
complex to TSSs via the interaction with the H3K14ac mark. Indeed, Htz1-depleted cells
do not alter nucleosome positioning at TSSs in yeast (Hartley and Madhani, 2009),
further suggesting that the Htz1 histone H2A variant may not be required for RSC
recruitment to chromatin. Given that ScRsc4p represents another H3K14ac reader module
in the RSC complex (Kasten et al., 2004; VanDemark et al., 2007), these two essential
RSC components may cooperatively bind to two histone tails, thereby increasing the
binding affinity and specificity of the RSC complex at active TSSs (Figure 4). In addition
to the H3K14ac binding, the ScSth1p bromodomain recognizes the H4K20ac
modification, yet its biological function in yeast remains elusive.
14bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
In conclusion, our structure-based mutational analysis shows that substrate binding
affinity and specificity are finely balanced by individual residues in the ScSth1p
bromodomain. Instead of recognizing a single chromatin mark with high affinity, our
results are consistent with the notion that multiple histone modifications—each
interacting with intermediate affinity—are cooperatively “read” with high affinity and
specificity. In this scenario, the bromodomains of the RSC components ScSth1p and
ScRsc4p may cooperatively facilitate recruitment of the RSC complex towards a subset
of TSSs marked by the H3K14 acetylation. Our ScSth1p bromodomain point mutants
may prove as valuable tools to probe the mechanism of ScSth1p-dependent recruitment of
the RSC complex to chromatin in vivo.
15bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Materials and Methods
Protein Expression and Purification
The DNA fragment corresponding to the S. cerevisiae Sth1p bromodomain (residues
1250-1359) was generated by PCR from genomic DNA and cloned into a modified
pGEX-4T1 vector (Addgene) using Ligation Independent Cloning (LIC), as previously
described (Seo et al., 2013). The resulting fusion protein contained an N-terminal GST
tag followed by a cleavage site for the tobacco etch virus (TEV) protease. The ScSth1p
bromodomain was expressed in E. coli BL21-CodonPlus(DE3)-RIL cells (Stratagene)
following overnight induction at an OD600nm of 0.8 with 0.5 mM isopropyl-β-D-thio-
galactoside (IPTG) at 18°C in LB media containing chloramphenicol (34 mg/L) and
ampicillin (100 mg/L). Cells were lysed using a cell disruptor (Avestin) in a lysis buffer
containing 20 mM HEPES (pH 7.5), 300 mM NaCl, 0.5 mM EDTA, 5% (v/v) glycerol, 3
mM DTT, 1 mM benzamidine, supplemented with phenylmethanesulfonyl fluoride
(PMSF), 1 µg/mL bovine lung aprotinin and leupeptin (Sigma). After centrifugation at
30,000 × g, the supernatant was loaded onto a GSTrap column (GE Healthcare) and
washed extensively with lysis buffer. The ScSth1p bromodomain was eluted with 20 mM
HEPES pH 7.5, 200 mM NaCl, 5% (v/v) glycerol, and 3 mM DTT supplemented with 15
mM reduced glutathione (Sigma). Following an on-column GST-tag cleavage by TEV
protease at 4°C overnight, the bromodomain was further purified over a HiLoad 16/60
Superdex 75 gel filtration column (GE Healthcare) equilibrated with 20 mM HEPES pH
7.5, 200 mM NaCl, and 3 mM DTT. Peak elution fractions were concentrated to 5-10
mg/mL, flash-frozen in liquid nitrogen, and stored at -80°C.
16bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Peptide Synthesis
Synthetic histone peptides for ITC binding experiments were synthesized and HPLC-
purified to 95-98% purity at the Proteomics Resource Center (The Rockefeller
University). The correct sequence for each peptide was confirmed by mass spectrometry.
All peptides are 15 amino acids long and contain a C-terminal non-histone tyrosine for
concentration measurements at OD280nm.
ScSth1p Bromodomain Mutagenesis
Constructs for the ScSth1p bromodomain mutants (Ser1276Glu, His1272Ala,
Trp1338Gln, or Tyr1290Ala/Asn1333Ala) were generated using QuikChange site-
directed mutagenesis kit (Stratagene). Mutant bromodomain expression and purification
were carried out as for the wild-type protein. Gel filtration-purified protein was
concentrated to 5-10 mg/mL, flash-frozen in liquid nitrogen and stored at -80°C.
ITC Binding Assays
Samples for the ITC experiments were dialyzed extensively against 20 mM HEPES pH
7.5, 150 mM NaCl, 0.5 mM TCEP using Float-A-Lyzer dialysis devices with a molecular
weight cutoff of 0.5-1 kDa (Spectrum Laboratories). For a subset of ITC experiments,
composition of the binding buffer (pH 7.5) was adjusted to 75 or 300 mM NaCl, or the
buffer pH was set to 6.0 or 8.5 at 150 mM NaCl (see Table S1 for details). After dialysis,
purified bromodomain and histone peptides were filtered (0.22 µm) and centrifuged,
followed by determining their concentration by UV absorbance at 280 nm. All titration
experiments were performed using an automated MicroCal Auto-iTC200 instrument (GE
17bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Healthcare) at 15°C. For each titration, a concentrated solution of the peptide (at 1 mM)
was titrated into the ITC reaction cell containing the wild-type or mutant ScSth1p
bromodomain (at 90 µM). The heat of sample dilution was separately measured for each
experiment or determined from the final plateau of each titration, and subtracted from the
binding data. The corrected ITC curves were analyzed using a nonlinear least-squares
minimization method in ORIGIN 7.0 to determine the binding stoichiometry (n),
dissociation constant (Kd) and enthalpy (ΔH). These parameters were subsequently used
to determine free Gibbs energy (ΔG) and the entropic component (TΔS) using ∆G = -RT
ln(1/Kd) and TΔS = ΔH – ΔG equations, where R and T are gas constant (1.99
cal/(mol*K)) and absolute temperature (288 K), respectively. The fitting parameters were
allowed to float freely in the fitting procedure. All titrations were repeated at least twice.
Thermodynamic parameters for representative ITC titrations are provided in Table S1.
Crystallization and Structure Determination of the ScSth1p Bromodomain
Crystals of the ScSth1p bromodomain were grown at 20°C in sitting drops containing 1
µL of the protein at 5 mg mL-1 and 1 µL of a reservoir solution consisting of 0.2 M
magnesium chloride hexahydrate, 0.1 M BIS-TRIS pH 6.5, 25% (w/v) polyethylene
glycol 3,350, and 3% (w/v) trimethylamine N-oxide dihydrate. For cryoprotection,
crystals were stabilized in the mother liquor supplemented with 20% (v/v) glycerol and
flash cooled in liquid nitrogen. X-ray diffraction data were collected at 100 K at the NE-
CAT beamline 24ID-E at the Advanced Photon Source, Argonne National Laboratory,
and processed using XDS (Kabsch, 2010). The crystals belong to the space group C121,
with unit cell dimensions of a=141.8 Å, b=74.6 Å, c=72.5 Å, α=γ=90°, and β=92.9° and
18bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
diffracted to 2.7 Å resolution (Table 1). Crystallographic phases were obtained by
molecular replacement using the fourth bromodomain of human poly-bromodomain
containing protein 1 (PB1; pdb: 3TLP) as a search model using PHASER (McCoy et al.,
2007) from the CCP4 program suite. After molecular replacement, iterative cycles of
manual rebuilding and restrained refinement were carried out in COOT (Emsley and
Cowtan, 2004) and PHENIX (Adams et al., 2010), respectively. The stereochemical
quality of the structural model was assessed with MolProbity (Chen et al., 2010). There
were no residues in the disallowed region of the Ramachandran plot (Table 1). Figures
were generated with PyMOL (DeLano Scientific) (Schrodinger, 2015).
Acknowledgements
First and foremost, we would like to acknowledge our mentor G. Blobel for supporting
our research. We would like to thank C. Adura for helping us with the Auto-iTC200
instrument (High-Throughput and Spectroscopy Resource Center, Rockefeller
University), H. Zebroski III for peptide synthesis (Proteomics Resource Center,
Rockefeller University), and R. Rajashankar (NE-CAT) for assisting with x-ray data
collection. We would like to thank E. Coutavas for critical reading of the manuscript.
This work was funded by the Rockefeller University and the Howard Hughes Medical
Institute.
19bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Author Contributions
B.J.B., H.H, H-S.S, and E.W.D designed the experiments; B.J.B., H.S-S., and A.K
performed the experiments; B.J.B., H.H., H-S.S, and E.W.D analyzed the data, and
B.J.B., H.H., A.K., and E.W.D. wrote the manuscript.
Accession Code
The ScSth1p bromodomain coordinates and structure factors have been deposited at the
Protein Data Bank (PDB) with the accession number 6MR4.
20bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure 1. Purification and Sequence Alignment of the ScSth1p bromodomain.
(A) Domain architecture of the ScSth1p protein. The N-terminal helicase/SANT-
associated (HSA) domain, the catalytic ATPase module consisting of the recA domains,
Snf2 ATP coupling (SnAc) domain, and the C-terminal bromodomain are shown.
(B) A size-exclusion chromatography (SEC) elution profile for the ScSth1p
bromodomain (residues 1250-1359) is shown together with peak elution fractions
analyzed by SDS-PAGE. The bromodomain elutes as a single monomeric peak.
(C) A structure-guided multiple sequence alignment of the ScSth1p bromodomain with
similar yeast and human bromodomains. The sequences of the six bromodomains in the
human PB1 protein are displayed. Secondary structure elements and the loop regions are
shown above the alignment. Similar and identical residues within the conserved
bromodomain sequences are colored grey and yellow, respectively, and the residues
mutated in this study are highlighted in red. Highly-conserved sequence motifs across all
bromodomain families are shown below the alignment. Φ and x denote hydrophobic and
any residues, respectively.
Figure 2. The ScSth1p Bromodomain Specifically Interacts with Histone Marks
Associated with Active Gene Transcription
(A) Primary sequences of the acetylated 15-mer histone peptides used in the ITC binding
assay. Acetyl-lysine is highlighted in the center for each peptide. Positively and
negatively charged residues are colored in blue and red, respectively. The +1 position in
each sequence marks a residue critical for bromodomain specificity. All peptides contain
a C-terminal non-histone tyrosine (y) for spectrophotometric concentration determination
21bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
at 280 nm.
(B-C) ITC binding profiles for the ScSth1p bromodomain interactions with acetylated
histone peptides: H3K14ac, H4K20ac, HsH2AK15ac (panel B) and ScHtz1K14ac,
ScH2AK13ac (panel C). Raw heat signals generated during each titration are displayed in
the top panel as time traces. Below, integrated titration data are displayed and were
analyzed by the one-set of sites model (continuous lines). The determined dissociation
constants (Kds) are displayed for the interacting peptides in (B), whereas no binding
(N.B.) was detected in (C).
Figure 3. Structure and Mutational Analysis of the ScSth1p Bromodomain
(A) Overall structure of the ScSth1p bromodomain shown with labeled secondary
structure elements: αZ (blue), ZA-loop (magenta), αB (green), αC (orange) and αA
(pink). The ZA-loop contains the SIF motif as part of the αZ’ helix and the β-hairpin.
(B) Structural alignment of the ScSth1p and HsSMARCA2 (pdb: 4QY4) bromodomains.
(C) A detailed view of the interactions between the β-hairpin and αC helix. His1272 in
the β-hairpin forms two hydrogen bonds: one with Asp1270 in the β-hairpin, the other
with Asp1342 in the αC helix.
(D) ITC binding profiles for interactions between the ScSth1p His1272Ala mutant
bromodomain and H3K14ac, H4K20ac or HsH2AK15ac peptides. Binding data were
analyzed by the one-set of sites model (continuous lines) and the obtained Kd values are
displayed.
(E) Electrostatic surface potential of the ScSth1p bromodomain (left panel). Positive and
negative surface potential is colored in blue and red, respectively. The electrostatic
22bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
surface potential scale is shown, where kB is the Boltzmann constant and T is the absolute
temperature. A cartoon representation of the ScSth1p bromodomain structure is displayed
in the same orientation with the mutated residues highlighted (right panel). The
hydrophobic acetyl-lysine binding pocket and the slightly negatively charged cleft
between the β-hairpin and αC helix are indicated.
(F) ITC binding profiles for the interactions between the ScSth1p Ser1276Glu mutant
bromodomain and H3K14ac, H4K20ac, and HsH2AK15ac peptides. Binding data were
analyzed by the one-set of sites model (continuous lines) and the obtained Kd values are
displayed.
(G) A close-up view of the acetyl-lysine binding site in the ScSth1p bromodomain.
(H) ITC binding profiles for the interactions between the ScSth1p Trp1338Gln mutant
bromodomain and H3K14ac, H4K20ac, and HsH2AK15ac peptides. The H3K14ac
peptide binding data were analyzed by the one-set of sites model (continuous lines),
whereas the remaining peptides do not bind (N.B.).
Figure 4. The ScSth1p Bromodomain Contributes to the RSC Complex Association
with H3K14ac-Modified Chromatin.
A proposed schematic representation of the RSC complex binding to transcriptionally
active chromatin. The bromodomains in ScSth1p and ScRsc4p—key members of the RSC
complex—are thought to interact with acetylated histones to direct RSC to promoter
transcription start sites (TSSs) marked with H3K14 acetylation. The remaining
bromodomain-containing components of the RSC complex, ScRsc1 and ScRsc2, are
23bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
omitted for clarity. Once attached to promoters, the RSC complex creates a nucleosome-
free region flanked by strongly positioned nucleosomes.
24bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure 1.
25bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure 2.
26bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure 3.
27bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure 4.
28bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Table 1.
Table 1. Data Processing and Refinement Statistics
PDB ID 6MR4
Wavelength [Å] 0.97918
Space group C121
Unit cell (a, b, c) [Å] 141.8, 74.6, 72.5
Unit cell (α, β, γ ) [°] 90.0, 92.9, 90.0
Resolution [Å] 72.45-2.71 (2.84-2.71)a
Rmerge 0.083 (0.581)
10.8 (1.7)
Completeness [%] 98.6 (99.5)
Redundancy 3.1 (3.2)
CC ½ 0.99 (0.60)
Obs. Reflections 64,102 (8,612)
Unique Reflections 20,385 (2,722)
Refinement
Resolution [Å] 2.71
No. Reflections 20,371
Rwork/ Rfree [%] 23.8/27.0
No. Atoms
Protein 5476
Solvent 27
B-factors [Å2]
Protein 41.6
Solvent 58.9
R.m.s. deviations
Bond lengths [Å] 0.006
Bond angles [°] 0.6
All atom clash score 7.7
Ramachandranb [%]
Favored 98.74
Allowed 1.26
Rotamer outliers [%] 0
Cβ deviation 0
Data were collected on a single crystal.
a
Values in parentheses are for highest-resolution shell.
b
As determined by MolProbity.
29bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Supplemental Information
Substrate Affinity and Specificity of the ScSth1p Bromodomain
Are Fine-tuned for Versatile Histone Recognition
Bartlomiej J. Blus, Hideharu Hashimoto, Hyuk-Soo Seo, Aleksandra Krolak, and Erik W.
Debler
30bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure S1.
Figure S1. Related to Figure 2. The ScSth1p Bromodomain Contains a Canonical
Binding Pocket for Acetyl-Lysine Recognition.
(A-B) ITC binding profiles for the ScSth1p bromodomain interactions with unmodified
(non-acetylated) (A) H3K14 and (B) H4K20 peptides are shown in the top panel as time
traces. No appreciable heat of binding (N.B.) was observed for either peptide. (C) ITC
binding profiles for the ScSth1p Tyr1290Ala/Asn1333Ala mutant bromodomain
interactions with H3K14ac, H4K20ac, and HsH2AK15ac peptides. No appreciable heat
of binding (N.B.) was observed. (D) A size-exclusion chromatography (SEC) elution
profile for the ScSth1p bromodomain mutant bromodomain (Tyr1290Ala/Asn1333Ala).
Below, peak elution fractions were resolved by SDS-PAGE. The mutant ScSth1p
bromodomain elutes as a single monomeric peak.
31bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure S2.
Figure S2. Related to Figure 2. Effect of Buffer Ionic Strength and pH on ScSth1p
Bromodomain Interactions with Acetylated Histone Peptides.
(A-B) ITC binding profiles for the ScSth1p bromodomain interactions with H3K14ac and
H4K20ac peptides at various binding conditions. ITC titrations were performed in 20
mM HEPES pH 7.5, 0.5 mM TCEP supplemented with 75 mM or 300 mM NaCl (panel
A) or in 20 mM HEPES pH 6.0 or 8.0, 0.5 mM TCEP, and 150 mM NaCl (panel B).
Binding data were analyzed by the one-set of sites model (continuous lines) and the
obtained Kd values are displayed.
32bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure S3.
Figure S3. Related to Figure 3. SEC Elution Profiles for the ScSth1p Mutant
Bromodomains.
(A-C) Size-exclusion chromatography (SEC) elution profiles for the ScSth1p
bromodomain mutants. (A) The ScSth1p His1272Ala mutation likely affects the stability
of the β-hairpin, resulting in a shifted and asymmetric elution profile. (B-C) Ser1276Glu
and Trp1338Gln mutant bromodomains elute like the wild-type protein. Below, peak
elution fractions were resolved by SDS-PAGE.
33bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Figure S4.
Figure S4. Related to Figure 3. Surface Charge Distribution of the ScSth1p and
HsSMARCA2 Bromodomains.
(A) Surface charge distribution of the human SMARCA2 bromodomain (left panel).
Positively charged regions of the structure are colored in blue, whereas negative and
neutral regions are labeled in red and white, respectively. The electrostatic potential scale
is shown, where kB is the Boltzmann constant and T stands for absolute temperature. The
structure is displayed as a cartoon in the same view (right panel). The negatively-charged
acetyl-lysine binding pocket and the hydrophobic cleft formed by the β-hairpin and αC
helix are highlighted. These structural features distinguish the HsSMARCA2 and
ScSth1p bromodomains (B). The highlighted Glu1407 and Gln1473 residues in
HsSMARCA2 (A) correspond Ser1276 and Trp1338 in the ScSth1p bromodomain (B),
which we mutated in this study.
34bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Table S1: Thermodynamic Parameters for the ScSth1p Bromodomain Binding to
Acetylated Histone Peptides
ScSth1p ∆H ∆S
Peptide Binding condition Kd (µM)
bromodomain (kcal·mol-1) (cal·mol-1·K-1)
WT H3K14ac 75 mM NaCl, pH 7.5 8.8 ± 0.3 -11.1 ± 0.1 -15.4
18.2 ± 0.3
WT H3K14ac 150 mM NaCl, pH 7.5 -9.5 ± 0.1 -11.4
18.8 ± 1.7*
WT H3K14ac 300 mM NaCl, pH 7.5 32.8 ± 0.8 -10.0 ± 0.1 -14.2
WT H3K14ac 150 mM NaCl, pH 6.0 16.2 ± 0.5 -9.4 ± 0.1 -10.6
WT H3K14ac 150 mM NaCl, pH 8.5 28.2 ± 0.6 -10.0 ± 0.1 -13.7
WT H3K14 150 mM NaCl, pH 7.5 N/B - -
3.4 ± 0.1
S1276E H3K14ac 150 mM NaCl, pH 7.5 -14.6 ± 0.1 -25.5
3.3 ± 0.8*
12.9 ± 0.6
H1272A H3K14ac 150 mM NaCl, pH 7.5 -18.1 ± 0.0 -40.3
12.7 ± 1.2*
Y1290A/N133A H3K14ac 150 mM NaCl, pH 7.5 N/B - -
23.8 ± 0.6
W1338Q H3K14ac 150 mM NaCl, pH 7.5 -7.2 ± 0.1 -4.5
21.8 ± 2.7*
WT H4K20ac 75 mM NaCl, pH 7.5 28.4 ± 0.4 -12.3 ± 0.1 -21.7
48.1 ± 1.2
WT H4K20ac 150 mM NaCl, pH 7.5 -12.7 ± 0.2 -24.3
46.2 ± 4.9*
WT H4K20ac 300 mM NaCl, pH 7.5 69.9 ± 2.4 -10.4 ± 0.3 -16.9
WT H4K20ac 150 mM NaCl, pH 6.0 26.8 ± 0.1 -10.9 ± 0.1 -16.9
WT H4K20ac 150 mM NaCl, pH 8.5 62.5 ± 2.0 -10.9 ± 0.2 -18.4
WT H4K20 150 mM NaCl, pH 7.5 N/B - -
30.9 ± 1.0
S1276E H4K20ac 150 mM NaCl, pH 7.5 -12.4 ± 0.2 -22.4
30.2 ± 2.0*
84.0 ± 2.8
H1272A H4K20ac 150 mM NaCl, pH 7.5 -21.3 ± 0.1 -55.2
70 ± 20*
Y1290A/N1333A H4K20ac 150 mM NaCl, pH 7.5 N/B - -
W1338Q H4K20ac 150 mM NaCl, pH 7.5 N/B - -
WT ScHtz1K14ac 150 mM NaCl, pH 7.5 N/B - -
WT ScH2AK13ac 150 mM NaCl, pH 7.5 N/B - -
44.2 ± 0.6
WT HsH2AK15ac 150 mM NaCl, pH 7.5 -12.7 ± 0.1 -24.3
40.1 ± 4.8*
13.4 ± 0.2
S1276E HsH2AK15ac 150 mM NaCl, pH 7.5 -15.6 ± 0.1 -31.9
13.9 ± 0.6*
22.2 ± 0.5
H1272A HsH2AK15ac 150 mM NaCl, pH 7.5 -19.7 ± 0.0 -47.0
22.8 ± 2.7*
Y1290A/N133A HsH2AK15ac 150 mM NaCl, pH 7.5 N/B - -
W1338Q HsH2AK15ac 150 mM NaCl, pH 7.5 N/B - -
*Indicates the mean value of Kd and sample standard deviation error obtained
from three independent measurements
N/B indicates lack of binding
35bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
References
Adams, P.D., Afonine, P.V., Bunkoczi, G., Chen, V.B., Davis, I.W., Echols, N., Headd, J.J.,
Hung, L.W., Kapral, G.J., Grosse-Kunstleve, R.W., et al. (2010). PHENIX: a
comprehensive Python-based system for macromolecular structure solution. Acta
Crystallogr D Biol Crystallogr 66, 213-221.
Cairns, B.R., Lorch, Y., Li, Y., Zhang, M., Lacomis, L., Erdjument-Bromage, H., Tempst,
P., Du, J., Laurent, B., and Kornberg, R.D. (1996). RSC, an essential, abundant
chromatin-remodeling complex. Cell 87, 1249-1260.
Charlop-Powers, Z., Zeng, L., Zhang, Q., and Zhou, M.M. (2010). Structural insights
into selective histone H3 recognition by the human Polybromo bromodomain 2. Cell
Res 20, 529-538.
Chen, V.B., Arendall, W.B., 3rd, Headd, J.J., Keedy, D.A., Immormino, R.M., Kapral, G.J.,
Murray, L.W., Richardson, J.S., and Richardson, D.C. (2010). MolProbity: all-atom
structure validation for macromolecular crystallography. Acta Crystallogr D Biol
Crystallogr 66, 12-21.
Dhalluin, C., Carlson, J.E., Zeng, L., He, C., Aggarwal, A.K., and Zhou, M.M. (1999).
Structure and ligand of a histone acetyltransferase bromodomain. Nature 399, 491-
496.
Du, J., Nasir, I., Benton, B.K., Kladde, M.P., and Laurent, B.C. (1998). Sth1p, a
Saccharomyces cerevisiae Snf2p/Swi2p homolog, is an essential ATPase in RSC and
differs from Snf/Swi in its interactions with histones and chromatin-associated
proteins. Genetics 150, 987-1005.
Emsley, P., and Cowtan, K. (2004). Coot: model-building tools for molecular
graphics. Acta Crystallographica Section D 60, 2126-2132.
Filippakopoulos, P., and Knapp, S. (2012). The bromodomain interaction module.
FEBS Lett 586, 2692-2704.
Filippakopoulos, P., and Knapp, S. (2014). Targeting bromodomains: epigenetic
readers of lysine acetylation. Nat Rev Drug Discov 13, 337-356.
Filippakopoulos, P., Picaud, S., Mangos, M., Keates, T., Lambert, J.P., Barsyte-Lovejoy,
D., Felletar, I., Volkmer, R., Muller, S., Pawson, T., et al. (2012). Histone recognition
and large-scale structural analysis of the human bromodomain family. Cell 149, 214-
231.
Fujisawa, T., and Filippakopoulos, P. (2017). Functions of bromodomain-containing
proteins and their roles in homeostasis and cancer. Nat Rev Mol Cell Biol 18, 246-
262.
Garcia, B.A., Hake, S.B., Diaz, R.L., Kauer, M., Morris, S.A., Recht, J., Shabanowitz, J.,
Mishra, N., Strahl, B.D., Allis, C.D., et al. (2007). Organismal differences in post-
translational modifications in histones H3 and H4. J Biol Chem 282, 7641-7655.
Hartley, P.D., and Madhani, H.D. (2009). Mechanisms that specify promoter
nucleosome location and identity. Cell 137, 445-458.
Holm, L., and Rosenstrom, P. (2010). Dali server: conservation mapping in 3D.
Nucleic Acids Res 38, W545-549.
Huang, H., Lin, S., Garcia, B.A., and Zhao, Y. (2015). Quantitative proteomic analysis
of histone modifications. Chem Rev 115, 2376-2418.
36bioRxiv preprint first posted online Feb. 25, 2019; doi: http://dx.doi.org/10.1101/559005. The copyright holder for this preprint
(which was not peer-reviewed) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity.
All rights reserved. No reuse allowed without permission.
Jurkowska, R.Z., Qin, S., Kungulovski, G., Tempel, W., Liu, Y., Bashtrykov, P.,
Stiefelmaier, J., Jurkowski, T.P., Kudithipudi, S., Weirich, S., et al. (2017). H3K14ac is
linked to methylation of H3K9 by the triple Tudor domain of SETDB1. Nat Commun
8, 2057.
Kabsch, W. (2010). Xds. Acta Crystallogr D Biol Crystallogr 66, 125-132.
Kasten, M., Szerlong, H., Erdjument-Bromage, H., Tempst, P., Werner, M., and Cairns,
B.R. (2004). Tandem bromodomains in the chromatin remodeler RSC recognize
acetylated histone H3 Lys14. EMBO J 23, 1348-1359.
Latham, J.A., and Dent, S.Y. (2007). Cross-regulation of histone modifications. Nat
Struct Mol Biol 14, 1017-1024.
Letunic, I., Doerks, T., and Bork, P. (2015). SMART: recent updates, new
developments and status in 2015. Nucleic Acids Res 43, D257-260.
Lorch, Y., Maier-Davis, B., and Kornberg, R.D. (2018). Histone Acetylation Inhibits
RSC and Stabilizes the +1 Nucleosome. Mol Cell 72, 594-600 e592.
Marchand, J.R., and Caflisch, A. (2015). Binding Mode of Acetylated Histones to
Bromodomains: Variations on a Common Motif. ChemMedChem 10, 1327-1333.
McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Winn, M.D., Storoni, L.C., and Read,
R.J. (2007). Phaser crystallographic software. J Appl Crystallogr 40, 658-674.
Plotnikov, A.N., Yang, S., Zhou, T.J., Rusinova, E., Frasca, A., and Zhou, M.M. (2014).
Structural insights into acetylated-histone H4 recognition by the bromodomain-PHD
finger module of human transcriptional coactivator CBP. Structure 22, 353-360.
Pokholok, D.K., Harbison, C.T., Levine, S., Cole, M., Hannett, N.M., Lee, T.I., Bell, G.W.,
Walker, K., Rolfe, P.A., Herbolsheimer, E., et al. (2005). Genome-wide map of
nucleosome acetylation and methylation in yeast. Cell 122, 517-527.
Ramachandran, S., Zentner, G.E., and Henikoff, S. (2015). Asymmetric nucleosomes
flank promoters in the budding yeast genome. Genome Res 25, 381-390.
Rawal, Y., Chereji, R.V., Qiu, H., Ananthakrishnan, S., Govind, C.K., Clark, D.J., and
Hinnebusch, A.G. (2018). SWI/SNF and RSC cooperate to reposition and evict
promoter nucleosomes at highly expressed genes in yeast. Genes Dev 32, 695-710.
Ruthenburg, A.J., Li, H., Patel, D.J., and Allis, C.D. (2007). Multivalent engagement of
chromatin modifications by linked binding modules. Nat Rev Mol Cell Biol 8, 983-
994.
Schrodinger, LLC (2015). The PyMOL Molecular Graphics System, Version 1.8.
Seo, H.S., Blus, B.J., Jankovic, N.Z., and Blobel, G. (2013). Structure and nucleic acid
binding activity of the nucleoporin Nup157. Proc Natl Acad Sci U S A 110, 16450-
16455.
Singh, M., Popowicz, G.M., Krajewski, M., and Holak, T.A. (2007). Structural
ramification for acetyl-lysine recognition by the bromodomain of human BRG1
protein, a central ATPase of the SWI/SNF remodeling complex. Chembiochem 8,
1308-1316.
VanDemark, A.P., Kasten, M.M., Ferris, E., Heroux, A., Hill, C.P., and Cairns, B.R.
(2007). Autoregulation of the rsc4 tandem bromodomain by gcn5 acetylation. Mol
Cell 27, 817-828.
Vidler, L.R., Brown, N., Knapp, S., and Hoelder, S. (2012). Druggability analysis and
structural classification of bromodomain acetyl-lysine binding sites. J Med Chem 55,
7346-7359.
37You can also read

















































