Reproductive success of threatened northern Lesser Black-backed Gulls (Larus fuscus fuscus) in relation to nest predation by Ravens (Corvus corax)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Ornis Fennica 99: 1–14. 2022
Reproductive success of threatened northern
Lesser Black-backed Gulls (Larus fuscus fuscus)
in relation to nest predation by Ravens (Corvus corax)
Jan Ove Bustnes*, Morten Helberg & Bård-Jørgen Bårdsen
J. O. Bustnes, Norwegian Institute for Nature Research, High North Research Centre
on Climate and the Environment (FRAM), NO-9296 Tromsø, Norway * Corresponding
author's e-mail: Jan.Bustnes@nina.no
M. Helberg, BirdLife Norway, Sandgata 30 B, 7012 Trondheim
B.J. Bårdsen, Norwegian Institute for Nature Research, High North Research Centre on
Climate and the Environment (FRAM), NO-9296 Tromsø, Norway
Received 29 April 2021, accepted 17 February 2022
Many seabird populations suffer heavily from the destruction of nests by generalist
predators. In this study, we analyzed 16 years of data (2005–2020) on the reproductive
output of the northern Lesser Black-backed Gull (Larus fuscus fuscus) at Horsvær, the
largest assemblage of this subspecies in Norway (up to ~400 pairs), in relation to the
occurrence of breeding Ravens (Corvus corax). A pair of Ravens were firstly discovered
at Horsvær in 2010, and between 2011 and 2016 they were observed with broods (2–5
fledglings) in most years. Between 2017 and 2020, human intervention prevented the
Ravens from breeding in the colony. However, in 2020 a pair of Ravens brought their
fledglings over from a neighboring island in the middle of the incubation period for the
gulls. On average, the nest predation rate was 43% when Ravens had fledglings within
the study area. In contrast, only 10% of nests were depredated in years when Ravens
did not reproduce successfully or were absent. Moreover, only 0.07 fledglings were on
average produced per nest in years when Ravens bred successfully, compared to 0.71
fledglings per nest in years with no Raven reproduction. A high level of nest predation
led to a decline in the number of nesting gulls, which was not observed in a neighboring
Raven-free colony. Finally, in years with high Raven predation at Horsvær, production
of fledglings was still high in yet another nearby Lesser Black-backed Gull colony.
The Ravens were established at Horsvær in the absence of people in the spring, and the
only option to save these threatened gulls may be to prevent the Ravens from nesting
successfully in or near their colonies.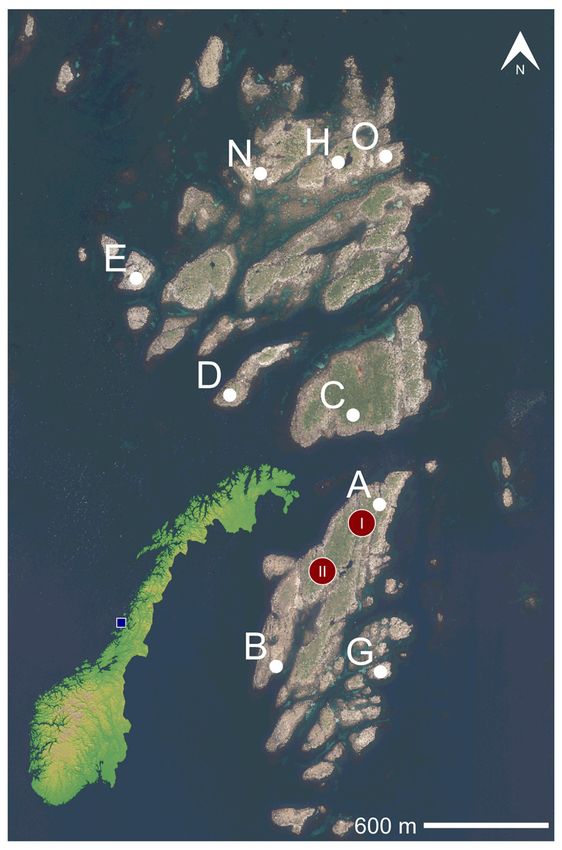
2 ORNIS FENNICA Vol.99, 2022
1. Introduction Bustnes et al. unpublished data).
The northern Lesser Black-backed Gull has
Predation on nests has long been identified as a been found to behave differently from other gull
primary source of reproductive loss in various species regarding feeding ecology and migration
bird species (Ricklefs 1969, Martin 1993). It is, (Strann & Vader 1992, Juvaste et al. 2017,
however, debated how important nest predation Helberg et al. 2009, Bustnes et al. 2013). In this
by generalist predators, such as corvids, is for the study, we noted that these gulls were less aggres-
productivity and abundance of birds (Madden et sive than the other gull species, such as Common
al. 2015). However, there are several examples Gull (L. canus), Herring Gull (L. argentatus) and
of Ravens (Corvus corax) having substantial Great Black-backed Gull (L. marinus). When
negative impacts on the nesting success of seabirds we were present, the Lesser Black-backed Gulls
(e.g. Maccarone 1992, Avery et al. 1995, Peery & mostly flew high and rarely feigned attacks at
Henry 2010, Carle et al. 2017, Ekanayake et al. us, or they only sat in flocks in the tidal zone.
2015). Seabirds such as gulls may be fierce nest
predators themselves, but some species are also
vulnerable to nest predation (Massaro et al. 2001,
Kazama 2007, Scopel & Diamond 2017, Mills et
al. 2018), including the Lesser Black-backed Gull
(Larus fuscus) (Calladine 1997, Bukacinski et al.
1998, Hario 1994, Hallgrimsson & Hersteinsson
2012).
A large proportion of the Lesser Black-backed
Gull population in northern Norway consists of
the nominate subspecies (L. f. fuscus) which is
threatened over its whole distribution range, espe-
cially in Finland and Norway (Hario et al. 1998,
Helberg et al. 2009, Juvaste et al. 2017). The
Norwegian population has declined strongly since
the early 1970s (Bustnes et al. 2010a), which has
mostly been attributed to the crash in the Atlantic
herring (Clupea harengus) stock in the late
1960s (Myrberget 1985, Røv 1986, Bevanger &
Thingstad 1990, Strann & Vader 1992), although
other species of fish may also be important in the
gulls’ diet (Bustnes et al. 2010b).
In 2005, a demographic study of the northern
Lesser Black-backed Gull was started at Horsvær,
a small archipelago in the southern part of
Nordland County (Fig. 1), which was the largest
assemblage of this subspecies in Norway (Bustnes
et al. 2010a). Anecdotal evidence suggests high Fig.1. Aerial photo of Horsvær, on the Norwegian Coast,
breeding numbers at Horsvær in the 1950s (S. showing the position of 9 different sub-colonies of Lesser
Black-backed Gulls (A–O) occupied between 2005 and
Jørgensen, pers. comm.). The first nest counts 2020. The A-, D-, and H-sub-colonies have been mostly
in 2005 and 2006 recorded close to 400 nests, abandoned after 2014, whereas N and O sub-colonies
distributed over seven sub-colonies (Bustnes et were largely established after 2014. In addition, the
al. 2020). Subsequent studies have shown that positions of two Raven nests are given in red (I = first
adult Lesser Black-backed Gulls frequently move nesting site; II = second nesting site). The aerial photo
was generated through three projects (Nordland Sør
between these sub-colonies, permanently or 2014, 2010 and 2009) and was downloaded from https://
visiting, but rarely move to other colonies once norgeibilder.no/ (a collaboration between the Norwegain
established at Horsvær (Bustnes et al. 2020, J.O. Mapping Aurthorties and Geovekst).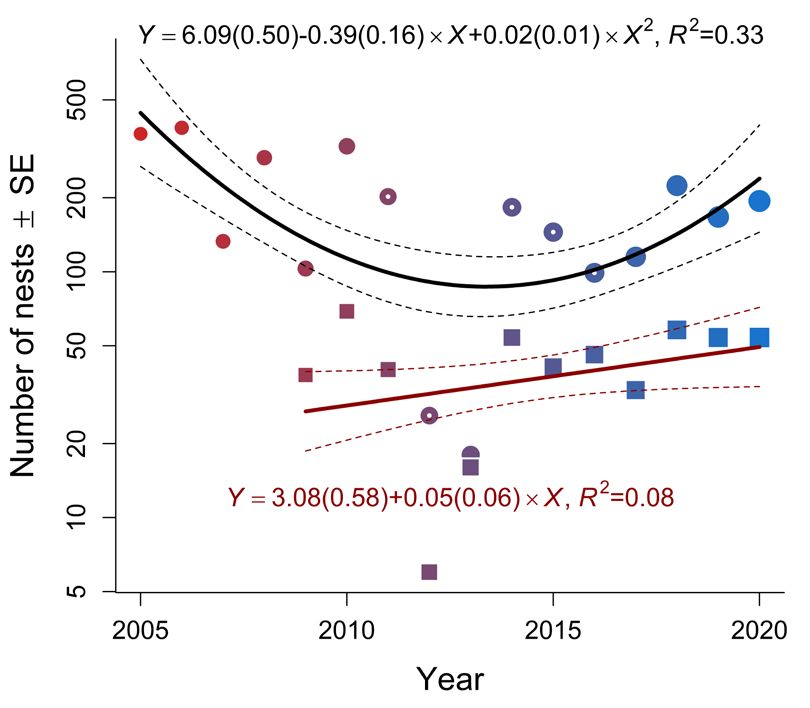
Bustnes et al.: Raven predation on Lesser Black-backed Gulls 3
They also appeared to show low aggression environmental conditions; 3) contrasted Horsvær
toward potential nest predators, such as corvids with the fledgling production in Fjordholmen,
and other gulls. This contrasts with another sub- a Raven-free colony 46 km from Horsvær (data
species of Lesser Black-backed Gulls (L. f. available for the years: 2015, 2017–2018 and
graellsi), which may be very aggressive in en- 2020). A key point in our study design was that
counters with herring gulls (Garthe et al. 1999). the distance between the colonies is far enough
The number of Lesser Black-backed Gulls to prevent Ravens from making routine trips, but
attempting to nest varied dramatically among short enough for environmental conditions to be
the years, as did their reproductive output. For similar.
example, in 2005 and 2006, more than 350 fledg-
lings were produced, whereas in 2012 and 2013,
less than 30 pairs nested, and no fledglings were 2. Material and methods
produced (Bustnes et al. 2020). We firstly attrib-
uted this to variation in feeding conditions, but 2.1. Study area
we also noted an increasing decoupling between
the number of birds attempting to nest and the The study location was Horsvær (65º19’N,
production of young over the years. Hence, eggs 11º37’E) an archipelago at Helgeland (Nordland
and chicks seemed to be lost at a higher rate. County) in northern Norway. Totally, nine
People inhabited Horsvær for hundreds of different sub-colonies (A–O) were included in
years, and subsistence exploitation of natural this study (Fig. 1), two being on the same island
resources, such as seabird eggs and Common (A and B) whereas all other sub-colonies were on
Eider (Somateria mollissima) down, was different islands. There were seven sub-colonies
important. People kept generalist predators such when the study started in 2005 (A−H; Fig. 1), but
as corvids at bay. However, after the permanent all nests in the G-colony were depredated in 2008
settlements were abandoned in the 1970s, and and the sub-colony was subsequently abandoned
people gradually ceased visiting the islands, (Bustnes et al. 2020). After 2014, the A, D and H
Ravens could successfully establish (i.e. get sub-colonies have more or less vanished (usually
through their vulnerable nesting phase). We firstly 0–2 nests per years), and the N- and O-colonies
noted a pair of Ravens in 2010, and between 2011 were established (Fig. 1). In addition there were
and 2016, they reproduced successfully at or in a varying number of gulls nesting outside of the
close vicinity of Horsvær in most years. In 2017, established sub-colonies, which has been included
we were permitted to remove the Ravens, and in the total number of nests in Table 1. Four of the
they were consequently prevented from breeding sub-colonies (B, E, G, H and N) were dominated
at Horsvær in the subsequent years. by open-rocky habitats and had no vegetation
This study aimed to analyze the relationship taller than a few cm, and nearly all nests were
between the reproductive success of northern openly exposed (> 90%). The other colonies (A,
Lesser Black-backed Gulls and the occurrence C, D and O) were dominated by dense vegetation
of breeding Ravens. We applied a natural exper- (~50−80 cm tall) consisting of meadowsweet
imental design where we: 1) contrasted the depre- (Filipendula ulmaria): in the A- and D-colony
dation of nests and fledgling production in years more than 70% of the nests were in the vegetation,
with and without breeding Ravens; 2) contrasted and more than 90% of the nests were located in
the number of nests at Horsvær with Svindraget, the vegetation in the C-colony.
a Raven-free neighboring colony located 8 km Since 2009, we have monitored the number of
apart. This allowed us to estimate the temporal nests at Svindraget (65º15’10’’N, 11º41’31’’E),
trends in the number of nests both at Horsvær and a Lesser Black-backed Gull colony on a flat
Svindraget, and the extent to which the temporal open rocky island located ~7.8 km southeast of
dynamics in the number of nests differs across Horsvær, which is presently not inhabited by
these two areas. The contrasts between Svindraget people. In addition, in the years 2015, 2017–2018
and Horsvær has the potential to estimate the and 2020, we visited Fjordholmen (65°37'27"N,
effect of Raven predation in the context of overall 12°18'13"E) in Vevelstad Municipality (~46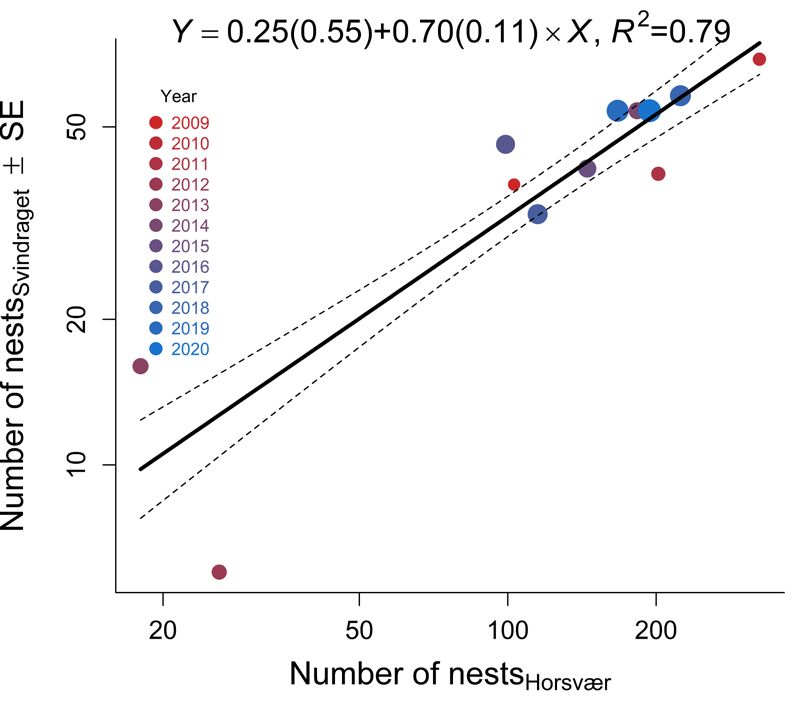
4 ORNIS FENNICA Vol.99, 2022
Table 1. Reproductive variables of Lesser Black-backed Gulls from Horsvær in relation to occurrence of breeding
Ravens, and the number of nests at a nearby colony, Svindraget, where Ravens were absent.
No. of Nests Percent Fledglings Breeding Nests at
Year Fledglings
nests depredated depredated nest -1 Ravens Svindraget
2005 364 5 372 1.37 1.02 Absent –
2006 385 1 349 0.3 0.91 Absent –
2007 133 17 100 12.8 0.75 Absent –
2008 291 28 130 9.6 0.45 Absent –
2009 103 28 33 27.2 0.32 Absent 38
2010 324 25 312 7.7 0.96 Absent 69
2011 202 58 6 28.7 0.03 Present 40
2012 26 23 0 88.5 0 Present 6
2013 18 14 0 77.8 0 – 16
2014 183 24 16 13.1 0.09 Present 54
2015 145 55 31 37.9 0.21 Present 41
2016 99 50 0 50.5 0 Present 46
2017 114 23 55 20.2 0.48 Absent 33
2018 224 19 158 8.5 0.71 Absent 58
2019 166 12 142 7.2 0.86 Absent 54
2020 193 7 20 3.6 0.10 Present * 54
*Arrived after we left Horsvær
km northeast from Horsvær), after banding depredated when found (i.e. destroyed eggs in or
fledglings at Horsvær. The Lesser Black-backed near the nest), and the rate of nest predation was
Gull colony at Fjordholmen consists of three assessed as the percentage of nests depredated
sub-colonies, within a limited vegetated area, (either when found or between nest checks)
with 100–150 pairs in total. Importantly, people of the total number of nests. During both nest
still inhabit Fjordholmen in the gulls’ breeding checks (searches), we recorded clutch sizes.
season. Egg laying starts in early June, and is primarily
finalized in the latter half of June when we
were present. We are thus confident that our
2.2. Study protocol estimate of the number of nests in the colony is
reasonably accurate. However, we sometimes
In 2005, the fieldwork lasted from mid-June to found a few nests with eggs during our second
late July. In the subsequent years, the fieldwork visit, which we recorded. Each year during our
was divided in two field trips: 1) a period of second field trip, all islands were visited once
6–10 days in mid-June; and 2) 1–3 days in late and carefully searched for chicks (fledglings).
July early August. This set-up was chosen to The chicks were laid on the ground (in the
reduce the human disturbance of this threatened grass or close to rocks) to calm them while still
subspecies. During the first period, all colonies handling other chicks. Chicks that we were
were searched, and all nests were recorded and unable to catch, e.g. escaped to the sea, were
marked with a numbered wooden stick, and eggs counted. The exception was 2005, when banding
were marked with a waterproof pen. After 4−6 of chicks was conducted over a two week period.
days (3−7 days), we revisited all the nests again, This year we assumed that nearly all chicks were
and new nests were recorded. Some nests were found and banded, and no counts of unmarked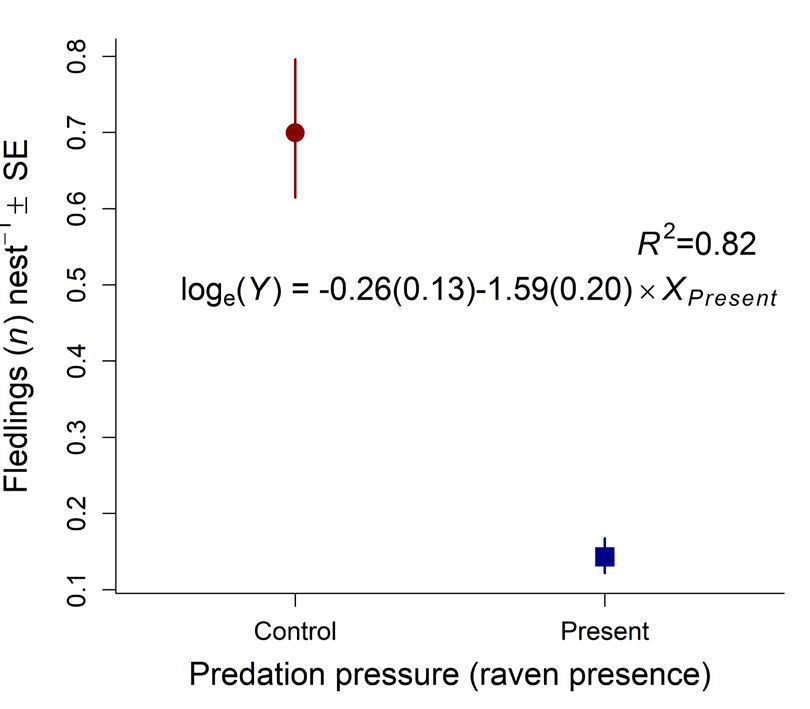
Bustnes et al.: Raven predation on Lesser Black-backed Gulls 5
Fig. 2. Photos showing the Raven nest on a scaffold on a building at Horsvær, and egg remains found under the
Raven nest and a roosting site: Lesser Black-backed Gull, Common Eider and Greylag Goose.
chicks were conducted. Fledgling production was observed on a small neighboring island
was assessed as the number of juveniles banded, when we arrived 8th of June. On the 14th of June,
in addition to the number of unmarked juveniles when we left, the Ravens were observed to bring
counted, in relation to the number of nests. This their brood of four fledglings from this neighbor-
also includes nests found when banding chicks ing island over to Horsvær. A license to kill the
since we assumed it was implausible that nests Ravens was granted by the Sømna Municipality
hatching later than 25th of July would success- in 2017, but was never effectuated.
fully produce fledglings. In the middle of our
first stay, we went to Svindraget and made a
single nest count, covering the whole island. At 2.4. Statistical analyses
Fjordholmen, due to vegetation and limited time
in the colony, an exact number of fledglings is We performed all statistical analyses and plotting
challenging to achieve, but acceptable estimates in R (R Core Team 2021). Our tests were two-
of production status can be achieved. Since 2017, tailed (rejecting the null hypothesis at an α-level
the Horsvær archipelago has also been visited in of 0.05), and Wald statistics were used to test the
April to record the presence of Ravens. hypothesis that the estimates were not signifi-
cantly different from zero. We used the treatment
contrast whenever predator pressure was included
2.3. Occurrence and behaviour of Ravens in a model: a two-level factor comparing years
with the presence of Ravens (Present: treatment) to
We recorded the first pair of Ravens in 2010. years when breeding Ravens was absent (Control)
In 2011 and 2012 they were observed with two from the Horsvær Archipelago. Testing our
fledglings each year, but they may have had more biological hypotheses required several different
fledglings. In 2014, a Raven nest was discovered statistical methods. The proportions of nests being
on a scaffold on an abandoned building amidst predated (the number of nests predated divided by
the Horsvær archipelago (Fig. 1 and Fig. 2). In the number of nests; see Table 1) were analyzed
2015, we removed the scaffold, and in 2016 the using beta regressions, while linear models (LMs)
Ravens moved to another abandoned building and Linear Mixed Effects (LME) models were
500 m further south of the same island (Fig. 1). used on responses that either were normally
Between 2014 and 2016, the Ravens produced distributed (clutch size; the number of eggs nest-1)
five fledglings annually. In 2017 the Raven nest, or when loge-transformation made them fulfill
including the eggs, was removed in April, and no the normality assumption [number of fledglings
relaying occurred. After 2017, no nest was found nest-1, nest counts (see Table 1), and date (days
at Horsvær, although a non-breeding pair of since June 1st); see below for details].
Ravens was observed in 2018 and 2019. In 2020,
however, a brood of Ravens (six birds in total)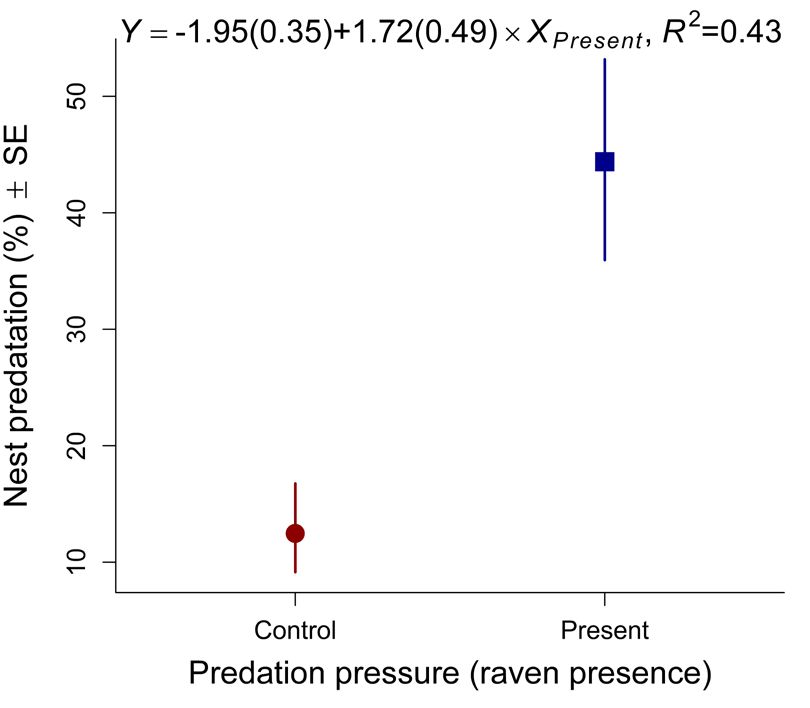
6 ORNIS FENNICA Vol.99, 2022
2.4.1. Nest predation and reproductive success First, in the log-log model, both the number
of nests at the Raven-free colony (Svindraget;
We used beta regression (Cribari-Neto & Zeileis response) and at our control area (predictor) were
2010) and the betareg-package (Zeileis et al. loge-transformed. We did this for two reasons.
2020) in the analyses of the proportion of nests First, we realized that the diagnostics for the model
being predated as a function of the predation without loge-transformation was poor (results not
pressure. In this analysis, we used a logit- and shown). Second and more importantly, the log-log
a log-link for the mean- and precision-model, models have the desired property that the slope
respectively (Cribari-Neto & Zeileis 2010). There for the predictor is approximately interpreted at a
were two reasons why we chose this modelling percentage increase in the response relative to one
approach over standard LMs: 1) the residuals percent change of the predictor (Gelman & Hill
plots revealed potential violations of the assump- 2007). Second, we ran two different linear models
tions behind LMs (e.g. Zuur et al. 2010); 2) the assessing the temporal dynamics of the number of
response represent proportions (defined within nests – in analyses of the data from each colony
an interval between zero and one). The average separately: 1) a simple linear regression model
of the beta-distribution is estimated by the α- and where we predicted the number of nests based on
β-parameters [E(Y) = μ = α / (α + β)], whereas the year (setting the intercept to 2005; i.e., 2005 = 0);
variance is modelled by estimating the precision 2) a second-order polynomial where we added
parameter [ɸ = α + β, where the variance is year2 to the first model. We did model selection
VAR(Y) = μ(1 − μ)/(1 + ɸ): Cribari-Neto & Zeileis similar to in the analyses of nest predation above.
2010]. We fitted two different models: 1) one with
the same structure for the precision and the mean
and precision sub-model (both were a function 2.4.3. Confounding effects
of predation pressure); 2) another one where the
precision sub-model only included the constant There were two crucial confounding factors
(i.e., assumed similar across the two levels of related to our study design that we wanted to
predation pressure). We selected the candidate test for. First, it was important to assess if clutch
model with the lowest second-order Akaike’s size at laying was confounded by the presence of
Information Criterion (AICc where Δi represents Ravens. For example, if clutch size was small in
the difference in AICc value between model i the years when Ravens were present for reasons
and the model with the lowest AICc value: (e.g., other than predation that could seriously affect our
Burnham & Anderson 2002, Anderson 2008) results. In line with B.J. Bårdsen et al. (unpub-
value using the AICcmodavg-package (Mazerolle lished) who predicted clutch size at laying (June
2020). In the analyses of fledgling success, we 1st; from a model where annual clutch size was
fitted standard LMs, using the lm-function in predicted based on date and year of recording)
R, but with a transformed response to fulfil the and used it as a predator-free measure of repro-
normality assumption: loge(fledglings nest-1 + duction, we used clutch size at our first visit as
0.1). Like in the analysis above, we used predation our closest empirical measure of laying clutch
pressure as the only predictor. size. We fitted several LME models (Pinheiro &
Bates 2000) – all with predator pressure as the
only fixed effect, but with three different random
2.4.2. Spatial synchrony effects (random intercepts only): 1) Year; 2)
Colony and 3) Colony nested within Year. These
To assess the spatial synchrony in the number models were fitted to nest-level annual data using
of nests at Horsvær (our control area; subject the lme-function, in the nlme-package (Pinheiro et
to Raven predation – at least some years) with al. 2020), with a Restricted Maximum Likelihood
the number of nests to a neighboring colony (i.e., setting the method-argument to “REML”)
(Svindraget) where Ravens were absent, we used: as the fixed effects were constant (Pinheiro &
1) log-log models; 2) LMs assessing temporal Bates 2000). We selected one model and used it
dynamics (both fitted using the lm-function). for inference, adopting the same model selectedBustnes et al.: Raven predation on Lesser Black-backed Gulls 7
procedure as in the other analyses. Second, we was statistically significant in the beta-regression
measured predation by visiting the nests twice, model (R2 = 0.36, Table 2, Fig. 3). The alterna-
it was important for us to assess if the number of tive model, including a similar structure for the
days in-between the visits were confounded by precision model as the mean model, had poor
the presence of breeding Ravens. We used LME support in our data (Table 2).
models and the same set-up as in the analyses of In the analysis of the production of fledglings,
clutch size above in this analysis. we included 2020 as a year with the presence
of breeding Ravens. The estimated number of
gull fledglings produced varied from 0 to 372,
3. Results whereas the mean number of fledglings per nest
varied from 0 to 1.02 (Table 1). In the six years
3.1. Nest predation and fledgling success when breeding Ravens were present, on average
0.07 (SE = 0.033, range = 0–0.23) fledglings
The number of Lesser Black-backed Gull nests were produced per nest, which was significantly
found at Horsvær varied from 18 (2013) to 385 lower than the average of 0.71 (SE = 0.083, range
(2006), on average 185.8 (SE = 27.5). Of these, = 0.33–1.02) produced in the nine years when
between 0.26% (2006) and 88.5% (2012) were breeding Ravens were absent (Table 3a, Fig 3).
depredated (Table 1). In the subsequent analysis
of predation pressure, we removed 2013 because
of extremely poor feeding conditions and that we 3.2. Raven behaviour and predation
did not know the status of the Ravens in that year,
as we did not observe them. Raven parents and their brood were observed to
Defining 2020 as a year where Ravens were operate as a unit, attacking sub-colonies system-
absent (as they moved into the study area the atically and clearing large proportions of the nests
day we left and hence did not affect the number within a few days. In 2011, 53% of 73 nests in the
of nests observed), 43.1% (SE = 12.8, range = C-colony (Fig. 1) were lost before the second nest
13.1−88.5%) of the nests on average were depre- check, compared to only 10% of 50 nests in the
dated in the five years when Ravens reproduced. H-colony. However, in 2011 only two fledglings
In comparison, the nest predation rate was only were produced in the C- and none in the H-colony.
9.6% (SE = 2.5, range = 0.26–26.2%) in the ten Similarly, in 2015, 84% of 31 nests were de-
years when Ravens did not reproduce (Table 1). predated in the E-colony before the second nest
This 4-fold increase in the average nest predation check, compared to only 20% out of 15 nests in the
rate in years with breeding Ravens present N-colony. However, no fledglings were produced
compared to control years (with only baseline in N- and the E-colony in 2015. In 2016, all out of
predation levels; i.e. predators other than Ravens) 21 nests were depredated in the C-colony before the
Table 2. Estimates from the beta-regression model relating the proportions of nests at Horsvær (2005–2019)
being predated as a function of predation pressure using the logit- and log-link for the mean- and precision-model,
respectively. We used the treatment contrast, estimating the difference between years when Ravens were present
(treatment) and control-years when Ravens were absent (Intercept; see main text for details regarding how predation
pressure was defined). R 2 is the pseudo-R 2 reported in the output from models fitted in betareg-package. The
difference in AICc-values (Δ i ) between this an alternative models where the precision was modelled in the same
manner as the average structure.
Parameter Estimate SE z P-value
Intercept –1.883 0.329 –5.7268 ORNIS FENNICA Vol.99, 2022 second nest check whereas the E-, N-, O-colonies at Horsvær in 2016. Hence, the Ravens continued (13, 21 and 29 nests, respectively) lost ~38% of the to clear the remaining nests in these years after we nests, each. However, no fledglings were produced left the area in the second half of June. A A B B Fig. 3. (A) Predicted proportions (re-calculated into Fig. 4. (A) The relationship between number of Lesser percentages) of nest predated as a function of Predation Black-backed Gull nests at Horsvær and Svindraget pressure (Control: years where breeding Ravens (both axis at loge-scale), where the distance from were absent; Present: years where breeding Ravens Horsvær’s center colony (C-colony, Fig. 1) and was present at the archipelago; Table 1) in the beta- Svindraget is 7.8 km, in the period between 2009 and regression model reported in Table 2. (B) Similarly, 2020 (see Table 1 for the underlying data). (B) The predicted fledglings nest-1, back-transformed from temporal dynamics for the same response in each loge(fledglings nest-1+0.1) to normal-scale, as a function area (Horsvær and Svindraget in circles and squares, of predation pressure in a linear model (i.e., a one-way respectively). Svindraget showed no evidence of Analysis of Variance; see Table 3 for details). In both any temporal dynamics whereas Horsvær showed figures, text shows the coefficient of determination (R2; evidence of a curved relationship – being at its lowest please note that this was estimated differently across the in the 2012 (i.e. after Ravens started to breed in the models) and the estimated parameters (including theirs area; see Table 3b–d for the underlying statistical precision in parentheses): the difference between when analyses). Color figure is available in the online Ravens were present (Xpresent) and the control situation version of this article. when Ravens were absent (Intercept).
Bustnes et al.: Raven predation on Lesser Black-backed Gulls 9
Table 3. Estimates from linear models relating: (a) loge(number of fledglings nest-1+0.10) to Predation pressure (Table
2 provided details regarding the predictor variable and how to interpret its effect; Table 1 provides the underlying
data); (b) the temporal dynamics of the number of nests at Horsvær (where Ravens were present some years); (c) the
temporal dynamics of the number of nests at Svindraget (where Ravens have not been observed) and (d) Hosrvær
(where breeding ravens have been observed some years). R 2 is the coefficient of determination, and the difference in
AICc-values (Δ i ; b–c) represents the difference between the selected model presented here and the alternative model
(see main text for details).
Parameter Estimate SE t P-value
(a) Fledglings nest-1 + 0.10 (loge-scale)
Intercept –2.031 0.089 –22.95010 ORNIS FENNICA Vol.99, 2022
Table 4. Estimates from linear mixed effect models relating: (a) clutch size and (b) loge(number of days in-between
visits) to Predation pressure (a two-level factor variable consisting of a Control, or baseline, in which Ravens were
absent compared to when Ravens were present; Table 1 provides the underlying data). The difference in AICc-values
(Δ i ) between the selected models, which constates of a random effect including both Colony and Year, and the two
other candidate models were as follows (for the analyses of Clutch size and days between visits, respectively): 1)
41.451 and 667.659 (Year); and 2) 173.431 and 2482.737 (Colony).
Parameter Estimate SE df t P-value
(a) Clutch size
Fixed effects
Intercept 2.351 0.079 2237 29.935Bustnes et al.: Raven predation on Lesser Black-backed Gulls 11
roosting sites, in addition to eggs of Lesser Black- years, which seems mainly to be a result of
backed Gulls (Fig. 2). variation in the feeding conditions. This is
The relatively dense nesting colonies and low supported by the high correlation between the
aggression of the gulls, combined with a cooper- number of nests at Horsvær and Svindraget. For
ative hunting tactic, made Ravens able to rapidly example, in the poor breeding seasons of 2009
clear the sub-colonies. Hence, if unmanaged, a and 2013, we noted that the gulls were feeding
single pair of Ravens could effectively curtail gull heavily on Blue Mussels (Mytilus edulis), an
reproduction over many years. Moreover, gulls may indication of low fish availability and thus poor
abandon their colonies completely if all their nests feeding condition. Hence, high predation on nests
are destroyed synchronously (Coulson & Coulson could also be a multiple stressor effect: i.e., if
2009), which happened to the G-colony (Fig. 1) in predation increased due to poor feeding condi-
2008. Although the culprit was not identified, we tions, the low prospect of success may have made
suspect that it was Ravens with a brood coming birds less motivated for defending their eggs, and
over from another island (Bustnes et al. 2020). thus increasing the likelihood that predators gain
Furthermore, the abandonment of the three other access to the nests. However, in some years with
sub-colonies (A, D and H, Fig. 1) happened after Ravens (2011, 2014 and 2020), the number of
the Ravens had ravaged most of the reproduction nesting attempts were high, and it is noteworthy
for several years. It is also noteworthy that a large that the birds seemed to be highly motivated in
proportion of the colonies of northern Lesser Black- 2020. In all years since 2010, it was difficult to
backed Gulls in the region of southern Nordland catch birds with nest cages, but in 2020 they went
disappeared between 1980 and 2007 (Bustnes & straight into the cages, as soon as we withdrew
Helberg 2010c). This happened, although there is from the sites. However, despite high reproduc-
little evidence of generally poor feeding conditions tive investment, the gulls appeared to be unable to
in this period (Bustnes et al. 2010a). avoid nest predation by the Ravens that arrived in
People have inhabited the Norwegian Coast the middle of their incubation period. Moreover,
for millennia and influenced the wildlife popula- our observations from Fjordholmen clearly shows
tions, but in the post-war era depopulation gained that a Raven-free colony may have a stable and
traction, accelerating over the last 60 years. In high reproductive output independent of Raven
northern Norway, subsistence exploitation of activities at Horsvær. There might be several
seabirds, such as gulls, alcids and eiders was differences between Fjordholmen and Horsvær,
important, and to maximize production, people such as breeding habitats. However, there is little
persecuted predators such as corvids and eagles. evidence that this factor is essential for production
For White-tailed Eagles (Haliaeetus albicilla), in these Lesser Black-backed Gulls (Bustnes et al.
conservation efforts have led to population 2020). The most important difference is probably
growth, which has caused increased predation on that people still inhabit Fjordholmen.
nesting seabirds, also in Norway (Hipfner et al. Open nesting seabird species, such as gulls,
2012, Hentati-Sundberg et al. 2020). Populations terns and guillemots, seem to be more vulnerable
of many corvids have increased worldwide in to nest predation by generalist predators than
response to human changes to the environment species with concealed nesting (McMahon et al.
(Marzluff et al. 2006), but the population devel- 2020, Hentati-Sundberg et al. 2020). However,
opment of Ravens in Norway is poorly known. at Horsvær, both Herring Gulls and Great
However, less persecution of Ravens has probably Black-backed Gulls seemed far less vulnerable
led to birds becoming bolder and increased their to Ravens than Lesser Black-backed Gulls, and
numbers in seabird colonies. Hence, when people we did not observe Ravens in or near the nests of
were leaving their coastal settlements, the Ravens these aggressive gulls. The Common Gull is also a
had better prospects of success, as seen at Horsvær potentially aggressive species, and in 2016
where Ravens only established after people ceased when no Lesser Black-backed Gull chicks were
coming regularly there in spring. produced, we still found surviving Common Gull
The number of Lesser Black-backed Gulls fledglings. However, the number of Common
attempting to nest varied dramatically between Gulls pairs have also declined at Horsvær over12 ORNIS FENNICA Vol.99, 2022
the last ten years (J.O. Bustnes & M. Helberg, fuscus) häckningsframgång vid Horsvær, den
unpublished data), so heavy predation on this största samlingen av denna underart i Norge (upp
species cannot be excluded. A central question emot 400 par), i förhållande till förekomsten av
is whether these gull species and other potential häckande korpar (Corvus corax). Ett korppar
nest predators, such as White-tailed Eagles, återfanns för första gången på Horsvær 2010 och
Hooded Crows (Corvus corone cornix), Arctic mellan 2011 och 2016 producerade de kullar med
Skuas (Stercorarius parasiticus), American Mink 2–5 flygga ungar under de flesta åren. Mellan 2017
(Mustela vison) and Common Otters (Lutra lutra), och 2020 gjordes ingrepp som hindrade korparna
could be responsible for some the heavy predation från att häcka i kolonin. Under 2020 hämtade
on the Lesser Black-backed Gulls. All these ett korppar sina flygga ungar till Horsvær från
species are present, except American Mink, and en närliggande ö, mitt under ruvningsperioden
likely to prey on nests of the Lesser Black-backed för trutarna. Bopredationsraten var 43% när
Gulls. However, they have been there during the korparna hade sina flygga ungar i området. Till
whole study period, and the heavy predation only skillnad från detta skedde bopredation endast i
occurred when Ravens were producing fledglings. 10% av trutboen när korparna inte häckade eller
This study suggests that an intermediate-sized, var borta. Dessutom producerades i medeltal
colonial nesting gull such as the non-aggressive endast 0.07 flygga trutungar per bo när korparna
northern Lesser Black-backed Gull does not hade ungar, medan det producerades i medeltal
have a working defensive strategy when Ravens 0.71 ungar per bo när korparna inte häckade. En
establish close to their colonies, even when they hög nivå av bopredation ledde till en minskning
appear to be highly motivated for breeding. i antalet häckande trutar, medan en minskning
Thus, even the largest Norwegian colony of this inte observerades i en närliggande korpfri koloni.
subspecies seems to have a gloomy future if Slutligen, under år med hög korppredation på
no protective measures are taken. As such, our Horsvær var häckningsframgången fortfarande
analyses do suggest declining numbers of nesting hög i en närliggande koloni. Korparna etablerade
gulls since the study started, a trend different sig på Horsvær när inga människor befann sig
from the nearby Svindraget. Human beings are på ön, och det enda alternativet att skydda dessa
an intrinsic part of most ecosystems (Mace 2014), hotade trutar är genom att förhindra att korpar kan
and the Norwegian Coast has been inhabited for häcka nära eller i trutkolonierna.
thousands of years. People have influenced the
ecosystem by their exploitation of seabirds and Acknowledgements. We are grateful to several people
their control of predators. This has probably laid assisting us during fieldwork, notably Trond Johnsen,
Nils H. Lorentsen, Klaus Torland, Geir A. Bustnes and
the groundwork for large populations of some
Jorg Welcker. We are grateful to Harald Bustnes, Sigfred
species, including Lesser Black-backed Gulls Jørgensen and Runar Omnø for logistical support during
in some areas. We believe that the only way to our stay at Horsvær. We also thank two anonymous
protect these northern Lesser Black-backed Gulls reviewers that gave valuable comments to the manuscript.
is by preventing Ravens from reproducing in or The study was funded by Norwegian Seabird Program
close to their colonies. We propose that people (SEAPOP).
either locate the Raven nests and impairing their
reproduction or visit the gull colonies before and
at the onset of egg-laying of the Ravens. References
Anderson, D.R. 2008: Model based inference in the life
Förökningsframgång hos den hotade silltruten sciences: a primer on evidence. Springer Science, New
York, United States of America.
(Larus fuscus fuscus) i förhållande till
Avery, M.L., Pavelka, M.A., Bergman, D.L., Decker, D.G.,
bopredation av korpar (Corvus corax) Knittle, C.E. & Linz, G.M. 1995: Aversive conditioning
to reduce Raven predation on California least tern eggs.
Många sjöfågelpopulationer lider kraftigt av — Colonial Waterbirds 18: 131–138.
bopredation. I denna studie analyserade vi 16 års Bevanger, K. & Thingstad, P-G. 1990: Decrease in some
data (2005–2020) om silltrutens (Larus fuscus Central Norwegian populations of the northernBustnes et al.: Raven predation on Lesser Black-backed Gulls 13
subspecies of the Lesser Black-backed Gull (Larus Hallgrimsson, G.T. & Hersteinsson, P. 2012: Spatial
fuscus fuscus) and its possible causes. — Fauna contraction in a large gull colony in relation to the
Norvegica Ser C, Cinclus 13: 19–32. position of arctic fox dens. — European Journal of
Burnham, K. P., & Anderson, D. R. 2002: Model selection Wildlife Research 58: 441−450.
and multimodel inference: a practical information- Hario, M. 1994: Reproductive-performance of the nominate
theoretic approach. Second edition. Springer, Inc., New Lesser Black-backed Gulls under the pressure of
York, USA. herring gull predation. — Ornis Fennica 71: 1–10.
Bukacinski, D., Bukacinska, M. & Spaans, A.L. 1998: Hario, M., Bianki, V. & Zimin, V. 1998: Larus fuscus fuscus.
Experimental evidence for the relationship between — In: Kotiranta, H., Uotila, P., Sulkava, S. & Peltonen
food supply, parental effort and chick survival in the S.-L. (eds.). Red Data Book of East Fennoscandia.
Lesser Black-backed Gull. Larus fuscus. — Ibis 140: Ministry of Environment, Finnish Environment Institute
422−430. and Botanical Museum, Finnish Museum of Natural
Bustnes, J.O., Anker-Nilssen, T. & Lorentsen, S-H. 2010a: History, Helsinki, pp. 247–249.
Local and large-scale climatic variables as predictors of Helberg, M., Systad, G. H., Birkeland, I., Lorentzen, N. H. &
the breeding numbers of endangered Lesser Black- Bustnes, J. O. 2009: Migration patterns of adult and
backed Gulls on the Norwegian Coast. — Journal of juvenile lesser black-backed gulls breeding in northern
Ornithology 151: 19–26. Norway. Ardea 97: 281–286.
Bustnes, J.O., Barrett, R.T. & Helberg, M. 2010b: Northern Hentati-Sundberg, J., Berglund, P-A., Hejdström, A. &
Lesser Black-backed Gulls; what do they eat? — Olsson O. 2020: COVID-19 lockdown reveals tourists
Waterbirds 33: 534−540. as seabird guardians. — Biological Conservation 254:
Bustnes, J.O. & Helberg, M. 2010c: What happened to the 108950.
Lesser Black-backed Gulls in northern Norway. — Hipfner, M.J., Blight, L.K., Lowe, R.W., Wilhelm, S.J.,
Ottar 5: 36−42 (in Norwegian). Robertson, G.J., Barrett, R.T., Anker-Nilssen, T. &
Bustnes, J.O., Moe, B., Helberg, M. & Phillips, R. 2013: Good, P. 2012: Unintended consequences: how recovery
Rapid long-distance migration in Norwegian Lesser of sea eagles Haliaeetus spp. Populations in the northern
Black-backed Gulls Larus fuscus fuscus along their hemisphere is affecting seabirds. — Marine Ornithology
eastern flyway. — Ibis 155: 402−406. 40: 39−52.
Bustnes, J.O., Bårdsen, B.J. & Helberg M. 2020: To hide or Juvaste, R., Arriero, E., Gagliardo, A., Holland, R., Huttunen,
not to hide: nesting habitat dynamics in a threatened M.J., Mueller, I., Thorup, K., Wikelski, M., Hannila, J.,
gull. — Waterbirds 43: 163–173. Penttinen, M.L. & Wistbacka, R. 2017: Satellite tracking
Calladine, J. 1997: A comparison of Herring Gull Larus of red-listed nominate Lesser Black-backed Gulls
argentatus and Lesser Black-backed Gull Larus fuscus (Larus f. fuscus): Habitat specialization in foraging
nest sites: their characteristics and relationships with movements raises novel conservation needs. — Global
breeding success. — Bird Study 44: 318–326. Ecology and Conservation 10: 220–230.
Carle, R.D., Calleri, D.M., Beck, J.N., Halbert, P. & Hester, Kazama, K. 2007: Factors affecting egg predation in Black-
M.M. 2017: Egg depredation by common Ravens tailed Gulls. — Ecological Research 22: 613−618.
Corvus corax negatively affects pelagic cormorant Maccarone, A.D. 1992: Predation by common Ravens on
Phalacrocorax pelagicus reproduction in central cliff-nesting black-legged kittiwakes on Baccalieu
California. — Marine Ornithology 45: 149–157. Island, Newfoundland. — Colonial Waterbirds 15: 253–
Coulson, J.C. & Coulson, B.A. 2009: Ecology and colonial 256.
structure of large gulls in an urban colony investigations Mace, G.M. 2014: Whose conservation? — Science 345:
and management at Dumfries, SW Scotland. — 1558–1560
Waterbirds 32: 1–15. Madden, C.F., Arroyo, B. & Amar, A. 2015: A review of the
Cribari-Neto, F., & Zeileis, A. 2010: Beta Regression in R. impact of corvids on bird productivity and abundance.
Journal of Statistical Software 34:1–24. https://www. — Ibis 157: 1–16.
jstatsoft.org/v034/i02. Martin, T.E. 1993: Nest predation and nest sites. —
Ekanayake, K.B., Sutherland, D.R., Dann, P. & Weston, BioScience 43: 523−532.
M.A. 2015: Out of sight but not out of mind: corvids Marzluff, J.M. &Neatherlin, E. 2006: Corvid response to
prey extensively on eggs of burrow-nesting penguins. human settlements and campgrounds: causes,
— Wildlife Research 42: 509–517. consequences, and challenges for conservation. —
Garthe, S., Freyer, T., Huppop, O. & Wolke, D. 1999: Biological Conservation 130: 301− 314.
Breeding Lesser Black-backed Gull Larus graellsii and Mazerolle, M. J. 2020: AICcmodavg: Model selection and
Herring Gulls Larus argentatus: Coexistence or multimodel inference based on (Q)AIC(c). R package
competition? — Ardea 87: 227–236. version 2.3–1.
Gelman, A. & Hill, J. 2007: Data analysis using regression Massaro, M., Chardine, J.W. & Jones, I.L. 2001:
and multilevel/hierarchical models. Cambridge Relationships between Black-legged Kittiwake nest site
University Press, Cambridge, United Kingdom. characteristics and susceptibility to predation by large14 ORNIS FENNICA Vol.99, 2022
gulls. — Condor 103: 793−801. effects model. R package version 3.1–152.
McMahon, B.J., Doyle, S., Gray, A., Kelly, S.B.A. & R Core Team. 2021: R: a language and environment for
Redpath, S.M. 2020: European bird declines: do we statistical computing. R Foundation for Statistical
need to rethink approaches to the management of Computing, Vienna, Austria.
abundant generalist predators? — Journal of Applied Ricklefs, R. E. 1969: An analysis of nesting mortality in
Ecology 57: 1885–1890. birds. — Smithsonian Contribution in Zoology 9: 1−48.
Mills, J.A., Yarrall, W., Bradford-Grieve, J.M., Morrissey, Røv, N. 1986: Bestandsforhold hos sildemåke Larus fuscus i
M. & Mills, D.A. 2018: Major changes in the red-billed Norge med hovedvekt på L. f. fuscus. — Vår Fuglefauna
gull (Larus novaehollandiae scopulinus) population at 9: 79–84. (In Norwegian with English summary)
Kaikoura Peninsula, New Zealand; causes and Scopel, L.C & Diamond A.W. 2017: The case of lethal
consequences: a review. — Notornis 65: 14–26. control of gulls on seabird colonies. — Journal of
Myrberget, S. 1985: Breeding of Lesser Black-backed Gulls Wildlife Management 81: 572−580.
at Tranøy in Senja [Hekking av sildemåke på Tranøya i Strann, K-B. & Vader, W. 1992: The nominate Lesser Black-
Senja]. — Fauna 38: 140–145. (In Norwegian, with backed Gull Larus fuscus fuscus, a gull with a tern-like
English summary). feeding biology, and its recent decrease in northern
Peery, M.Z. & Henry, R.W. 2010: Recovering marbled Norway. — Ardea 80: 133–142.
murrelets via corvid management: A population viability Zeileis, A., Cribari-Neto, F., Gruen, B., Kosmidis, I., Simas,
analysis approach. — Biological Conservation 143: A.B., & Rocha, A.V. 2020: betareg: Beta Regression. R
2414–2424. package version 3.1–3.
Pinheiro, J. C. & Bates, D.M. 2000: Mixed effect models in Zuur, A.F., Ieno, E.N. & C.S. Elphick. 2010: A protocol for
S and S-PLUS. Springer, New York, USA. data exploration to avoid common statistical problems.
Pinheiro, J.C., Bates, D. M., DebRoy, S. & Deepayan, S., Methods in Ecology and Evolution 1:3–14. https://doi.
Team, R.C. 2020: nlme: linear and nonlinear mixed org/10.1111/j.2041-210X.2009.00001.x.You can also read

















































