Spatial and temporal variations of the subtidal meiobenthic community of the central coast of Venezuela Variación espacial y temporal de la ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Spatial and temporal variations of the subtidal
meiobenthic community of the central coast of
Venezuela
Variación espacial y temporal de la comunidad
meiobentónica submareal de la costa central de
Venezuela
Carlos Pereira1*, Evelyn Zoppi De Roa1†, Vanessa Hernández1, María Angélica Crespo2,
Joxmer Scott-Frías1 and Edie Montiel1
ABSTRACT
Meiobenthos is a sediment-dwelling community with various taxonomic groups ranging in size
from 63 to 500 µm. As a matter and energy link between primary producers and macrofauna,
they have other functions in the ecosystem. Since this group has been scarcely studied in
Venezuela, it was proposed to analyze the spatial and temporal variations in the composition and
abundance of subtidal meiobenthos in the country’s central coast. Samplings were conducted
monthly from June 2014 to March 2015. In each sampling campaign, 12 random stations were
established, and three samples were taken at each using a cylindrical sampler of 5.0 cm internal
diameter. Organisms were identified and counted using microscopes, and data was processed
using hypothesis tests (ANOSIM) and multivariate analyzes (SIMPER, nMDS). Meiobenthos
showed a richness of 164 morphotypes and 14 phyla, with a dominance of planktonic copepods
and foraminifers, followed by cnidarians and mollusks. Out of the total species reported, 84
are strictly benthic. Regarding abundance, there was a greater representation of foraminifera,
followed by nematodes and mollusks, with values higher than those reported for tropical areas.
Spatially, no differences were found in the composition and abundance of this community.
However, two maximums were found throughout the study period, one starting in June until
reaching maximum densities between August and October and the second one in February.
Keywords: Caribbean Sea, community structure, marine ecology, meiobenthos, meiofauna
1 Laboratorio de Ecología de Sistemas Acuáticos, Instituto de Zoología y Ecología Tropical, Universidad Central de
Venezuela. Caracas, Venezuela. carlosjpereyra2@gmail.com* ORCID: http://orcid.org/0000-0002-5850-0775 OR-
CID: http://orcid.org/0000-0003-2659-0857 ORCID: http://orcid.org/0000-0001-9557-6333 ORCID: https://orcid.
org/0000-0002-0901-6768
2 Escuela de Ciencias Aplicadas del Mar, Universidad de Oriente. Isla de Margarita, Venezuela. ORDIC: https://orcid.
org/0000-0003-1336-0972
Recepción 07 septiembre 2020 • Corregido 05 diciembre 2020 • Aceptado 07 diciembre 2020
DOI: http://dx.doi.org/10.15359/revmar.12-2.5 95
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020 Licencia Creative Commons
Atribución-No-Comercial
ISSN: 1659-455X • e-ISSN: 1659-407X Compartir Igual 4.0 Costa Rica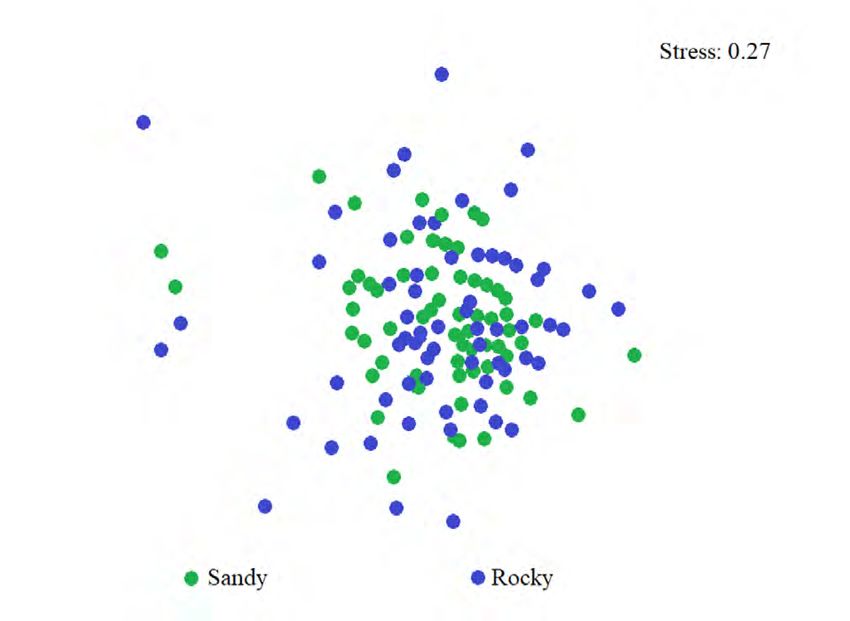
Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
RESUMEN
El meiobentos es una comunidad que habita en los sedimentos, tiene una variedad de grupos
taxonómicos con tamaños entre 63 y 500 µm. Representan un vínculo de materia y energía entre
los productores primarios y la macrofauna, con otras funciones para el ecosistema. En Venezuela,
este grupo ha sido escasamente estudiado, por lo cual se planteó analizar las variaciones espaciales
y temporales de la composición y abundancia del meiobentos submareal en la zona costera central
del país. Los muestreos fueron realizados, por mes, desde junio de 2014 hasta marzo de 2015.
En cada campaña de muestreo, se establecieron 12 estaciones aleatorias y, en cada una de ellas,
se obtuvieron 3 muestras, con el uso de un dispositivo cilíndrico de 5 cm de diámetro interno.
La identificación y conteo de los organismos se realizó con microscopio. Para el análisis de los
resultados, se hicieron pruebas de hipótesis (ANOSIM) y análisis multivariados (SIMPER, nMDS).
La meiofauna mostró una riqueza de 164 morfotipos y 14 phyla, con dominancia de copépodos
planctónicos y foraminíferos, seguidos de cnidarios y moluscos. De estas especies, 84 eran
estrictamente bentónicas. En cuanto a la abundancia, hubo mayor representación de foraminíferos,
seguido de nemátodos y moluscos, con valores superiores a los reportados para áreas tropicales.
Con respecto al espacio, no se encontraron diferencias en la composición y abundancia de esta
comunidad. Sin embargo, temporalmente, se hallaron dos máximos, uno que inició en junio, que
alcanzó las densidades máximas entre agosto y octubre, y el segundo en febrero.
Palabras claves: ecología marina, estructura comunitaria, mar Caribe, meiobentos, meiofauna
INTRODUCTION The meiobenthos community
resides in interstitial spaces of sedi-
In marine sediments, countless ments and above other organisms in
invertebrate species coexist and interact all aquatic habitats, from the intertidal
with each other, making up a complex to the deep ocean areas in all latitudes
trophic network (Giere, 2009). These (Wołowicz et al. 2011; Dupuy et al.
organisms could be classified based 2015). Marine meiobenthos include
on their size. In this work, we used the more than 30 taxonomic groups, of
definition of meiobenthos from Coull which nematodes, foraminifera, har-
& Bell (1979) and Giere (2009) that pacticoid copepods, cumaceans, ostra-
groups aquatic intermediate-sized ben- cods, platyhelminthes, annelids, and
thic organisms and includes taxonom- juvenile stages of other groups are the
ically diverse metazoans and proto- most abundant (Giere, 2009; Woło-
zoa. In fact, those organisms that pass wicz et al. 2011; Dupuy et al. 2015;
through a 500 µm mesh-size sieve and Warwick, 2018).
are retained on a 63 µm mesh, includ- Meiobenthos play a modulating
ing permanent and temporary small role as a trophic link to the macro-
metazoans and larger protozoans (cil- fauna. Secondary production in shal-
iates, amoebozoans), are considered low aquatic systems is highly depen-
meiobenthos (Giere, 2009). dent on them because they transfer
96 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5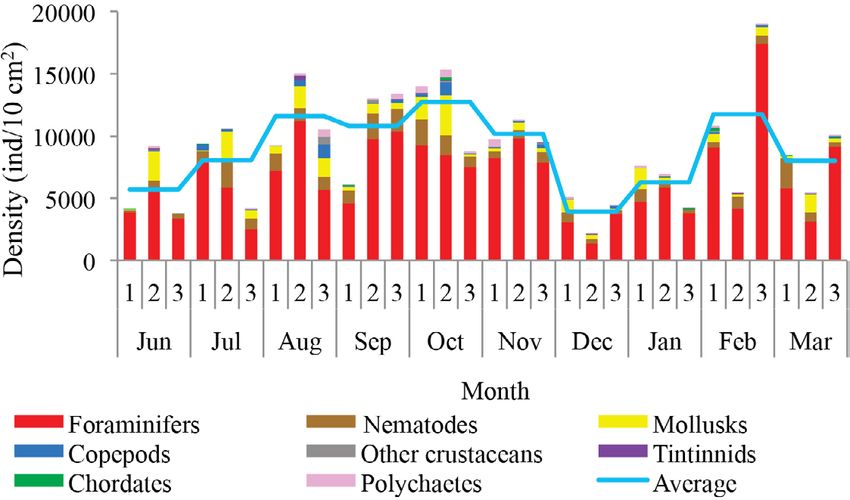
Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
the energy produced by microalgae MATERIALS AND METHODS
and bacteria to benthic consumers of
higher trophic levels (Moncreiff & Study area
Sullivan, 2001; Ansari et al. 2012). The study area is located in the
Meiobenthos modify the physical, north-central continental region of Ve-
chemical, and biological properties of nezuela, at the end of the foothills of
sediments, participating in sediment Cordillera de la Costa. It includes the
stabilization, biogeochemical cycles, populations from Chirimena to Puerto
and waste removal (Wang et al. 2019). Francés in the state of Miranda (Fig.
Additionally, it has been widely used 1). It is a heterogeneous area with
for monitoring environmental quality deep and shallow coasts influenced by
due to its fast response to changes in rivers and ephemeral streams, such as
the environment, species richness, and the Tuy river and the Tacarigua coastal
short life cycles with 3 to 5 genera- lagoon (Herrera & Bone, 2011). This
tions per year (Giere, 2009; Ansari et area is characterized by a dry tropical
al. 2012; Kitahashi et al. 2018). climate, according to Köppenˈs clima-
In Venezuela, only a few studies te classification, with an average tem-
have been conducted on this commu- perature around 29.1°C. Annual preci-
nity. Woods & Tietjen (1985) analyzed pitation is 1,327 mm with the highest
the meiobenthos of the Venezuelan ba- precipitation in November (Schwarz,
sin; Reyes et al. (1999) studied the ben- 2014). During the first months of the
thic community in the western side of year, coastal upwelling occurs, which
the country; Martín et al. (2007) con- keeps the waters fertilized during the
ducted a baseline study in the eastern dry season (Castellanos et al. 2002).
part of the country; Vásquez-Suárez et Regarding marine currents, the area
al. (2010) analyzed meiofauna tempo- presents a current pattern with an East-
ral variation in Raya Lagoon on Mar- West direction (Pereira et al. 2020).
garita Island; and Liñero et al. (2013) Sampling design
studied the spatial and temporal va- Samples were taken in each zone
riation of the subtidal meiofauna on from a multistage cluster design with
a beach in the eastern zone. For this subsamples. A 12 km coastline was es-
reason, this study proposed to descri- tablished and divided into three zones
be the spatial and seasonal variations separated by areas with sandy bottoms
of the composition and abundance of of 500 m approximately. For purposes
subtidal meiobenthos in the central of this study, these sandy areas were
coast of Venezuela. considered barriers between zones.
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 97
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5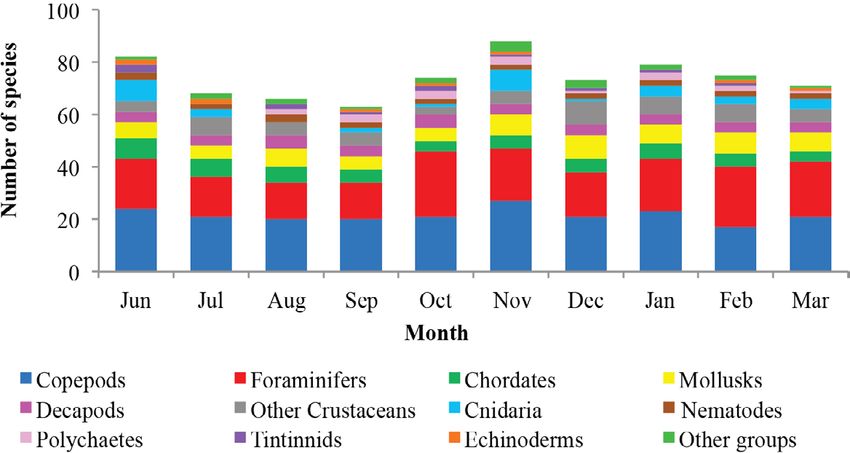
Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
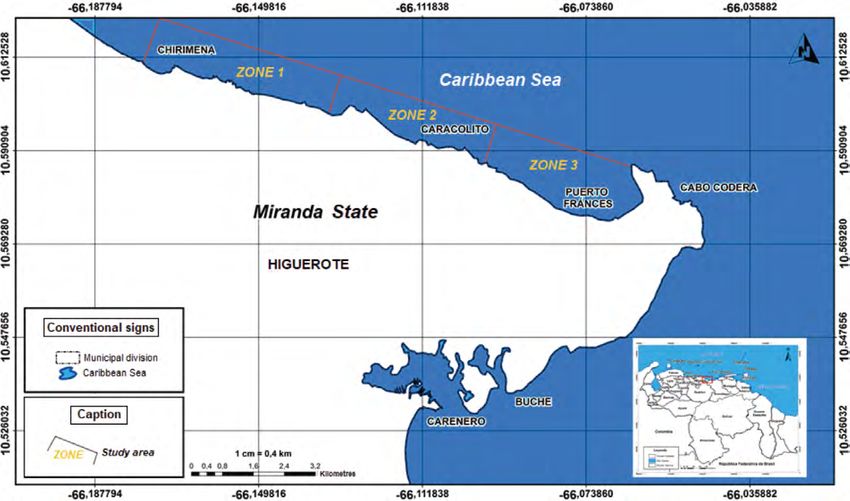
Fig. 1. Location of the sampling zones in the coastal area between Chirimena and
Puerto Francés, Venezuela
Fig. 1. Ubicación de las zonas de muestreo en el área costera entre Chirimena y Puerto
Francés, Venezuela
Zones were called Chirimena (10°36’ The first one was creating a grid on a
42’’ N, 66°10’ 37.15’’ W - 10°36’ 1.3’’ map with five rows and five columns
N 66°8’ 8.3’’ W), Caracolito (10°36’ in each sampling zone. Each cell had
1.3’’ N 66°8’ 8.3’’ W - 10°35’30.7’’ a dimension of 50 meters long towards
N 66°6’6.7’’ W), and Puerto Francés the sea and 870 meters along the coast
(10°35’ 30.7’’ N 66°6’ 6.7’’ W - 10°34’ with depths from 5 to 25 m. The se-
52’’ N 66°3’ 40.8’’ W) (Fig. 1). cond one refers to the bottom type, viz.
Sampling campaigns were con- sand and rock. Each cell was assigned
ducted monthly from June 2014 to a number, and sampling sites were
March 2015 at the three zones men- chosen monthly through a random
tioned above (Chirimena, Caracolito, number selection system.
and Puerto Francés). In each sampling Sampling
zone, four stations were randomly se- Sediment samples were collec-
lected every month, resulting in a total ted at each station by SCUBA divers,
of 12 stations per month (Fig. 1). Two using a 0.25 m2 quadrant divided into
criteria were established for the ran- 25 quadrants of the same size. Three (3)
domization of the sampling stations. random samples were taken inside the
98 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5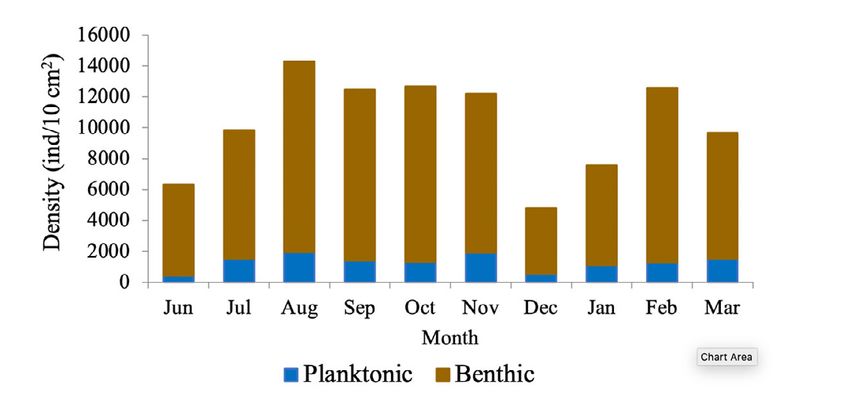
Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
quadrant, using a cylindrical corer of 5 to species and others to family, gene-
cm inner diameter slowly buried about ra, or morphotype. The validity of the
1 cm in sandy substrates. The sampler scientific names was verified at mari-
was then covered to obtain a layer of nespecies.org (WoRMS, 2020).
sediment with the inhabiting organisms Statistical analysis
using suction. In the case of rocky subs- Monthly data on the number of
trates, the sediment surface was scraped species or morphotypes, density, and
with the same sampler with circular mo- Shannon-Wiener biological diversity
vements at the rock bottom. The sam- of the meiobenthic community at three
pler was quickly covered on both sides. zones at the Caribbean Sea is presented,
On the boat, samples were fixed with a using descriptive statistical techniques.
5% v/v solution of buffered formalin. As part of the community’s characteri-
Laboratory analysis of samples zation, hypothesis tests were performed
Sediment samples were was- from a bifactorial model (month and
hed with distilled water through a set zone) without interactions, using a si-
of 500 and 63 μm sieves. The filtered milarity analysis (ANOSIM). In cases
material was washed eight times insi- where significant differences were found
de the 63 µm mesh sieve and stained with a type I error (α) = 0.05, subsequent
with Rose Bengal. Organisms were tests (Less Significant Differences, LSD)
identified and counted directly, obser- were applied. A SIMPER test based on
ving the sediment under a Leica MZ95 abundance was performed to determi-
microscope with a 400X magnification ne the percentage of similarity between
in the Ecology of Aquatic Systems sample groups according to the time and
laboratory at the Central University space factors. Non-metric multidimen-
of Venezuela. The works of Willey sional scaling (nMDS) ordering analyzes
(1930, 1932, 1935), Day (1967), were applied to visualize the pattern of
Owre & Foyo (1967), Trégouboff & similarity of the samples (Clarke et al.
Rose (1967a;b), Wells (1978, 1979), 2014). The Bray-Curtis similarity index
Platt & Warwick (1983; 1998), Fiers was used for both analyzes. Statistical
(1984, 1986a, b, 1995), Todd & La- analyzes were performed with the PAST
verack (1991), Suárez-Morales & 4.03 software (Hammer et al. 2001).
Gasca (1997), Warwick et al. (1998),
Conway (2012a,b), and Johnson & RESULTS
Allen (2012), as well as the Nemys
database (Bezerra et al. 2020) were Total community composition
used as references to identify sam- Meiobenthos of the marine and
ples. Organisms were identified to the coastal zone between Chirimena and
lowest possible taxonomic level, some
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 99
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5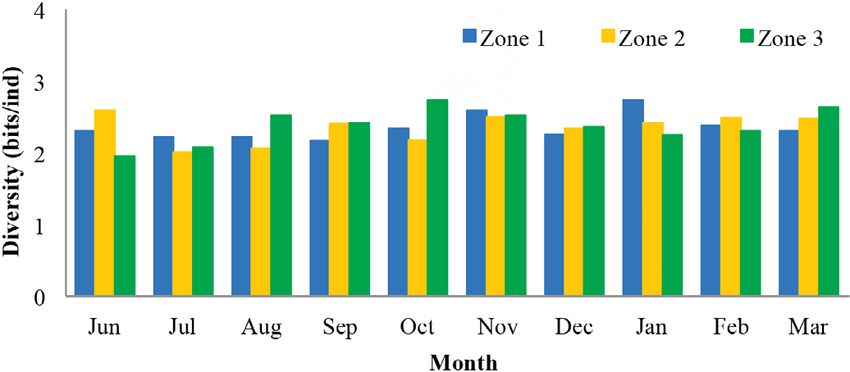
Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
Puerto Francés was represented by 164 84 species of the total reported are
species or morphotypes and 14 phyla. considered strictly benthic (51%). The
The phylum with the highest number rest of the species belonged to plank-
of species was Arthropoda with 69 ton and were found in the sediment
morphotypes represented by 48 spe- samples at very low densities (Fig. 2).
cies of copepods (representing 29.3% Total densities of meiobenthos
of the total richness), 8 decapods, 3 varied between 1,230 and 23,058
cladocerans, 3 amphipods, 2 isopods, ind/10 cm2 with an average density of
2 cirripeds, 1 stomatopod, 1 ostracod, 8,900 ± 4,037 ind/10 cm2. Regarding
and 1 mysid. Other important groups the density by taxonomic groups, a
were Foraminifera with 34 species greater contribution of foraminiferans
(20.7%), Cnidaria with 16 (9.8%), was observed with an average densi-
Mollusca with 13 (7.9%), Chordata ty of 6,729 ± 3,187 ind/10 cm2 that
with 9 (5.5%), and Polychaeta with represented approximately 76.2% of
6 (3.7%). Moreover, other taxonomic the total community. It was followed
groups with less than 5 species were by nematodes with 967 ± 609 ind/10
identified, belonging to the phyla cm2 (11%), mollusks with 739 ± 551
Bryozoa, Chaetognata, Ciliophora, ind/10 cm2 (8.4%), and copepods,
Echinodermata, Nematoda, Phoroni- mainly harpacticoid, with 346 ± 379
da, and Radiozoa, which together rep- ind/10 cm2 (3.9%). The most abun-
resented 11% of the total species. Only dant species were Neogloboquadrina
sp., Globorotalia me-
nardii, Globorotalia
scitula, Quinquelocu-
lina sp., Woodringina
sp., and Globigerina
sp. foraminiferans,
Euchromadora vul-
garis and Epsilonema
sp. nematodes, the
Fig. 2. Monthly variation of the mean density of meiobenthos pteropod Limacina
in the coastal zone between Chirimena and Puerto Francés, helicina, an uniden-
June 2014 - March 2015, showing the proportion of planktonic tified gastropod, and
and benthic species several genera and
Fig. 2. Variación mensual de la densidad media del meiobentos species of harpacti-
de la zona costera entre Chirimena y Puerto Francés entre coid copepods such
junio 2014 y marzo 2015, mostrando la proporción de las
as Euterpina acuti-
especies planctónicas y bentónicas
frons, Harpacticus
100 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
spp., Longipedia sp., Microsetella ro- difference was found between zones
sea, Paralaophonte sp., Peltidium sp., (R = 0.133; P = 0.1). An average
Phyllopodopsyllus sp., Pseudobradya density of 8,341 ± 3,087 ind/10 cm2
sp., and Tisbe spp. was obtained in zone 1 (Chirimena),
Spatial variability 9,029 ± 3,968 ind/10 cm2 in zone 2
No remarkable differences were (Caracolito), and 8,993 ± 3,965 ind/10
observed in the spatial richness of the cm2 in zone 3 (Puerto Francés). The
three established zones (ANOSIM: proportion of taxonomic groups was
R = 0.146; P = 0.07). Richness was similar between zones, where nem-
between 121 and 124 with the same atodes and mollusks were the ones
proportion of taxonomic groups. Re- generating these slight differences.
garding the substrate type, greater The SIMPER test showed an aver-
richness was found in the sandy sub- age similarity among the samples of
strate with 146 species compared to 27.71% in zone 1, 28.34% in zone 2,
the rocky substrate with only 133 spe- and 23.47% in zone 3. The species
cies. Although the proportion of tax- contributing over 70% to this similar-
onomic groups was similar, the sandy ity were the foraminiferans Globoro-
substrate had particular species, in- talia menardii, Globorotalia scitula,
cluding some planktonic species not and Neogloboquadrina sp., and the
found in the rocky substrate. Strictly nematode Euchromadora vulgaris.
considering the meiobenthic species, Additionally, a one-way ANOSIM
the following species were found in test was performed to compare the spe-
the sandy substrate: Foraminifera Am- cies composition between the two types
monia sp., Baculogypsina sphaeru- of substrates, and no significant differen-
lata, and two species of the order ces were found (ANOSIM, R = 0.033;
Rotaliida, the harpacticoid copepods P = 0.3). This result was confirmed with
Ameira trisetosa and Laophonte sp., the nMDS, which showed a single point
an amphipod of the genus Elaspomus cloud (Fig. 3). Using the SIMPER test, a
sp., three mollusks, two polychaetes, 30.79% similarity was obtained among
and an unidentified Phoronidae. The sandy substrate samples, 22.34% among
following species were found in the rocky substrate samples, and 25.5%
rocky substrate: the tintinnid Undella between both substrates. The foramini-
hyalina and the copepods: Tisbe spp. ferans Globigerina sp2., Globorotalia
and Robertsonia sp. menardii, Globorotalia scitula, Neoglo-
Using a two-way nested ANO- boquadrina sp., and Woodringina spp.,
SIM test to evaluate species compo- one gastropod, the nematode Euchroma-
sition (in which zones were includ- dora vulgaris, and several Calanoid co-
ed within months), no significant pepods contributed with more than 70%
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 101
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
to these similarities.
Regarding the biolo-
gical diversity index,
the lowest diversity
values were estimated
in zone 1 with an ave-
rage of 2.5 bits/ind,
while zones 2 and 3
had both a mean value
of 2.6 bits/ind (Fig. 4).
Temporal
variability
Regarding the
Fig. 3. Non-metric multidimensional scaling analysis (nMDS) total temporal vari-
of the total abundance of meiobenthos in sandy substrate ability of richness,
(green) and rocky substrate (blue) in the coastal zone between two maximums were
Chirimena and Puerto Francés, June 2014 - March 2015 observed: one in
Fig. 3. Análisis de ordenación de escalamiento multidimensional June, in which 86
no métrico (nMDS) de la abundancia total del meiobentos en species were iden-
sustrato arenoso (verde) y rocoso (azul) de la zona costera entre tified, and the other
Chirimena y Puerto Francés entre junio 2014 y marzo 2015 in November with
92 identified species.
These peaks coin-
cided with the tran-
sitions between the
rainy (August-No-
vember) and dry
(January-March)
seasons. Samples
presented a similar
Fig. 4. Shannon biological diversity index of meiobenthos by proportion of species
zone in the coastal zone between Chirimena and Puerto Francés, with some excep-
June 2014 – March 2015. Zone 1 (blue): Chirimena, Zone 2 tions. For example,
(yellow): Caracolito, and Zone 3 (green): Puerto Francés. the copepods pro-
Fig. 4. Índice de diversidad biológica de Shannon del meiobentos portion varied from
por zonas en la zona costera entre Chirimena y Puerto Francés 27 to 30%, except
entre junio 2014 y marzo 2015. Zona 1 (azul): Chirimena, zona 2 in February, when
(amarillo): Caracolito y zona 3 (verde): Puerto Francés it decreased to 22%
102 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
with 17-24 species. Foraminifera pre- February with 10,478 ± 4,832 ind/10
sented percentages between 20 and cm2. The lowest density was obtained
22%, except in October, February, in December with 3,991 ± 1,555 ind/10
and March when their proportion in- cm2 (Fig. 6). The two-way nested
creased, varying between 27 and 32%. ANOSIM test showed significant dif-
Foraminifera richness varied between ferences between months (R = 0.587;
14 species in July, August, and Sep- P = 0). The post-test to compare each
tember and 25 species in October. month indicated differences between
Another group that showed varied July/November, July/January, July/
proportions between months was Cni- February, July/March, August with
daria, going from 0 species in August the months between November and
to 8 species in June and November, March, September/December, Sep-
influencing the temporal variation of tember/January, September/March,
total richness. The rest of the groups October/November, October/January,
remained somewhat constant (Fig. 5). as well as between December, January,
Regarding temporal variability February, and March.
of density, two maximum densities As far as the SIMPER test,
were observed, the first between Au- samples within each month present-
gust and November (rainy season) ed different composition similari-
with an average density of 10,825 ties with an undefined pattern. The
± 2,898 ind/10 cm2 and the other in highest similarity values (>35%)
occurred in July,
September, and
October, and the
lowest was in
June (12%). Re-
garding the sim-
ilarity between
months, it was
found that June
presented a sim-
ilarity with the
Fig. 5. Monthly variation of the richness of meiobenthos in the rest of the months
coastal zone between Chirimena and Puerto Francés, June 2014 ranging from 9
- March 2015 to 12.1%, July
Fig. 5. Variación mensual de la riqueza del meiobentos de la 12.1 - 29.8%, Au-
zona costera entre Chirimena y Puerto Francés entre junio 2014 gust 9.7 - 26%,
y marzo 2015 September 10.7
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 103
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
DISCUSSION
In this study,
a diverse meioben-
thic community with
a significant num-
ber of copepod spe-
cies characteristic
of the zooplankton
community was ob-
served in the central
Fig. 6. Spatial and temporal variation of the mean density coast of Venezuela,
of meiobenthos in the coastal zone between Chirimena and from the Chimin-
Puerto Francés, June 2014 - March 2015 era region to Puerto
Fig. 6. Variación temporal y espacial de la densidad media Francés. Although
del meiobentos de la zona costera entre Chirimena y Puerto their densities were
Francés entre junio 2014 y marzo 2015 low, their presence
in the meiobenthos
- 27.5%, October 9.7 - 21.6%, No- could indicate the link between the
vember 11.5 - 27.5%, December 9.6 planktonic and benthic communi-
- 23.6%, January 9 - 24.5%, Feb- ties, where these planktonic organ-
ruary 10.9 - 29.8%, and March 9.3 isms migrate to the bottom to feed on
- 24.9%. Some foraminifera, nema- benthic organisms (Schnack-Schiel
todes, and mollusks species contrib- & Isla, 2005). As in this study, Liñe-
uted in a greater proportion to this ro et al. (2013) identified 14 taxa on
similarity. a beach in eastern Venezuela. The au-
The Shannon Diversity Index thors excluded foraminifera from their
showed temporal variation with va- analysis, and found a dominance of
lues between 1.8 (low diversity) and nematodes with an average density of
3.1 bits/ind (high diversity). Two di- 155.80 ± 29.07 ind/10 cm2 that repre-
versity peaks were observed, the first sented 58.36%, followed by ostracods
one in October with 2.9 bits/ind and with 45.60 ind/10 cm2 (17.10%), and
the second one between February and harpacticoid copepods with 41.96
March with 2.7 bits/ind. The lowest ind/10 cm2 (15.7%).
mean diversity value was 2.2 bits/ind Liñero et al. (2013) explained
in June and December (Fig. 4). the difficulty of making comparisons
with other studies due to the method-
ological differences that exist during
104 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
sampling, data analysis, and interpre- and French Guiana (Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
Páez et al. (2001) indicated that total abundance. These researchers
foraminifera represent the most abun- reported copepods with a density be-
dant group of organisms in the marine tween 11.57 and 54 ind/10 cm2, which
meiofauna with more than 5,000 spe- represented about 15% of total abun-
cies, of which benthic species are 30 dance, and foraminifera with densities
times more numerous than the plank- between 16 and 47.29 ind/10 cm2, rep-
tonic species. These protists inhabit resenting 14% of the abundance. How-
all depths of the seas, demonstrating ever, unlike their study (Ansari et al.
a high level of efficiency and ecolog- 2012), ours did find mollusks.
ical adaptability to different substrates The nematode density estimated
(Páez et al. 2001, Sabbatini et al. in this study (1,218 ± 628 ind/ 10cm2)
2014), which explains the high domi- coincided with the one estimated by
nance of foraminifera in this study. Vásquez-Suárez et al. (2010) in Raya
Other important groups in this lagoon (1,209 ind/ 10cm2) and by
study were nematodes with 11% of Díaz-Asencio et al. (2009) in Cien
total abundance and mollusks with an Fuegos bay, Cuba (1,212.5 ± 889.7 ind/
8.4% representation. Gourbault et al. 10cm2). Moreover, the nematode spe-
(1998) pointed out that nematodes are cies identified in this study coincided
the most dominant group in reef ar- with the most frequent and abundant
eas, which suggests their need to stay in the Cuban archipelago, such as Eu-
in more stable or protected areas. In chromadora vulgaris and Epsilonema
fact, Dye & Lasiak (1986) and Liñero sp. (Pérez-García et al. 2020). Another
et al. (2013) found that nematodes had important group in terms of abundance
preferences for fine sands. Although was harpacticoid copepods.
the sediments were not characterized A study conducted by Alfon-
in this study, this area is characterized so & López (2006) in several coastal
by being of high energy with mud- localities in the northern zone of Ha-
dy-sandy sediments (Solé & Vera, vana showed wide-ranging densities
1997, Pereira et al. 2020), which ex- between 892.12 and 7,133.48 ind/10
plains the abundance values found in cm2, excluding foraminifera, of which
this research. nematodes represented 49-90% of to-
Despite not being the most rep- tal abundance, while copepods repre-
resentative group, the density of nem- sented 51-66%, which are at least an
atodes in this research exceeded the order of magnitude higher than the
one obtained by Ansari et al. (2012) abundances obtained in this study.
in a coastal area in southeast India Soltwedel (2000) saw that in the ma-
with densities between 34.6 and 207.7 jority of oceanic areas, the proportion
ind/10 cm2, representing 57% of the of nematodes exceeds 75% of the total
106 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
meiobenthos, excluding foraminifera. in which the highest densities were
Only in the tropical areas (in western reached from October to December
India and northeast Australia), did and the lowest in February, July, and
nematodes constitute less than 60% August (Liñero et al., 2013).
of the total abundance of microscop- Although no correlations were
ic metazoan organisms (Soltwedel made in this study with environmen-
2000). In fact, an analysis of 621 sta- tal variables, some studies indicate a
tions worldwide showed an average trend to increase density during the
nematode density of 525.27 ind/10 dry season (Coull, 1970; Chindah &
cm2 and 1,116.25 ind/10 cm2 in 107 Braide, 2001). In the coastal area be-
samples at depths between 20 and 200 tween Chirimena and Puerto Francés,
m (Mokievskii et al. 2007). The results an increase was observed in density
obtained in this work are in the inter- during the rainy season and maximum
val reported by these authors. temperatures, coinciding with what
Although granulometry was not was found by Venekey et al. (2014) in
analyzed in this study, samples were an intertidal zone in Brazil, Zaleha et
collected in a heterogeneous area al. (2016) on Karah Island in China,
with rocky and sandy substrates, con- and Baia & Venekey (2019) on san-
sisting mainly of medium to coarse- dy beaches in Brazil. This could be
grained sand. As indicated by De- explained by the availability of food
cho et al. (1985), Armenteros et al. during the rainy season due to the en-
(2009), Vásquez-Suárez et al. (2010), richment of the marine environment
and Liñero et al. (2013) in their spa- by the discharge of nutrients from the
tial analyzes, the lowest meiobenthic adjacent rivers and streams.
densities were obtained on sandy Other variables that influence
beaches, while the highest were found the composition and distribution of
in finer grain sediments. In contrast, meiobenthos are salinity, water move-
Ruiz-Abierno & Armenteros (2016) ment, sediment characteristics, food
found no differences in the distribu- quality and availability, predation,
tion of meiobenthos depending on dispersal, biogenic structures, and bi-
the substrate, inferring that hydrody- ological disturbance (Edwards 2009;
namics nullified these differences as Giere, 2009; Baia & Venekey, 2019).
found in this study. Temporal patterns However, Alfonso & López (2006)
with two increases in density (Au- found that changes in the structure
gust-November and February-March) and composition of the meiofaunal
during this study period coincided communities were related to stochas-
with the one obtained by Liñero et al. tic events such as storm drains, tropi-
(2013). However, a gap was evident cal storms, and hurricanes.
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 107
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
In addition, a study conducted samples from the sandy substrate pre-
by Kumary (2016) about the temporal sented a greater number of species than
variation of the interstitial meiofauna the rocky substrate. Concerning abun-
in India indicated that some factors dance, there was an important repre-
such as ecological interactions be- sentation of foraminifera, followed by
tween species, organic matter content, nematodes and mollusks. Temporarily,
sediment grain size, and oxygen had two peaks were found. The first one
a considerable influence on the spa- began in June with progressive increa-
tial variation of the components of ses until reaching maximum densities
the meiobenthic community. While between August and October. Subse-
temporarily, taxonomic composition, quently, density gradually decreased
density, and distribution depend on from October to December, after which
increases in temperature and salinity, it started to climb until reaching the se-
the stability of sediments, and proba- cond maximum in February.
bly the availability of food favored by
pre and post-monsoon population enri- ACKNOWLEDGMENTS
chment. Therefore, subsequent studies
should estimate other variables related We would like to thank Petró-
to the characteristics of the sediment leos de Venezuela, S.A. for funding
and complement community studies the project and the reviewers for their
with population analysis of the modu- contributions to improve the quality of
lating species of this community. the manuscript.
CONCLUSIONS BIBLIOGRAPHY
These results represent a contri- Alfonso, H. & López, C. (2006). Distribución
bution to the knowledge on meioben- espacio-temporal de la meiofauna bén-
thos ecology in Venezuela and the Ca- tica en cuatro playas del Litoral Norte
de la Habana. Rev. Biol. Trop., 54(3),
ribbean Sea. Meiobenthos showed high 985-995. https://doi.org/10.15517/RBT.
richness with the presence of plankto- V54I3.14078
nic species and an outstanding repre- Ansari, K., Lyla, P. & Ajmal-Khan, S. (2012).
sentation of copepods and foraminifera, Faunal composition of metazoan meio-
followed by cnidarians and mollusks. fauna from the southeast continen-
tal shelf of India. Indian J. Mar. Sci.,
Temporally, two maximums in species 41(5), 457-467.
richness were observed, while spatially, Armenteros, M., Ruiz-Abierno, A., Fer-
no differences were observed between nández-Garcés, R., Pérez-García, J.,
the three established zones, although Díaz-Asencio, L., Vincx, M. & De-
craemer, W. (2009). Biodiversity
108 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
patterns of free-living marine nemato- Nemertea, Rotifera and Mollusca. In
des in a tropical bay: Cienfuegos, Ca- A. John (Ed.), Occasional Publications
ribbean Sea. Estuar. Coast. Shelf. Sci., (pp. 1-138). England: Marine Biological
85, 179-189. https://doi.org/10.1016/j. Association of the United Kingdom.
ecss.2009.08.002 Conway, D. (2012b). Marine zooplankton of
Avilés-Ramírez, G., González-León, J., Poot-Es- southern Britain. Part 2: Arachnida,
parza, A., Medina-García, M. & Ro- Pycnogonida, Cladocera, Facetotecta,
jas-González, R. (2016). Influencia de Cirripedia and Copepoda. In A. John
arrecifes artificiales reef ball de la ense- (Ed.), Occasional Publications (pp.
nada de Xpicob, sobre la estructura de la 1-163). England: Marine Biological
meiofauna y sedimentos recientes. Agro- Association of the United Kingdom.
productividad, 9(9), 44-49. Coull, B. (1970). Shallow water meiobenthos
Baia, E. & Venekey, V. (2019). Distribution off the Bermuda platform. Oecologia,
patterns of meiofauna on a tropical ma- 4, 325-357. https://doi.org/10.1007/
crotidal sandy beach, with special focus BF00393393
on nematodes (Caixa d’Água, Amazon Coull, B. (1999). Role of meiofauna in es-
Coast, Brazil). Braz. J. Oceanogr., tuarine soft-bottom habitats. Aust.
67, e19230. https://doi.org/10.1590/ J. Ecol., 24, 327-343. https://doi.
S1679-87592019023006701 org/10.1046/j.1442-9993.1999.00979.x
Bezerra, T., Decraemer, W., Eisendle-Flöck- Coull, B. & Bell, S. (1979). Perspectives
ner, U., Hodda, M., Holovachov, O., of marine meiofaunal ecology. In R.
Leduc, D., … & Vanreusel, A. (2020). Livingston (Ed.). Ecological Pro-
Nemys: World Database of Nemato- cesses in Coastal and Marine Sys-
des. http://nemys.ugent.be tems (pp. 189-216). USA: Plenum
Castellanos, P., Varela, R. & Müller-Karger, F. Publishing Corporation. https://doi.
(2002). Descripción de las áreas de sur- org/10.1007/978-1-4615-9146-7_10
gencia al sur del mar Caribe examinadas Day, J. (1967). A monograph on the polychae-
con el sensor infrarrojo AVHRR. Mem. ta of Southern Africa. England.: British
Soc. Cs. Nat. La Salle, 154, 55-76. Museum Natural History. https://doi.
Chindah, A. & Braide, S. (2001). Meiofauna org/10.5962/bhl.title.8596
occurrence and distribution in different Decho, A., Hummon, W. & Fleeger, J.
substrate types of bonny brackish wet- (1985). Meiofauna-sediment inte-
land of the Niger Delta. J. Appl. Sci. ractions around subtropical seagrass
Environ. Manag., 5, 33-41. https://doi. sediments using factor analysis. J.
org/10.4314/jasem.v5i1.54937 Mar. Res., 43, 237-255. https://doi.
Clarke, K., Gorley, R., Somerfield, P. & org/10.1357/002224085788437389
Warwick, R. (2014). Change in marine Díaz-Asencio, L., Armenteros, M., Díaz-Asen-
communities: An approach to statisti- cio, M., Fernández-Garcés, R., Gó-
cal analysis and interpretation. United mez-Batista, M. & Alonso-Hernández, C.
Kingdom: PRIMER-E, Plymouth Ma- (2009). Spatial and temporal variations of
rine Laboratory. meiofaunal communities in Cienfuegos
Conway, D. (2012a). Marine zooplankton of Bay, Cuba. Rev. Biol. Mar. Oceanogr.,
southern Britain. Part 1: Radiolaria, 44(1), 13-22. http://dx.doi.org/10.4067/
Heliozoa, Foraminifera, Ciliophora, S0718-19572009000100002
Cnidaria, Ctenophora, Platyhelminthes,
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 109
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
Dupuy, C., Thanh, H., Mizrahi, D., Jourde, J., Diagoniceps Willey. Bull. Inst. R. Sci.
Bréret, M., Arogué, H., … & Bocher, P. Nat. Belg., 65, 151-236.
(2015). Structure and functional charac- Giere, O. (2009). Meiobenthology - the mi-
teristics of the meiofauna community croscopic motile fauna of aquatic sedi-
in highly unstable intertidal mudbanks ments. Germany: Springer-Verlag.
in Suriname and French Guiana (North Gourbault, N., Warwick, R. & Helléouet, M.
Atlantic coast of South America). Cont. (1998). Spatial and temporal variabi-
Shelf Res., 110, 39-47. https://doi.or- lity in the composition and structure
g/10.1016/j.csr.2015.09.019 of meiobenthic assemblages (espe-
Dye, A. & Lasiak, T. (1986). Microbenthos, cially nematodes) in tropical beaches
meiobenthos and fiddler crabs: tro- (Guadeloupe, FWI). Cah. Biol. Mar.,
phic interactions in a tropical man- 39, 29-39. https://doi.org/10.21411/
grove sediment. Mar. Ecol. Prog. Ser., CBM.A.452BA5CD
32, 259-264. https://doi.org/10.3354/ Guzmán, H., Obando, V. & Cortés, J. (1987).
meps032259 Meiofauna associated with a Pacific
Edwards, C. (2009). Meiobenthos of the dis- coral reef in Costa Rica. Coral Reefs,
covery Bay Lagoon, Jamaica, with an 6, 107-112. https://doi.org/10.1007/
emphasis on nematodes. (Unpublished BF00301379
doctoral thesis). University of London, Hammer, O., Harper, D. & Ryan, P. (2001).
England. PAST: Paleontological statistics sof-
Fiers, F. (1984). Harpacticoid copepods from tware package for education and data
the West Indian Islands: Canuellidae analysis. Palaeontologia Electronica,
and Longipediidae (Copepoda, Har- 4(1), 1-9. http://palaeo-electronica.
pacticoida). Bijdragen tot de Dier- org/2001_1/past/issue1_01.htm
kunde, 54(2), 197-210. https://doi. Hansen, J., Alongi, D., Moriarty, D. & Pollard,
org/10.1163/26660644-05402006 P. (1987). The dynamics of benthic mi-
Fiers, F. (1986a). Harpacticoid cope- crobial communities at Davies Reef,
pods from the West Indian Islands: central Great Barrier Reef. Coral Re-
Laophontidae (Copepoda, Harpacti- efs, 6, 63-70. https://doi.org/10.1007/
coda). Amsterdam Expedition to the BF00301375
West Indian Islands, Report 48. Bijd. Herrera, A. & Bone, D. (2011). Influence
Dierkunde, 56, 132-164. https://doi. of riverine outputs on sandy bea-
org/10.1163/26660644-05601009 ches of Higuerote, central coast of
Fiers, F. (1986b). Harpacticoid copepods Venezuela. Lat. Am. J. Aquat. Res.,
from the West Indian Islands: Dar- 39(1), 56-70. https://doi.org/10.4067/
cythompsoniidae (Copepoda, Harpac- S0718-560X2011000100006
ticoida). Amsterdam Expedition to the Johnson, W. & Allen, D. (2012). Zooplankton of
West Indian Islands, Report 51. Bijd. the Atlantic and Gulf Coasts. A guide to
Dierkunde, 56, 282-290. https://doi. their identification and ecology. USA:
org/10.1163/26660644-05602008 The Johns Hopkins University Press.
Fiers, F. (1995). New Tetragonicipitidae Kitahashi, T., Kayama, H., Tsuchiya, M.,
(Copepoda, Harpacticoida) from the Yamamoto, H. & Yamamoto, H.
Yucatecan continental shelf (Mexi- (2018). A new method for acqui-
co), including a revision of the genus ring images of meiobenthic images
110 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
using the FlowCAM. MethodsX, 5, Colombia. Arq. Ciên. Mar., Fortaleza,
1330-1335. https://doi.org/10.1016/j. 44(1), 81-96.
mex.2018.10.012 Owre, H. & Foyo, M. (1967). Copepods of the
Kumary, A. (2016). Temporal variations in Florida Current with illustrated keys to
the distribution of interstitial meio- genera and species. USA: Institute of
fauna along the southwest coast Marine Science, University of Miami.
of India. J. Climatol. Weather Fo- Páez, M., Zúñiga, O., Valdés, J. & Ortlieb L.
recast., 4(3), 178-141. https://doi. (2001). Foraminíferos bentónicos re-
org/10.4172/2332-2594.1000178 cientes en sedimentos micróxicos de
Liñero, I., Ojeda, S. & Amaro, M. (2013). Va- la bahía Mejillones del Sur (23º S),
riación espacio-temporal de la meio- Chile. Rev. Biol. Mar. Oceanogr., 36,
fauna submareal en una playa are- 129-139. http://dx.doi.org/10.4067/
nosa nororiental de Venezuela. Rev. S0718-19572001000200002
Biol. Trop., 61(1), 59-73. https://doi. Pereira, C., Fernández, A., Hernández, V. &
org/10.15517/RBT.V61I1.10884 Troccoli-Ghinaglia, L. (2020). Ocu-
Martín, A., Malavé, L., Sánchez, D., Aparicio, rrencia de floraciones nocivas de mi-
R., Arocha, F., Bone, D., … & Zoppi, croalgas en las costas del estado Mi-
E. (2007). Línea base ambiental Pla- randa, Venezuela. Cienc. Amb. Clima,
taforma Deltana. Venezuela: Petróleos 3(1), 55-71. https://doi.org/10.22206/
de Venezuela, S. A., Universidad Si- cac.2020.v3i1.pp55-71
món Bolívar. Pérez-García, J., Ruiz-Abierno, A. & Armen-
Mokievskii, V., Udalov, A. & Azovskii, A. (2007). teros, M. (2020). A checklist of aquatic
Quantitative distribution of meiobenthos nematodes from Cuban Archipelago.
in deep-water zones of the world ocean. Zootaxa, 4731(3), 301-320. https://doi.
Oceanology, 47(6), 797-813. https://doi. org/10.11646/zootaxa.4731.3.1
org/10.1134/S0001437007060057 Platt, H. & Warwick, R. (1983). Free-living
Moncreiff, C. & Sullivan, M. (2001). Trophic marine nematodes. Part 1. British Eno-
importance of epiphytic algae in sub- plids. England: The Linnean Society
tropical seagrass beds: Evidence from of London, The Estuarine and Bracki-
multiple stable isotope analyses. Mar. sh-Water Sciences Association.
Ecol. Prog. Ser., 215, 93-106. https:// Platt, H. & Warwick, R. (1988). Free-living
doi.org/10.3354/meps215093 marine nematodes. Part 2. Briti-
Netto, S., Attrill, M. & Warwick, R. (2003). sh Chromadorids. Netherlands: The
The relationship between benthic fau- Linnean Society of London, The Es-
na, carbonate sediments and reef mor- tuarine and Brackish-water Sciences
phology in reef-flat tidal pools of Rocas Association.
Atoll (north-east Brazil). J. Mari. Biol. Reyes, J., Godoy, A. & Casler, C. (1999). In-
Assoc. UK, 83, 425-432. https://doi. vertebrados bentónicos del bosque de
org/10.1017/S0025315403007288h manglar de la ciénaga de Los Olivitos,
Osorio-Dualiby, D. & Álvarez-León, R. Venezuela. Bol. Centro Invest. Biol.,
(2011). Characterization of meioben- 33(3), 227-241.
thos in areas of coastal influence by Ruiz-Abierno, A. & Armenteros, M. (2016).
two companies located in the indus- Coral reef habitats strongly influence
trial zone of Mamonal, Cartagena Bay, the diversity of macroand meioben-
thos in the Caribbean. Mar. Biodiv.,
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 111
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Carlos Pereira, Evelyn Zoppi De Roa, Vanessa Hernández,
María Angélica Crespo, Joxmer Scott-Frías and Edie Montiel
47, 101-111. https://doi.org/10.1007/ Vásquez-Suárez, A., González, M., Díaz, O.
s12526-016-0553-7 & Liñero, I. (2010). Variación tempo-
Sabbatini, A., Morigi, C., Nardelli, M. & ral de la meiofauna en sedimentos del
Negri, A. (2014). Foraminifera. In S. sistema lagunar “Laguna de Raya”, es-
Goffredo & Z. Dubinsky (Eds.). The tado Nueva Esparta, Venezuela. Inter-
Mediterranean Sea: Its history and ciencia, 35(2), 144-150.
present challenges (pp. 237-256). Venekey, V., Parreira, P. & Gomes, V. (2014).
England: Springer Science. https://doi. The influence of tidal and rainfall cycles
org/10.1007/978-94-007-6704-1 on intertidal nematodes: a case study in a
Schnack-Schiel, S. & Isla, E. (2005). The role tropical sandy beach. Braz. J. Oceanogr.,
of zooplankton in the pelagic-ben- 62(4), 247-256. https://doi.org/10.1590/
thic coupling of the Southern Ocean. s1679-87592014061706204
Sci. Mar., 69(2), 39-55. https://doi. Wang, X., Liu, X. & Xu, J. (2019). Distribu-
org/10.3989/scimar.2005.69s239 tion patterns of meiofauna assemblages
Schwarz, T. (2014). AmbiWeb GmbH. Cli- and their relationship with environ-
ma Higuerote. https://es.climate-da- mental factors of deep sea adjacent to
ta.org/america-del-sur/venezuela/ the Yap Trench, Western Pacific Ocean.
estado-miranda/higuerote-27656/ Front. Mar. Sci., 6(735), 1-12. http://
Solé, M. & Vera, B. (1997). Caracterización doi.org/10.3389/fmars.2019.00735
de las macroalgas marinas bénticas en Warwick, R., Platt, H. & Somerfield, P. (1998).
la región Chirimena-Punta Caimán, Free-living marine nematodes. Part 3.
Edo. Miranda, Venezuela. Carib. J. British Monhysterids. England: The
Sci., 33(3-4), 180-190. Linnean Society of London, The Es-
Soltwedel, T. (2000). Metazoan meiobenthos tuarine and Brackish-Water Sciences
along continental margins: a review. Association, Cambridge University.
Progr. Oceanogr., 46, 59-84. https://doi. Warwick, R. (2018). The contrasting histories
org/10.1016/S0079-6611(00)00030-6 of marine and freshwater meiobenthic
Suárez-Morales, E. & Gasca, R. (1997). Co- research-result of differing life histo-
pépodos (Crustacea) de las aguas su- ries and adaptive strategies? J. Exp.
perficiales del Mar Caribe Mexicano Mar. Biol. Ecol., 502, 4-11. http://dx.
(mayo, 1991). Rev. Biol. Trop., 45, doi.org/10.1016/j.jembe.2017.05.008
1523-1529. Wells, J. B. J. (1978). Keys to aid in the identifi-
Todd, C. & Laverack, M. (1991). Coastal ma- cation of marine harpacticoid copepods.
rine zooplankton: a practical manual Amendment Bull. No. 1. Zoology Publi.
for students. USA: Cambridge Univer- Victoria Univ. Wellington., 70, 1-11.
sity Press. Wells, J. B. J. (1979). Keys to aid in the identifi-
Trégouboff, G. & Rose, M. (1967a). Manuel cation of marine harpacticoid copepods.
de planctonologie Méditerranéenne. Amendment Bull. No. 1. Zoology Publi.
Tome I, texto. France: Centre National Victoria Univ. Wellington., 72, 1-8.
de la Recherche Scientifique. Willey, A. (1930). Harpacticoid Copepo-
Trégouboff, G. & Rose, M. (1967b). Manuel da from Bermuda. Part. I. Ann. Mag.
de planctonologie Méditerranéenne. Nat. Hist., 6, 81-114. https://doi.
Tome II, planches. France: Centre Na- org/10.1080/00222933008673192
tional de la Recherche Scientifique. Willey, A. (1932). Copepod phenology. Ob-
servations based on new material from
112 Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020.
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5Spatial and temporal variations of the subtidal meiobenthic
community of the central coast of Venezuela
Canada and Bermuda. Atti del Con- WoRMS Editorial Board. (2020). World Re-
gresso Internazionale Zoologica, 1930. gister of Marine Species. http://www.
Archo. Zool. Ital., 16(2), 601-617. marinespecies.org
Willey, A. (1935). Harpacticoid Copepoda Zaleha, K., Nasiratul, M., Siang, H. & Kama-
from Bermuda. Part. II. Ann. Mag. ruzzaman, B. (2016). Trend of meio-
Nat. Hist., 15, 50-100. https://doi. benthos density and composition in
org/10.1080/00222933508654944 Karah Island, South China Sea. Sains
Woods, D. & Tietjen, J. (1985). Horizon- Malaysiana, 45(7), 1019-1024.
tal and vertical distribution of meio-
fauna in the Venezuela Basin. Mar.
Geol., 68, 233-241. https://doi.
org/10.1016/0025-3227(85)90014-3
Wołowicz, M., Sokołowski, A., Urban-Ma-
linga, B. & Szymelfenig, M. (2011).
Meiofauna as consumers in coas-
tal food webs. In E. Wolanski & D.
McLusky (Eds). Treatise on estuarine
and coastal science (pp. 173-202). Po-
land: Elsevier. https://doi.org/10.1016/
B978-0-12-374711-2.00611-2
Rev. Mar. Cost. Vol. 12 (2): 95-113, julio-diciembre 2020. 113
ISSN: 1659-455X • e-ISSN: 1659-407X
DOI: http://dx.doi.org/10.15359/revmar.12-2.5You can also read

















































