Optimizing deep brain stimulation based on isostable amplitude in essential tremor patient models - IOPscience
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Neural Engineering
PAPER • OPEN ACCESS
Optimizing deep brain stimulation based on isostable amplitude in
essential tremor patient models
To cite this article: Benoit Duchet et al 2021 J. Neural Eng. 18 046023
View the article online for updates and enhancements.
This content was downloaded from IP address 46.4.80.155 on 09/05/2021 at 23:05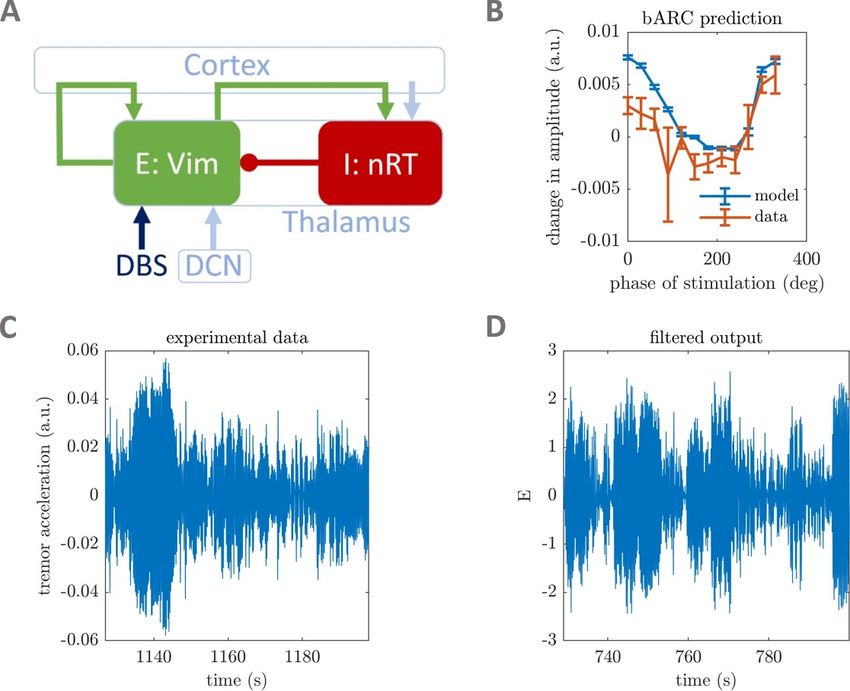
J. Neural Eng. 18 (2021) 046023 https://doi.org/10.1088/1741-2552/abd90d
Journal of Neural Engineering
PAPER
Optimizing deep brain stimulation based on isostable amplitude
OPEN ACCESS
in essential tremor patient models
RECEIVED
19 June 2020 Benoit Duchet1,2, Gihan Weerasinghe1,2, Christian Bick3,4,5 and Rafal Bogacz1,2
REVISED 1
16 December 2020 Nuffield Department of Clinical Neurosciences, University of Oxford, Oxford, United Kingdom
2
MRC Brain Network Dynamics Unit, University of Oxford, Oxford, United Kingdom
ACCEPTED FOR PUBLICATION 3
6 January 2021
Oxford Centre for Industrial and Applied Mathematics, Mathematical Institute, University of Oxford, Oxford, United Kingdom
4
Centre for Systems, Dynamics, and Control and Department of Mathematics, University of Exeter, Exeter, United Kingdom
PUBLISHED 5
EPSRC Centre for Predictive Modelling in Healthcare, University of Exeter, Exeter, United Kingdom
31 March 2021
E-mail: benoit.duchet@ndcn.ox.ac.uk
Original Content from Keywords: deep brain stimulation, essential tremor, isostable amplitude, Hilbert amplitude, stimulation strategy, pathological oscillations
this work may be used
under the terms of the Supplementary material for this article is available online
Creative Commons
Attribution 4.0 licence.
Any further distribution
of this work must Abstract
maintain attribution to
the author(s) and the title Objective. Deep brain stimulation is a treatment for medically refractory essential tremor. To
of the work, journal
citation and DOI.
improve the therapy, closed-loop approaches are designed to deliver stimulation according to the
system’s state, which is constantly monitored by recording a pathological signal associated with
symptoms (e.g. brain signal or limb tremor). Since the space of possible closed-loop stimulation
strategies is vast and cannot be fully explored experimentally, how to stimulate according to the
state should be informed by modeling. A typical modeling goal is to design a stimulation strategy
that aims to maximally reduce the Hilbert amplitude of the pathological signal in order to
minimize symptoms. Isostables provide a notion of amplitude related to convergence time to the
attractor, which can be beneficial in model-based control problems. However, how isostable and
Hilbert amplitudes compare when optimizing the amplitude response to stimulation in models
constrained by data is unknown. Approach. We formulate a simple closed-loop stimulation strategy
based on models previously fitted to phase-locked deep brain stimulation data from essential
tremor patients. We compare the performance of this strategy in suppressing oscillatory power
when based on Hilbert amplitude and when based on isostable amplitude. We also compare
performance to phase-locked stimulation and open-loop high-frequency stimulation. Main results.
For our closed-loop phase space stimulation strategy, stimulation based on isostable amplitude is
significantly more effective than stimulation based on Hilbert amplitude when amplitude field
computation time is limited to minutes. Performance is similar when there are no constraints,
however constraints on computation time are expected in clinical applications. Even when
computation time is limited to minutes, closed-loop phase space stimulation based on isostable
amplitude is advantageous compared to phase-locked stimulation, and is more efficient than
high-frequency stimulation. Significance. Our results suggest a potential benefit to using isostable
amplitude more broadly for model-based optimization of stimulation in neurological disorders.
1. Introduction stimulation (DBS) of the thalamus is an effective ther-
apy [5–7]. In its clinically available version, DBS deliv-
Essential tremor (ET) is a neurological disorder ers pulses of electrical stimulation at about 130 Hz via
believed to originate from aberrant neural synchrony electrodes implemented deep into the brain. Repor-
in the cerebellar-thalamic-cortical pathway [1]. ET ted side effects of high-frequency (HF) thalamic
is characterized by a tremor of the upper limbs, DBS include speech impairment, gait disorders, and
and tremulous hand movements are coherent with abnormal dermal sensations [8].
thalamic activity [2–4]. When medications are inef- Such side-effects due to HF stimulation, the
fective or not tolerated, high-frequency deep brain inability to adapt stimulation to symptom severity,
© 2021 The Author(s). Published by IOP Publishing Ltd
J. Neural Eng. 18 (2021) 046023 B Duchet et al
frequent battery replacement surgeries, as well as an attractor [37]). Isostables have been defined both
habituation in some patients (diminishing stimula- for limit cycle dynamics [38, 39] and fixed point
tion efficacy [9]) are some of the key limitations of (FP) dynamics [37], and have been applied to con-
HF DBS in ET. New closed-loop DBS strategies have trol problems in both cases. Outside of neuroscience,
therefore been explored to stimulate less while pre- examples include stopping cardiac alternans [40],
serving or even improving clinical efficacy. Examples controlling non-linear flows [41], and optimal con-
of closed-loop strategies include phase-locked DBS, vergence to a stable FP [42]. In [43], a stimulation
where stimulation is delivered according to the phase strategy is designed based on isostable reduction to
of pathological oscillations [10, 11], and adaptive desynchronize coupled neurons modeling patholo-
DBS, where stimulation is triggered based on the gical synchronization in PD. Besides isostables, meth-
amplitude of pathological oscillations [12–17]. There ods to define control strategies based on the off-
is no consensus on how to stimulate to best minimize line analysis of phase space have been suggested for
aberrant synchrony, and experimental testing is lim- instance in [44, 45].
ited because patients fatigue quickly. How isostable and Hilbert amplitudes compare
Among model-based control approaches in neur- when optimizing the amplitude response to stimula-
ological disorders and in particular in movement tion in models constrained by data is unknown. Since
disorders, strategies based on the phase depend- isostable amplitude delineates convergence time to
ence of the response to stimulation, strategies based the attractor, it provides information on the long-
on optimal control, and strategies based on feed- term behavior of the system, which may by useful
back control have been particularly influential. Phase in model-based studies of DBS. In this work, using
dependence of the response to stimulation has been models fitted in [32] to phase-locked stimulation data
utilized both at the individual oscillator level and at from ET patients [10], we ask the question ‘Can phase
the population level. The knowledge of individual space stimulation based on isostable amplitude be
oscillator phase response is used in [18–20] to design advantageous compared to phase space stimulation
stimuli aiming at desynchronizing coupled oscillat- based on Hilbert amplitude, and compared to simpler
ors. At the neural population level, a strategy consist- stimulation strategies?’ Because of the transient char-
ing in providing bursts of stimulation phase-locked to acter of tremor, the deterministic part of best model
the phase of population activity was suggested in [21]. fits from [32] gave rise to stable foci. We will therefore
Recently, a Bayesian adaptive dual control algorithm consider isostable amplitude of stable foci [37].
was designed to learn how to best stimulate according On the basis of stimulation strategies benefit-
to the phase and power of recorded oscillatory activ- ing from the knowledge of phase space amplitude
ity [22]. Optimal control has been used to steer neural fields, we show that stimulation based on isostable
systems to a particular target set, such as the phaseless amplitude can be beneficial compared to stimulation
set of a coupled oscillator model [23], or a target set based on Hilbert amplitude when computation time
determined through bifurcation analysis of a neuron is limited to minutes. We also compare the perform-
model [24]. Some feedback control strategies in neur- ance of stimulation based on isostable amplitude to
ological diseases have relied on the online estima- phase-locked stimulation and HF stimulation. Our
tion of a model [14, 25, 26]. Model predictive control results open opportunities for model-based optimiz-
has been applied to Parkinson’s disease (PD) using ation of stimulation in neurological disorders where
an extended autoregressive model [27], and various stimulation aims at minimizing the amplitude of an
forms of delayed feedback have also been investigated output.
[28, 29]. Further review can be found for instance
in [30, 31]. 2. Methods
Modeling studies such as [20, 32] have been car-
ried out to provide insights on how to effectively To compare phase space stimulation strategies, we
reduce the Hilbert amplitude of the pathological sig- consider previously fitted models, and their isostable
nal with stimulation in ET. The Hilbert amplitude and Hilbert amplitude fields. We also implement
is one of the most commonly used measures of phase-locked stimulation and HF stimulation as
instantaneous amplitude in neurophysiological data references.
[10, 11, 33–36]. It is defined as the modulus of the
analytic signal, and gives the amplitude of the envel- 2.1. Wilson–Cowan models fitted to ET patient
ope of narrow-band signals. data
On the other hand, the notion of isostables, We rely on neural mass models previously fitted in
which define an amplitude coordinate, has been [32] to tremor data from ET patients receiving phase-
applied with great success to control the state locked DBS [10]. In the latter study, a brief burst
of various systems. Trajectories starting on the of stimulation was delivered to the thalamus once
same isostable approach the attractor synchronously per tremor oscillation cycle, locked to the phase of
(isostables define a set of coordinates with codimen- tremor recorded from an accelerometer fitted to each
sion one associated with the decaying eigenvalues of patient (the data are available online [46]). In this
2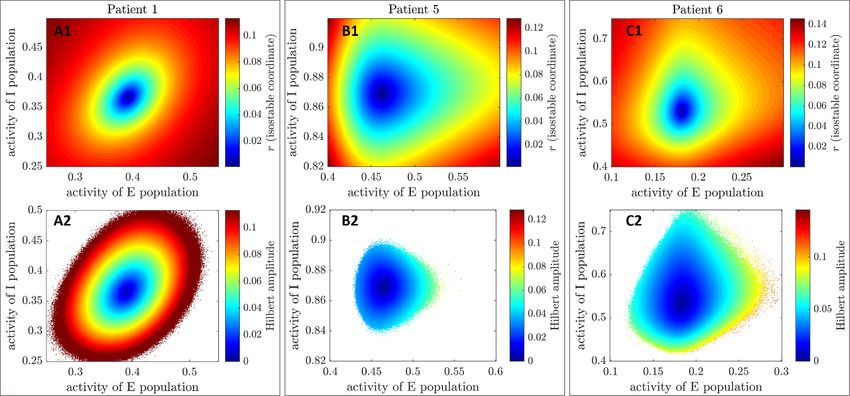
J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 1. Fitting WC models to phase-locked DBS tremor data from ET patients. The WC model can describe populations
thought to be involved in the generation of ET (A). Arrows denote excitatory connections or inputs, whereas circles denote
inhibitory connections. The VIM is the target of DBS and also receives an input from the deep cerebellar nuclei (DCN). The
self-excitatory loop of the VIM, as well as the excitatory connection from VIM to nRT are via cortex. Data and fitted model bARCs
for patient 5 are shown in panel (B) (patient numbers refer to [10]). Filtered tremor acceleration data (C) and fitted model output
(D) are also shown for patient 5. All four panels have been adapted from [32] under the creative commons CC BY 4.0 licence.
section, we review the results of [32] relevant to the 1
f(x) = ,
present study. The neural masses considered are two- 1 + e−β(x−1)
dimensional non-linear Wilson–Cowan (WC) mod-
parametrized by a steepness parameter β. In our (E, I)
els [47] with Gaussian white noise. The WC model
model, E is the key variable for two reasons. First,
depicts the interactions of a population of excitat-
DBS for ET is most commonly delivered to the VIM,
ory neurons, whose activity is denoted by E, and a
and we model stimulation by an increase δE in the
population of inhibitory neurons, whose activity is
activity of the corresponding population (E). Second,
denoted by I. The model can be mapped onto struc-
since the coherence between ventral thalamic activity
tures believed to be implicated in the generation of
and wrist flexor electromyographic recordings is high
ET as shown in figure 1, panel (A). The ventral inter-
[2–4], we use E to model the tremor signal.
mediate nucleus (VIM) of the thalamus is represen-
We consider in the present study the best fits to
ted by the excitatory population, while the reticular
the three ET patient datasets shown to have statistic-
nucleus (nRT) is represented by the inhibitory popu-
ally significant phase and amplitude response curves
lation. Their activities are given by the stochastic dif-
in [32], namely patient 1, patient 5, and patient 6.
ferential equations
Other patients were not fitted to in our previous
study as their phase-dependent response to stimu-
(
dE = 1
−E + f(ηE + wEE E − wIE I) dt + ζ dWE , lation was not statistically significant and was con-
ν sidered to be dominated by noise. The fitting pro-
dI = 1
ν −I + f(ηI + wEI E) dt + ζ dWI ,
cedure in our previous study subjected models to
(1) the same phase-locked stimulation paradigm used
in experimental data (see [32] for more details).
with wPR the weight of the projection from popula- The fits were based on features representing tremor
tion ‘P’ to population ‘R’, η P the constant input to dynamics (example of data and model tremor sig-
population ‘P’, and ν a time constant (assumed to be nals shown in panels (C) and (D) of figure 1), and
the same for both populations). We denote by W E the block phase response curve (bPRC, defined in
and W I Wiener processes, and ζ is the noise standard [32]). In addition to other model parameters, the
deviation. As in [48], the function f is the sigmoid stimulation magnitude was fitted to best reproduce
function these features. Here, we denote fitted stimulation
3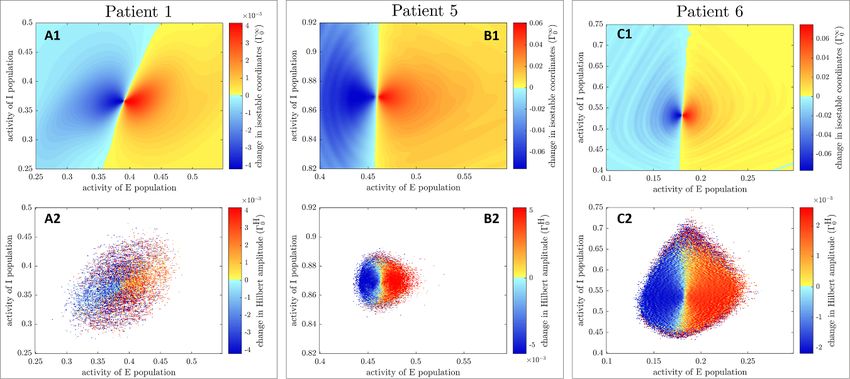
J. Neural Eng. 18 (2021) 046023 B Duchet et al
models. As mentioned earlier, the deterministic part
of the three fitted models give rise to stable foci.
2.2. Isostable amplitude of stable foci in 2D
To introduce isostable amplitude, we summarize the
concept of isostables for stable FPs presented in [37].
On an intuitive level, the isostables of a stable FP can
be defined as sets of points with the same asymptotic
convergence to the FP, i.e. sets of points approaching
the FP synchronously [37]. This is illustrated in the
animation provided in the online version of figure 2,
where trajectories starting on the same isostable reach
subsequent isostables at the same time. In what fol-
Figure 2. Isostables as sets of points with the same
asymptotic convergence. The animation corresponding to lows, we denote vectors in bold to distinguish scalars
this figure is available in the online version of this article. and vectors more easily. To provide a precise defini-
Trajectories starting on the same isostable are represented tion of isostables, let us consider a dynamical system
by dots of the same color and cross subsequent isostables at
the same time. Isostables are depicted by black contours.
This example shows the WC fit to patient 1. The state X
corresponds to (E, I). Ẋ = F(X), (2)
where X ∈ R2 for our purposes (see [37] for defin-
itions in Rn ), and the vector field F is analytic. The
magnitudes by δE0 and provide the fitted stimulation flow induced by equation (2) is denoted Φt (X). We
magnitude values from our previous study in table A assume that F has a stable FP which we denote X∗ ,
in the supplementary material (available online at with a basin of attraction B(X∗ ). We further assume
stacks.iop.org/JNE/18/046023/mmedia). As reported that the Jacobian J of F at X∗ has complex conjug-
in [32], the fitted models are able to reproduce ate eigenvalues λ± = σ ± iω with σ < 0. We denote
the data block amplitude response curves (bARCs, by v1 = a − ib and v2 the right normalized eigen-
defined in [32], example in figure 1, panel (B)), vectors associated with λ+ and λ− , respectively. The
without having been fit to the amplitude responses, isostable Iτ is defined in [37] when the leading eigen-
showing that the phase-dependence of the system value is not real as the one-dimensional manifold
response to stimulation has been captured by the
Iτ = X ∈ B(X∗ )|∃θ ∈ [0, 2π) s.t.
h i
lim e−σt Φt (X) − X∗ − < v1 ei(ωt+θ) eσ(t+τ ) = 0 , (3)
t→+∞
where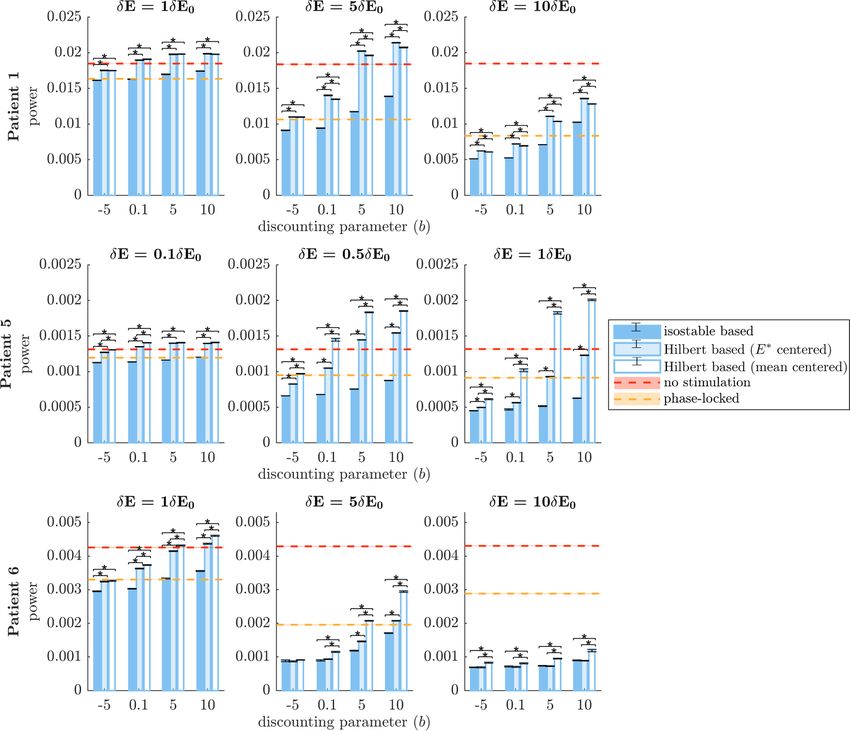
J. Neural Eng. 18 (2021) 046023 B Duchet et al
g2 : R2 7→ R are functions called observables. We section 3.3) is expected to translate into increased per-
define these observables as formance compared to simpler stimulation strategies
D h iE such as phase-locked stimulation. Thus we imple-
g1 = X − X∗ , −bb2 1 , g2 = hX − X∗ , [ −aa2 1 ]i , ment phase-locked stimulation as a comparison
h i for the phase space stimulation strategy presented
where a = [ aa12 ] and b = bb12 . These definitions sat- in section 3.3. The zero-crossing phase was used
isfy the three requirements on g 1 and g 2 of Propos- in [10] to investigate phase-locked stimulation in ET
ition 2 (ii) in [37]. Similarly to Laplace averages, this patients. We therefore provide phase-locked stimula-
method permits us to compute isostable coordinates tion during model integration according to the zero-
in the entire basin of attraction of the FP, but is less crossing phase which we estimate as in [32] (see
prone to numerical instability. appendix 7 therein). Briefly, we set the zero-crossing
phase to zero when positive zero-crossings are detec-
2.3. Obtaining Hilbert amplitude fields ted. Between zero-crossings, we evolve the phase lin-
Because the control goal is to reduce the Hilbert amp- early based on a frequency determined as the inverse
litude of the tremor, we use stimulation based on of the duration of the previous period. Given a selec-
Hilbert amplitude fields as a basis to compare the ted target phase for stimulation, one pulse of stimula-
performance of stimulation based on isostable amp- tion is delivered as soon as the target phase is reached.
litude fields. Hilbert amplitude fields can directly pre- In case the target phase has not yet been reached
dict the instantaneous impact of a given stimula- and the next positive zero-crossing is detected, it is
tion pulse in phase space on the Hilbert amplitude assumed that the phase has been underestimated and
of the tremor. We obtain Hilbert amplitude fields stimulation is provided right then. To maximize the
through averaging of numerical simulations (integ- benefits of phase-locked stimulation, the best target
ration time step is 1 ms). Specifically, we randomly phase for phase-locked stimulation is estimated from
draw 2000 initial positions in the phase space region simulations consisting of 5000 s of stimulation at
of interest. From these initial positions, we simulate one of twelve phase bins (same number of bins as in
the stochastic model (with the baseline noise level [10]). The phase bin resulting in the largest decrease
fitted to data) for 1000 periods. From each traject- in the power of the pathological oscillations is taken
ory, we obtain the Hilbert amplitude of the E com- as the target phase. This process is repeated for each
ponent as |e E(t) + iH(eE(t))|, where eE is the centered patient model and each stimulation magnitude stud-
E component, and H denotes the Hilbert trans- ied in section 3.4. For each condition, the stimulation
form. We clip 0.5% of the trajectories and the Hil- magnitude is matched to what is used in phase space
bert amplitude time series at both ends to remove stimulation.
edge effects. To reduce the effect of noise, we lightly We also compare the energy delivered by HF
smooth the trajectories and Hilbert amplitude time stimulation and by our amplitude-based phase space
series (moving average spanning four integration strategies for the same clinical effect. HF stimulation
time steps). The (E, I) region of interest is discretized is implemented by providing stimulation pulses at
into 0.001 × 0.001 unit bins (0.001 × 0.0002 units for 130 Hz in an open-loop fashion. Using optimization
patient 5 whose I range is notably narrower). For each (specifically the generalized pattern search algorithm
point in each trajectory, the corresponding space bin [49, 50]), we determine the stimulation pulse mag-
is found, and the associated Hilbert amplitude value nitude required by HF stimulation to attain the effic-
is added to the bin. The Hilbert amplitude field over acy of the best performing phase space stimulation
the region of interest is finally obtained by averaging strategy (within ±1%). This procedure is performed
Hilbert amplitude values within space bins. for each case presented in section 3.4, and six tri-
To investigate differences between isostable and als of 5000 s of HF stimulation are generated at each
Hilbert amplitude based stimulation, we use two ways optimization step.
of centering the E component to obtain e E, resulting While HF DBS has been previously investigated
in two sets of Hilbert amplitude fields. The first way in deterministic WC models in the limit cycle regime
is to center E by removing its mean, which is what [51, 52], its mechanism of action in our stable fixed-
is commonly done to obtain Hilbert amplitude. This point stochastic WC model is different. Stimulation at
results in what we call a ‘mean-centered Hilbert amp- 130 Hz is fast enough compared to intrinsic dynam-
litude field’. The second way is to center E by using ics that HF stimulation is similar in the model to
information from phase space and removing E* (the stimulation provided continuously (with an appro-
first coordinate of the FP). This results in what we call priately reduced stimulation magnitude to maintain
a ‘FP-centered Hilbert amplitude field’. the total energy delivered). Thus the effect of HF
stimulation can be illustrated by increasing the rate
2.4. Phase-locked stimulation and high-frequency of change of the E population activity (originally
stimulation given by equation (1)) by a constant. This modifies
The additional information used in stimulation the vector field, and in our patient models, the vec-
strategies based on amplitude fields (introduced in tor field with HF stimulation results in trajectories
5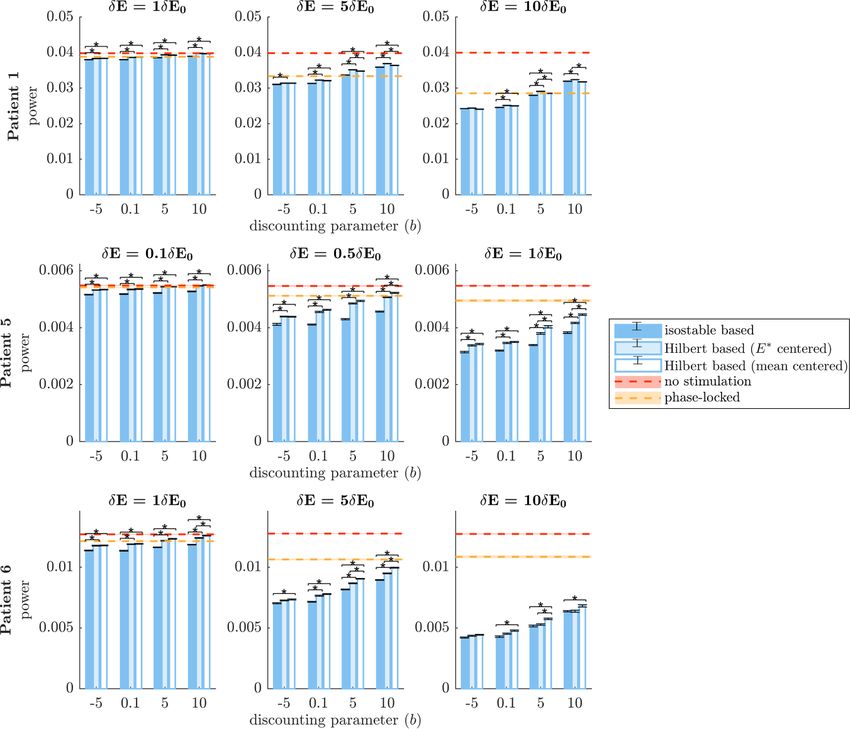
J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 3. Isostable and mean-centered Hilbert amplitude fields (full estimates) for the three WC patient fits. Each column
corresponds to a patient fit. Isostable amplitude fields are presented in the top row, and mean-centered Hilbert amplitude fields in
the bottom row. Colour scales are matched on a patient basis. For the WC model, the state X corresponds to (E, I), where E is the
activity of the excitatory population, and I the activity of the inhibitory population.
of lower E amplitude when stimulation is strong basin of attraction of the FP, Hilbert amplitude could
enough. not be determined in white zones in panels (A2),
(B2) and (C2) of figure 3. FP-centered Hilbert amp-
3. Results litude fields (figure A in the supplementary material)
are very similar to mean-centered Hilbert amplitude
We suggest a closed-loop stimulation strategy relying fields. We call the computationally intensive, high
on phase space amplitude fields and compare the per- quality estimates of isostable and Hilbert amplitude
formance of this strategy when the amplitude field fields presented in this section ‘full estimates’.
used portrays isostable amplitude to when it depicts
Hilbert amplitude. In both cases, the goal of the stim-
ulation strategy is to reduce as much as possible the 3.2. Quick estimates of amplitude fields
power of pathological oscillations. The high quality estimates of amplitude fields
obtained in the previous section require up to ten
3.1. Full estimates of amplitude fields hours of computation time for Hilbert amplitude
Reliable estimates of isostable amplitude fields for the fields, and on the order of one hour of computa-
three WC patient fits (figure 3, panels (A1), (B1), tion time for isostable amplitude fields (using no
and (C1)) were obtained using equation (4) and an more than four threads in both cases). It is of prac-
explicit Runge–Kutta (4,5) method to integrate tra- tical interest to obtain quicker estimates of amplitude
jectories. Isostable amplitude fields are given for the fields, and to compare the performance of isostable
deterministic version of the fitted models (i.e. with and Hilbert amplitude fields requiring similar com-
ζ = 0). As n is increased, convergence of isostable putational time. Indeed, amplitude fields may need to
coordinates happens before numerical instability sets be updated often in clinical practice. We obtain quick
in (this was not the case with the first order Euler’s estimates of isostable amplitude fields by relaxing the
method). We used n = 80 for patient 1, n = 60 for convergence requirement of the isostable field. This
patient 5, and n = 120 for patient 6. For these val- enables us to use the faster Euler’s method to integ-
ues, r has converged and there is no trace of numer- rate trajectories. We select on a patient basis n (see
ical instability. Since the fitted models reproduce equation (4)) as well as the time step to lower compu-
the particular tremor dynamics and phase-dependent tation time as much as possible while avoiding major
response to phase-locked DBS of each individual artefacts in the isostable amplitude field. We then
patient [32], differences in amplitude fields between approximately match on a patient basis the computa-
subjects should reflect patient specificities. tion time taken to compute Hilbert amplitude fields
Hilbert amplitude fields are estimated as by reducing the number of simulations and num-
described in section 2.3 for the three WC patient fits. ber of periods per simulations (the Euler–Maruyama
Mean-centered Hilbert amplitude fields (figure 3, method is already used). Single threaded computa-
panels (A2), (B2), and (C2)) are strikingly similar in tion times range from 19 to 454 s and are detailed in
shape to isostable amplitude fields. While isostable table B in the supplementary material together with
amplitude can easily be computed anywhere in the the parameters used. The resulting amplitude fields
6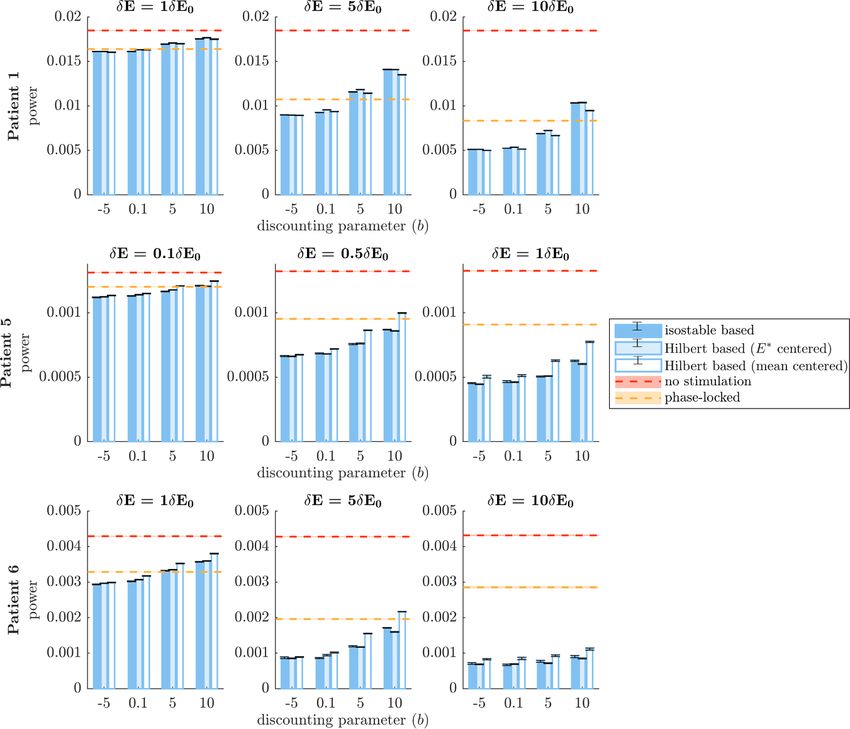
J. Neural Eng. 18 (2021) 046023 B Duchet et al
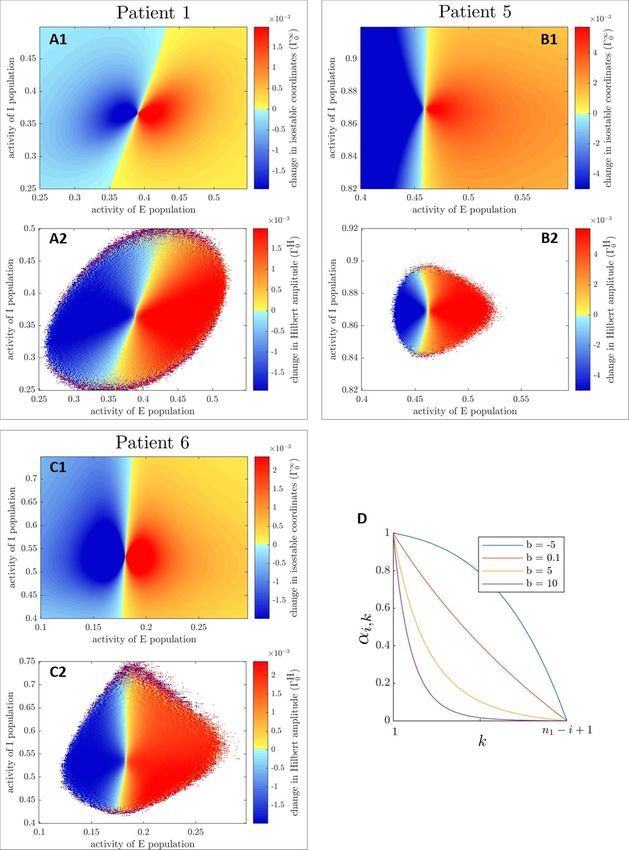
Figure 4. Instantaneous amplitude response fields for full estimates of asymptotic amplitude (Γ∞ 0 , panels (A1), (B1) and (C1))
and full estimates[of mean-centered
] Hilbert amplitude (ΓH
0 , panels (A2), (B2), and (C2)) are shown for the three fitted WC
models for δX = δ0E0 (δE0 are previously fitted stimulation magnitudes). White zones signify missing values. Negative values of
the response fields (in blue) signify a decrease in amplitude (beneficial stimulation), positive values (in yellow to red) signify an
increase in amplitude. Color scales are matched on a patient basis. Panel D illustrates the k − 1 step ahead discount factor αi,k at
time t i , for various values of the discounting parameter b. Time tn1 corresponds to the predicted end of the current period.
are similar to full estimates (see figure B in the sup- 3.3.1. Instantaneous and augmented response fields
plementary material). However the scale of isostable The starting point is a phase space amplitude field
amplitude fields is not the same since the conver- Ω(X) (we will be using isostable or Hilbert amp-
gence requirement has been relaxed. Hilbert amp- litude fields shown in figure 3, as well as in figures A
litude fields are noisier, with more missing values. and B in supplementary material). From there, we
define the corresponding ‘instantaneous amplitude
response field’ Γ0 (X) = Ω(X + δX) − Ω(X), where
3.3. A phase space stimulation strategy drawing on δX is the stimulation vector. Instantaneous amplitude
amplitude fields response fields for isostable amplitude (Γ∞ 0 ) and
We describe a stimulation strategy that uses inform- mean-centered Hilbert amplitude (ΓH 0 ) are presen-
ation from state space amplitude fields to decide ted in figure 4, panels (A)–(C). Zones where stimula-
whether it is more beneficial to stimulate now or at tion is beneficial correspond to negative values of the
a later time of the ongoing period. response fields (decrease in amplitude) and are shown
7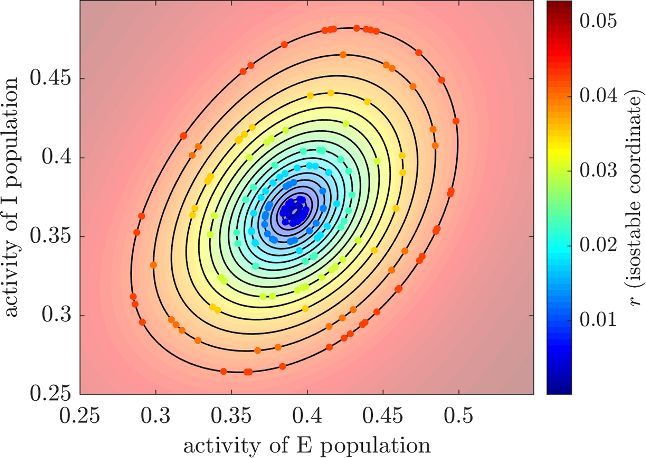
J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 5. Instantaneous amplitude response fields for quick estimates of asymptotic amplitude (Γ∞ 0 , panels (A1), (B1) and (C1))
H
and quick estimates
[ ] of mean-centered Hilbert amplitude (Γ 0 , panels (A2), (B2), and (C2)) are shown for the three fitted WC
models for δX = δ0E0 (δE0 are previously fitted stimulation magnitudes). White zones signify missing values. Negative values of
the response fields (in blue) signify a decrease in amplitude (beneficial stimulation), positive values (in yellow to red) signify an
increase in amplitude. Color scales are only matched for patient 1 as field values are too different for other patients.
in blue (the darker the blue, the greater the mag-
Algorithm 1. Obtain Γ
nitude of the change in amplitude). Conversely, zones
where stimulation is not beneficial correspond to pos- for (i, j) in {1, . . . , NE } × {1, . . . , NI } do
itive values of the response fields (increase in amp- simulate trajectories (E(tk ), I(tk ))k∈{1,...,u}
litude) and are indicated with a colorscale from yel- with t 1 = 0, t u = T, E(t1 ) = Ei , and I(t1 ) = Ij .
low to red. Instantaneous amplitude response fields for k in {1, …, u} do
find (p, q) ∈ {1, . . . , NE } × {1, . . . , NI } s.t.
for FP-centered Hilbert amplitude (which we denote
∗ (Ep , Iq ) is the closest to (E(tk ), I(tk ))
ΓH H
0 ) are very similar to Γ0 (see figure C in the if Γ0 (p, q) < 0 and Γ0 (p, q) ⩽ Γ0 (i, j) then
supplementary material). Instantaneous amplitude Γ(i, j, k) ← Γ0 (p, q)
response fields from quick estimates of Hilbert amp- else
litude fields are more noisy (mean-centered Hilbert Γ(i, j, k) ← 0
amplitude in the bottom row of figure 5, and FP- end if
centered Hilbert amplitude in figure D in the sup- end for
plementary material). Although quick estimates of end for
isostable amplitude fields do not exhibit visible arte-
facts (top row of figure B in the supplementary mater-
ial), patterns that were not present in figure 4 are seen of no interest to provide beneficial stimulation and
in the instantaneous response fields corresponding to are logged in Γ as zeros.
quick estimates of isostable amplitude fields. These
patterns include deformations of constant field value 3.3.2. Phase space stimulation using amplitude fields
contours (contrast the rightmost part of figure 4(A1) Based on an augmented instantaneous amplitude
and of figure 5(A1)), as well as ripples organized in a response field of choice, the decision to stimulate or
concentric fashion (compare figures 4(C1)–5(C1)). not is made at each model integration time step using
In what we call an ‘augmented instantaneous the current position in phase space (algorithm 2).
amplitude response field’ Γ(X, t), we embed the val- Stimulation is provided as a single pulse, at maximum
ues of Γ0 on deterministic trajectories starting at X once per period according to the scheme. We track
from t = 0 to t = T. The information will be used zero-crossings and estimate the zero-crossing phase
to inform the decision to stimulate or not at X. We (as detailed in section 2.4). To prevent instability
are considering the (E, I) phase space, and the range when zero-crossing tracking fails, we enforce a
of interest of E and I are discretized into N E and maximum stimulation frequency (corresponding to
N I values, respectively. Algorithm 1 describes how Nlim time steps in algorithm 2) of 10 Hz, which is
we obtain Γ from Γ0 in discretized space and time. roughly twice the tremor frequency. The benefit of
Stimulation is only beneficial when the amplitude stimulating at later times of the ongoing period is
response is negative (decrease in pathological signal estimated by the values stored in Γ in the space bin
amplitude, tremor amplitude). Positive values of the corresponding to the current position. The estim-
instantaneous amplitude response field are therefore ated values are based on deterministic trajectories,
8J. Neural Eng. 18 (2021) 046023 B Duchet et al
∗
algorithm 2 based on Γ∞ H
0 , on Γ0 , or on Γ0
H
to
Algorithm 2. Decide to stimulate at time step t i
the three fitted models for various stimulation mag-
if no stimulation for Nlim time steps and no stimulation nitudes δE (given as multiples or fractions of δE0 ),
yet in current period then and for various values of the discounting parameter b.
find (p, q) ∈ {1, . . . , NE } × {1, . . . , NI } s.t. (Ep , Iq ) Hilbert amplitude instantaneous response fields have
is the closest to (E(ti ), I(ti )) space bins with missing values (see for instance white
if Γ(p, q, 1) < 0 then
zones in figure 4, panels (A2), (B2), and (C2)). For fair
if predicted period end already reached then
stimulate
comparison, the corresponding space bins are emp-
else tied in isostable amplitude instantaneous response
if Γ(p, q, 1) < min (αi,k Γ(p, q, k)) then fields. For compatibility with algorithm 2, Γ0 is set
k∈{2,...,n1 −i+1}
to zero in both cases for empty space bins, so that
stimulate
end if stimulation cannot be triggered at these locations.
end if Smaller stimulation magnitude ratios δE/δE0 are used
end if for patient 5 because of the comparatively small
end if size of the Hilbert amplitude field available for this
patient and the comparatively larger δE0 (see table A
in the supplementary material). To compare Hilbert
whereas the model includes Gaussian white noise. and isostable amplitude stimulation as well as phase-
As a result, the actual benefit of waiting to stimu- locked stimulation, we measure stimulation efficacy
late is uncertain and will deviate from the average by integrating the power spectrum density (PSD) of
(noiseless) trajectory prediction, while the benefit of the E activity (which models the tremor signal). Effic-
stimulating now is known exactly. Future estimates acy is averaged using 30 trials of 5000 s of stim-
must therefore be discounted. From time t i , we define ulation. Since repeated measures are performed on
the k − 1 step ahead discount factor αi,k as each patient, linear mixed effect models with ran-
" # dom intercepts are used to assess statistical differences
1 1 between phase space methods while accounting for
αi,k = b
− b
(ti+k−1 − tn0 ) (tn1 − tn0 ) within patient dependence. Specifically, we fit to each
" #−1 scenario the model
1 1
× − , (5)
(ti − tn0 )
b
(tn1 − tn0 )
b
power ∼ method ∗ stim ∗ b + (1|patient), (6)
where b is the discounting parameter, n0 is the index where method, stim, and b are the factors corres-
of the first time step of the current period, and n1 is ponding to the choice of amplitude field, the stim-
the index of the last time step of the current period. ulation magnitude ratio δE/δE0 , and the discount-
As shown in panel (D) of figure 4, αi,k is one at t i (no ing parameter b, respectively. Following standard R
discounting of the present value), and zero at the pre- notation, ∗ indicates the inclusion of all main effects
dicted end of the current period. It is also apparent and interactions between the factors, and (1|patient)
that a larger b discounts more heavily future values. indicates a random intercept per patient. We eval-
In algorithm 2, discount factors are not used if the uate the significance of fixed effects using F-tests
end of the current period predicted by zero-crossing based on Satterthwaite’s method for denominator
phase has been reached before the next zero-crossing degrees-of-freedom [53]. Where the method factor
is detected. In this case, stimulation is applied right is significant, post hoc comparisons between the
then if beneficial. three phase space methods are performed using t-
tests with Bonferroni correction for robust multiple
3.4. Performance of phase space stimulation based comparisons.
on various amplitude fields
Using the three fitted patient models, we compare the 3.4.1. Phase space stimulation based on full estimates
performance of algorithm 2 when based on isostable of amplitude fields
amplitude, on mean-centered Hilbert amplitude, or Comparison of phase space stimulation based on
on FP-centered Hilbert amplitude under three scen- amplitude field full estimates in figure 6 shows that
arios. As a reference, the performance of phase- stimulation based on isostable amplitude is mostly
locked stimulation (see section 2.4) is also repor- on par with stimulation based on Hilbert amplitude,
ted for each scenario, and an efficiency comparison and that phase space methods can be more effect-
with HF stimulation is provided. We consider the ive than phase-locked stimulation. The method factor
cases of full estimates of the amplitude fields (see comparing phase space strategies was not signific-
section 3.1) and of quick estimates of the amplitude ant overall in the corresponding mixed effect model
fields (see section 3.2). We also investigate the influ- (p = 0.0605, see table C in the supplementary material
ence of doubling the noise standard deviation ζ (for for more details). Post hoc comparisons were there-
full estimates only). Under each scenario, we apply fore not performed. The slight advantage of isostable
9J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 6. Comparison of the efficacy of phase space stimulation strategies based on full estimates of amplitude fields. Isostable
amplitude fields correspond to darker blue bars, FP-centered Hilbert amplitude fields to lighter blue bars, and mean-centered
Hilbert amplitude fields to white bars. Each row corresponds to a WC patient fit. Results are shown for a range of stimulation
magnitude ratios δE/δE0 (increasing from left to right), and a range of discounting parameters b. The vertical axes represent
efficacy measured as the integral of the PSD of E (which models the tremor signal). A no stimulation reference is indicated by red
dashes, and phase-locked stimulation is indicated by orange dashes. Averages correspond to 30 trials of 5000 s of stimulation.
SEM error bars are shown in black for Hilbert and isostable based stimulation and as shading for no stimulation and phase-locked
stimulation.
amplitude for patients 5 and 6 for larger values of b and its potential for a lower occurrence of side-effects
compared to mean-centered Hilbert amplitude seems compared to HF stimulation.
to be due to a more favorable centering of the isostable
amplitude field. Indeed, the difference disappears for 3.4.2. Robustness to noise
FP-centered Hilbert amplitude. Furthermore, we have Robustness to noise was evaluated by doubling the
H∗
confirmed that the difference between ΓH 0 and Γ0 noise standard deviation in patient models. PSDs
∗
can be reproduced simply by shifting ΓH 0 along the E of model simulations for the three fitted patients
dimension (see figure E in the supplementary mater- are shown in the baseline and increased noise con-
ial). Despite similar performance under this scenario, ditions in figure G in the supplementary material.
∗
there are still non negligible differences between ΓH 0 The main PSD peak width is substantially enlarged
and Γ∞ 0 as evidenced by figure F in the supplementary in the increased noise condition, in particular for
material. Additionally, we note that our phase space patients 5 and 6. In this scenario using full estim-
stimulation strategies based on amplitude fields are ates of amplitude fields, the method factor of the
more effective than phase-locked stimulation for low corresponding mixed effect model was significant
discounting parameters and high stimulation mag- (p < 10−15 , see table C in the supplementary mater-
nitudes. To match the efficacy of stimulation based ial for more details). Significance of post-hoc com-
on isostable amplitude at b = −5, open-loop HF stim- parisons under Bonferroni correction is indicated by
ulation has to deliver from 6.4 times to 136 times black stars in figure 8. There is an overall reduction
more energy to the thalamus (see round markers in in efficacy of all stimulation strategies compared to
figure 7). This highlights the greater energy efficiency the baseline noise condition (figure 6), including
of phase space stimulation based on amplitude fields for phase-locked stimulation. As before, phase space
10J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 7. Comparison of the energy delivered to the thalamus for the same clinical effect by the best performing strategy based on
isostable amplitude and by HF stimulation. The vertical axis represents the ratio of the energy delivered by HF stimulation to the
energy delivered by phase space stimulation based on isostable amplitude (discounting parameter b = −5) for the same efficacy in
the model. Efficacy is measured as the integral of the PSD of E, which models the tremor signal, and the efficacy match between
stimulation types is within ±1%. The horizontal axis represents the increasing stimulation magnitude ratios δE/δE0 used for
isostable stimulation. Each color corresponds to a WC patient fit, and each marker type to a comparison scenario (see
sections 3.4.1–3.4.3).
stimulation strategies based on amplitude fields are estimates of amplitude fields which can be com-
more effective than phase-locked stimulation for puted in minutes is compared in figure 5. In this
b = −5 in all the cases shown in figure 8 (although scenario, the method factor of the corresponding
the difference is small for the lowest stimulation mixed effect model was found significant (p < 10−15 ,
case in patient 1), and for a broader range of dis- see table C in the supplementary material for more
counting parameters at high stimulation magnitude. details). Isostable amplitude based stimulation offers
Among phase space stimulation strategies, stimula- a clearer advantage compared to Hilbert amplitude
tion based on isostable amplitude outperforms stim- based stimulation (see post-hoc comparison under
ulation based on Hilbert amplitude in a number of Bonferroni correction in figure 8) than in previous
cases (patient 5, and in most conditions investigated scenarios. While instantaneous amplitude response
for patient 6). However these differences are small, fields based on quick estimates of isostable amplitude
and are even smaller or not favorable for patient 1. exhibit some artefacts (see top panels in figure 5),
When the noise standard deviation is larger, trajector- these artefacts do not seem to notably affect the
ies cover more of phase space. Because Hilbert amp- performance of isostable based stimulation. In fact,
litude fields rely on averaging, zones closer to the the efficacy of phase space stimulation based on
edges of Hilbert amplitude fields are noisier as tra- quick estimates of isostable amplitude and the effic-
jectories of large amplitude are more sparse. Isostable acy of phase space stimulation based on full estim-
amplitude fields are immune to this issue, which ates of the isostable amplitude are similar (com-
may explain the slightly better robustness of isostable pare the darker blue bars in figures 6 and 9). In
based stimulation to an increase in the noise stand- contrast, the increased noise in instantaneous amp-
ard deviation. Despite the increased noise level, stim- litude response fields based on quick estimates of
ulation based on amplitude fields is still more effi- Hilbert amplitude (see bottom panels in figure 5, as
cient than open-loop HF stimulation. To match the well as figure D in the supplementary material) is
efficacy of stimulation based on isostable amplitude responsible for the decreased performance of Hilbert
at b = −5, open-loop HF stimulation has to deliver amplitude based stimulation. Even when isostable
from 4.5 times to 120 times more energy to the thal- amplitude fields are computed in minutes, isostable
amus (see square markers in figure 7). amplitude based stimulation (and in fewer cases Hil-
bert amplitude based stimulation) is as before more
effective than phase-locked stimulation, in particu-
3.4.3. Phase space stimulation based on quick lar for low discounting values (with the exception of
estimates of amplitude fields patient 1 at the lowest stimulation level, where per-
In clinical practice, full estimates of amplitude fields formance is similar). Additionally, stimulation based
are unlikely to be useful because of their compu- on isostable amplitude fields computed in minutes is
tational cost (on the order of hours). The per- still more efficient than open-loop HF stimulation,
formance of stimulation strategies based on quick requiring from 6.6 times to 132 times less energy to
11J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 8. Comparison of the efficacy of phase space stimulation strategies based on full estimates of amplitude fields when
doubling the noise intensity. Isostable amplitude fields correspond to darker blue bars, FP-centered Hilbert amplitude fields to
lighter blue bars, and mean-centered Hilbert amplitude fields to white bars. Each row corresponds to a WC patient fit. Results are
shown for a range of stimulation magnitude ratios δE/δE0 (increasing from left to right), and a range of discounting parameters b.
The vertical axes represent efficacy measured as the integral of the PSD of E (which models the tremor signal). A no stimulation
reference is indicated by red dashes, and phase-locked stimulation is indicated by orange dashes. Averages correspond to 30 trials
of 5000 s of stimulation. SEM error bars are shown in black for Hilbert and isostable based stimulation and as shading for no
stimulation and phase-locked stimulation. Black stars denote significant differences under Bonferroni correction.
be delivered to the thalamus (see triangular markers Hilbert amplitude fields significantly degraded due
in figure 7). to noisier estimates. This suggests that there might
be a benefit to using more broadly amplitude fields,
4. Discussion in particular isostable amplitude fields, for model-
based optimization of stimulation in neurological
In this study, we showed that phase space stimula- disorders.
tion based on isostable or Hilbert amplitude fields
can be more effective than phase-locked stimulation
and is more efficient than open-loop HF stimulation, 4.1. Isostable amplitude
even when noise is increased from the baseline noise Although isostables have been developed with model
level fitted to patients. Stimulation based on isostable reduction in mind [37–39], there are similarities
amplitude was in most cases on par with stimula- between isostable amplitude and Hilbert amplitude,
tion based on Hilbert amplitude, with slight favorable which is one of the preferred measures of synchrony
differences accounted for by a more accurate center- in neurophysiological data. Trajectories starting from
ing of isostable amplitude fields. The performance of the same isostable converge synchronously to the
isostable amplitude based stimulation and its advant- attractor, i.e. reach subsequent isostables at the same
age over phase-locked stimulation and HF stimula- time. In our (E, I) model, trajectories starting from
tion was maintained when using quick estimates of the same isostable share the same Hilbert amplitude
amplitude fields obtained in minutes, which is likely (true regardless of whether the Hilbert amplitude is
to be a clinical requirement. In contrast, the per- defined based on E or on I). This is reflected by
formance of stimulation based on quick estimates of the fact that isostable and Hilbert amplitude fields
12J. Neural Eng. 18 (2021) 046023 B Duchet et al
Figure 9. Comparison of the efficacy of phase space stimulation strategies based on quick estimates of amplitude fields. Isostable
amplitude fields correspond to darker blue bars, FP-centered Hilbert amplitude fields to lighter blue bars, and mean-centered
Hilbert amplitude fields to white bars. Each row corresponds to a WC patient fit. Results are shown for a range of stimulation
magnitude ratios δE/δE0 (increasing from left to right), and a range of discounting parameters b. The vertical axes represent
efficacy measured as the integral of the PSD of E (which models the tremor signal). A no stimulation reference is indicated by red
dashes, and phase-locked stimulation is indicated by orange dashes. Averages correspond to 30 trials of 5000 s of stimulation.
SEM error bars are shown in black for Hilbert and isostable based stimulation and as shading for no stimulation and phase-locked
stimulation. Black stars denote significant differences under Bonferroni correction.
are strikingly similar (see figure 3). Although amp- which is not the case in the present work. Benefits
litude values and gradients are different, contours of of the isostable method chosen [37] include its com-
equal amplitude have very similar shapes. In R2 , a putational efficiency, and its truly global character.
relationship between isostable and Hilbert amplitude Isotables can be computed everywhere in the basin
might be revealed by studying the approximation of attraction of the FP without limitations. The the-
of isostable coordinates given by equation (4). This ory behind isostables considers purely deterministic
equation can be interpreted as the norm of a 2D vec- dynamical systems, whereas the fitted WC models
tor, similarly to the Hilbert amplitude. The Hilbert employed in our study involve Gaussian white noise.
amplitude is indeed the norm of the vector formed by Theories have been developed to account for the effect
the real and the imaginary parts of the analytic signal. of noise in isochrons [57–59], but isostables are lag-
In dynamical systems, phase-amplitude reduc- ging behind in this respect. However, isostable amp-
tions such as the isochron-isostable reduction are litude fields share similarities with Hilbert amplitude
augmentations of classical phase reductions, and vari- fields which were obtained by averaging noisy traject-
ous techniques are available to define an amplitude ories. It therefore seems likely that isostable amplitude
coordinate (see [54] for a review). In contrast with contours would not change substantially were Gaus-
methods based on isostable amplitude, the euclidean sian white noise explicitly accounted for.
distance to the attractor is used as an amplitude vari- Control strategies developed in [18–20] are based
able in [55], and a transverse variable obtained from on the knowledge of individual oscillator phase
parametrizing isochrons is used in [56]. These meth- response curves, likely challenging to obtain in
ods are applicable when the attractor is a limit cycle, patients. In [21], phase synchronization of neural
13J. Neural Eng. 18 (2021) 046023 B Duchet et al
populations is directly considered. However the amp- in most cases. This may be due to the fact that Hilbert
litude response of individual populations, which may amplitude fields are not as smooth (see figure 3) since
be of importance at the mesoscale level, is not taken they are obtained by averaging stochastic trajectories.
into account. Instead of considering phase synchron- For quick estimates, because stimulation maps based
ization of oscillators, we directly focus on amplitude on Hilbert amplitude are not as reliable as for full
response at the population level, which is more dir- estimates, stimulation is more likely to be delivered in
ectly related to symptoms. unfavorable phase space locations as seen in (C) pan-
els in figures H–J (supplementary material). Finally,
4.2. Comparing phase space stimulation strategies isostable and Hilbert amplitude response fields do not
based on isostable and Hilbert amplitude exhibit the same boundaries between regions where
The closed-loop phase space stimulation strategy sug- stimulation is beneficial and regions where it is not.
gested in section 3.3 produces overall similar results This is particularly true in patients 5 and 6 (see for
when based on full estimates of isostable amplitude instance figure 4, panels (A)–(C)).
and when based on full estimates of Hilbert amp-
litude. Minimizing isostable amplitude is minimizing 4.3. Limitations of the study
convergence time to the attractor. Knowledge of the An important limitation of this study is that because
future evolution of the (deterministic) non-linear sys- only three cases with a statistically significant phase
tem embedded in isostable coordinates may theoret- response were available, our comparison is based
ically provide an advantage in controlling the Hilbert on these three cases only. Because of experimental
amplitude of the stochastic system. Instantaneous difficulties, patient recordings are short, and non-
response fields obtained from isostable amplitude are significant patients might simply have required more
in fact different from their Hilbert amplitude coun- data. However essential tremor is an heterogeneous
terparts, even when FP-centered Hilbert amplitude disorder, and it might also be the case that cer-
is used (see figure F in the supplementary material). tain patient subtypes respond better to phase-locked
However convergence time information captured by stimulation. Nevertheless, the three patient models
isostable amplitude does not seem to directly translate studied are heterogeneous as shown by notably dif-
into major differences in performance with Hilbert ferent amplitude fields from one patient to another
amplitude based stimulation in the two-dimensional (figure 3), suggesting that phase space stimulation
models studied (see figure 6). The benefit of using based on amplitude fields may be applicable to vari-
isostable amplitude fields in two-dimensional mod- ous patient subtypes.
els may reside instead in the ability to obtain quick While the WC model is useful to model DBS
estimates of amplitude maps more reliably than with in ET, it is worth discussing its limitations in the
Hilbert amplitude. This is reflected in the better relat- context of the literature. The central origin of ET
ive performance of isostable amplitude based stimu- is well established (see e.g. [1, 60, 61]). In particu-
lation in figure 9. However Hilbert amplitude based lar, thalamic activity has been associated with activity
stimulation is still competitive in some cases, such in peripheral muscles driving the tremor [2–4], and
as for low discounting parameter and high stimula- some ET models are exclusively based on biophys-
tion for patients 5 and 6. It is worth emphasizing ically detailed representations of thalamic neurons
that the resolution and coverage that can obtained for [62, 63]. Other models include the broader cerebello-
isostable amplitude fields is to our best knowledge out thalamo-cortical network [51, 64, 65]. Since detailed
of reach for Hilbert amplitude fields. To obtain Hil- knowledge of the central mechanism of generation
bert amplitude fields, considering noisy trajectories of ET is not available, mean-field models are natural
is necessary to explore more of phase space, which candidates to study ET [20, 32, 51]. The WC model
drastically limits the resolution that can be obtained used in this study is a heuristically derived mean-field
for a given number of simulations. To level the playing model [47]. As such, it benefits from a low number of
field for our comparison, isostable amplitude fields parameters while retaining some level of description
were masked to match the coverage of Hilbert amp- of a microscopic biological reality. However, bursting
litude fields, and the same resolution was used for of thalamic neurons is not described by the WC model
both amplitude fields. and might play a role in ET [51]. Another limita-
Differences in where stimulation is provided in tion is that our WC model portrays a central feedback
phase space based on isostable or Hilbert amplitude loop, and does not account for the interaction of the
fields are highlighted in figures H, I, and J in the sup- centrally generated component of ET with the mech-
plementary material. Stimulation based on isostable anical stretch reflex, whose involvement may depend
amplitude tends to happen a little later in phase, on the disease state [66, 67]. The mesoscopic scale of
although this effect is diminished when comparing the WC model makes it particularly suited to be fit-
isostable amplitude to FP-centered Hilbert amplitude ted to mesoscopic data such as local field potential
instead of mean-centered Hilbert amplitude (see for recordings obtained from DBS electrodes, or tremor
instance panel (A) and (B) in figure I). Isostable amp- recordings (as used in [32]). It is likely that more com-
litude based stimulation also tends to be more focal plex models with more parameters would overfit such
14J. Neural Eng. 18 (2021) 046023 B Duchet et al
data, and would be too computationally demanding can run very quickly (real time is likely to be achiev-
to fit in practice. Despite the limitations mentioned able). While the model state would need to be estim-
above, the WC model has been shown to be adept ated in near real time, we expect model paramet-
at reproducing the effects of high-frequency DBS in ers to change more slowly. Model parameters would
ET [51] and PD [52], as well as the phase-dependent therefore be updated and the amplitude fields (and
effects of phase-locked DBS in ET in datasets with corresponding response fields) computed on this
statistically significant response curves [32]. Addi- slower time scale, which may be on the order of
tionally, our phase space stimulation strategies based minutes. This is compatible with our results based
on amplitude fields can be applied to more complex on quick estimates of amplitude fields (see table B
models than the WC model. The isostable compu- in the supplementary material for single threaded
tation method described in section 2.2 is limited to computation time of quick estimates). Estimation of
two-dimensional FP dynamics, but the method can model parameters and state may be achievable using
be extended to higher dimensions. unscented Kalman filtering as done in [25] for a spa-
The requirement to fit a model to patient data tially extended WC model. However the operational
to provide stimulation in phase space adds complex- practicality of phase space stimulation strategies
ity. To inform stimulation using the Hilbert trans- based on amplitude fields needs to be further
form without relying on models, data trajectories evaluated.
could directly be portrayed in the (y, H(y)) space,
where y is the centered tremor signal. However, 4.4. Future directions
in the case of ET, patients are asked to assume a Amplitude field based stimulation for neurological
tremor provoking posture to measure the response disorders could be investigated further. In PD, motor
to phase-locked DBS [10]. Because patients fatigue symptoms are correlated with increased subthalamic
quickly, only short recordings can be obtained (on nucleus (STN) beta band oscillatory power [68–72].
the order of 10 min [46]). In these short datasets, Excessive synchrony in PD has been explored with
the dependence of phase-locked DBS effects on the isostables in a model of coupled neurons [43], but
Hilbert phase of stimulation can be estimated in some not in a model constrained with experimental data.
patients [10, 32] (12 phase bins). The dependence of The WC model can describe the feedback loop com-
the effects of stimulation on the Hilbert amplitude posed of the STN and the globus pallidus pars
of the on-going tremor is more difficult to meas- externa [73–76], which plays a role in PD excess-
ure (three amplitude bins with large error bars in ive beta synchrony. Similarly to what was done in
[20]). An instantaneous amplitude response field in the present study, amplitude field based stimula-
the (y, H(y)) space detailed enough to optimize stim- tion could be applied to WC focus models fitted to
ulation is therefore out of reach. Moreover, providing PD patients, and compared to simpler stimulation
stimulation according to the Hilbert phase of tremor strategies. Moreover, because isostables are related
alone is unlikely to bring optimal benefits. to convergence properties of a system, isostable
An algorithm learning a target phase and a power amplitude could be provided as an additional fea-
threshold online to stimulate according to phase and ture to complement Hilbert amplitude when try-
power such as in [22] has many advantages, in par- ing to optimize stimulation with machine learning
ticular its simplicity and model-free character. How- approaches. Furthermore, isostable coordinates are at
ever, the response to stimulation is measured in [22] present limited to situations where a model is avail-
on a short timescale, and the subtle effects of phase- able. Obtaining the Koopman operator from data is
locked stimulation seen in patient data might there- being researched [77–79], which could make pos-
fore be overlooked. Moreover, stimulating accord- sible to recover isostables from data as there is a
ing to a target phase and a single power threshold strong connection between the Koopman operator
may be suboptimal compared to stimulation in phase and isostables [37]. A method to directly obtain
space based on amplitude fields, where the decision isostables from data has very recently been sugges-
to stimulate depends on the state, and predictions ted [80]. However it relies on limit cycle dynam-
about the future evolution of the state can be taken ics, and it is also unknown whether the technique
into account. How the strategies would compare in could be robust to the large levels of noise seen in
practice is however unclear, in particular in the light neurophysiological data. Finally, convergence time
of the greater complexity of phase space strategies information embedded in isostable amplitude fields
based on amplitude fields, which require model may prove more profitable to control more com-
fitting. plex, higher dimensional models, such as mean-field
Given model parameters, a corresponding aug- models of coupled populations of theta neurons [81,
mented instantaneous amplitude response field 82]. In higher dimensions, the computational advant-
(defined in section 3.3.1), and the state, our phase age of isostable amplitude fields compared to Hilbert
space stimulation strategy based on amplitude fields amplitude fields should also be larger.
15You can also read

















































