Obese dogs exhibit different fecal microbiome and specic microbial networks compared with normal weight dogs
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Obese dogs exhibit different fecal microbiome and
specific microbial networks compared with normal
weight dogs
Hanbeen Kim
Life and Industry Convergence Research Institute, Pusan National University
Jakyeom Seo
Life and Industry Convergence Research Institute, Pusan National University
Tansol Park
Chung-Ang University
Kangmin Seo
National Institute of Animal Science
Hyunwoo Cho
National Institute of Animal Science
Julan Chun
National Institute of Animal Science
Kihyun Kim ( kihyun@korea.kr )
National Institute of Animal Science
Article
Keywords: Body condition score, Obesity, Fecal microbiome, Specific network, Dog
Posted Date: June 6th, 2022
DOI: https://doi.org/10.21203/rs.3.rs-1616770/v2
License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Read Full License
Page 1/25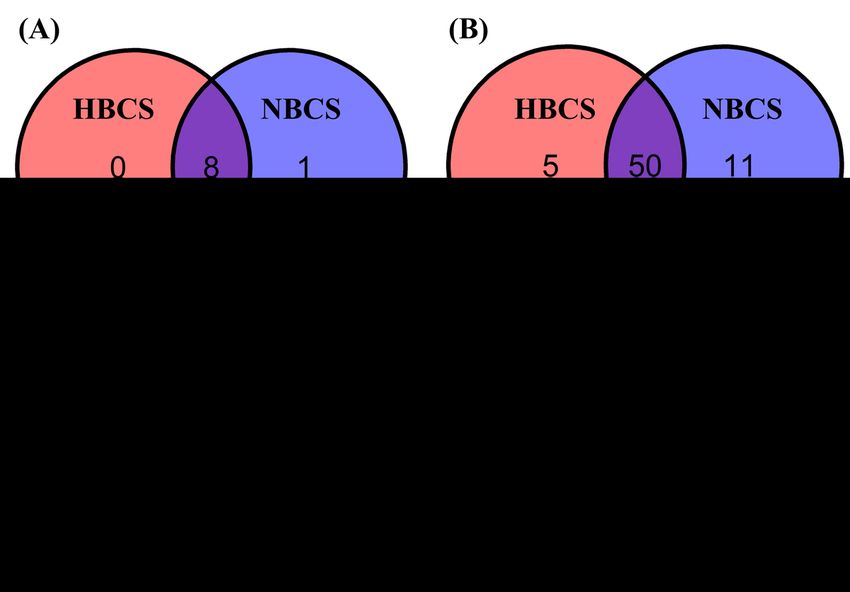
Abstract
Background
Canine obesity is a major health concern that predisposes dogs to various disorders. The fecal
microbiome has been attracting attention because of their impact on energy efficiency and metabolic
disorders of host. However, little is known about specific microbial interactions, and how these may be
affected by obesity in dogs. The objective of this study was to investigate the differences in fecal
microbiome and specific microbial networks between obese and normal dogs. A total of 20 beagle dogs
(males = 12, body weight [BW]: 10.5 ± 1.08 kg; females = 8, BW: 11.3 ± 1.71 kg; all 2-year-old) were fed to
meet the maintenance energy requirements for 18 weeks. Then, 12 beagle dogs were selected based on
body condition score (BCS) and divided into two groups: high BCS group (HBCS; BCS range: 7–9, males
= 4, females = 2) and normal BCS group (NBCS; BCS range: 4–6, males = 4, females = 2). In the final week
of the experiment, fecal samples were collected directly from the rectum, before breakfast, for analyzing
the fecal microbiome using 16S rRNA gene amplicon sequencing.
Results
The HBCS group had a significantly higher final BW than the NBCS group (P < 0.01). The relative
abundances of Faecalibacterium, Phascolarctobacterium, Megamonas, Bacteroides, Mucispirillum, and
an unclassified genus within Ruminococcaceae were significantly higher in the HBCS group than those in
the NBCS group (P < 0.05). Furthermore, some Kyoto Encyclopedia of Genes and Genomes (KEGG)
modules related to amino acid biosynthesis and B vitamins biosynthesis were enriched in the HBCS
group (P < 0.10), whereas those related to carbohydrate metabolism were enriched in the NBCS group (P <
0.10). Microbial network analysis revealed distinct co-occurrence and mutually exclusive interactions
between the HBCS and NBCS groups.
Conclusions
In conclusion, several genera related to short-chain fatty acid production were enriched in the HBCS
group. The enriched KEGG modules in the HBCS group enhanced energy efficiency through cross-feeding
between auxotrophs and prototrophs. However, further studies are needed to investigate how specific
networks can be interpreted in the context of fermentation characteristics in the lower gut and obesity in
dogs.
Background
Owing to the growing interest in dog welfare, more people have started to take an interest in the health
problems of dogs [1]. Among them, canine obesity is considered a major metabolic disease because it
can predispose dogs to a variety of disorders, such as cardiovascular disease [2], expiratory airway
Page 2/25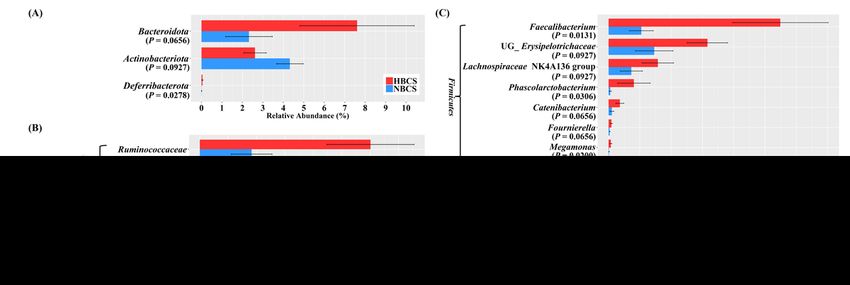
dysfunction [3], and diabetes mellitus [4], and has detrimental effects on longevity [5]. The main cause of
obesity in dogs is an imbalance between energy intake and expenditure [6]. Furthermore, some studies
suggest that breed, feed quality, and the owner’s interest in maintaining pet’s health are also possible
causes [7–9]. Therefore, the veterinary community is paying increasing attention to the prevention of
obesity in dogs.
Previously, companion dogs have been evaluated for being overweight and obese based on their body
weight (BW), body condition score (BCS), and body composition (fat mass and lean body mass) [10–12].
These methods have been recognized to be efficient in terms of cost and convenience. However, given the
limited data on the ideal weight for each breed and the subjective opinions of various veterinarians, these
methods need to be improved for better reliability [6]. Accordingly, recent advances in omics technology
are expected to replace traditional methods and provide new methods for investigating obesity indicators.
The gastrointestinal tract of dogs, especially the colon, is habituated by a complex microbiota that
contributes to energy source absorption, metabolic functions, and immunogenic responses [13]. Several
studies have investigated changes in the fecal microbiota in response to the modulation of dietary
components, including carbohydrates [14], proteins [15], and fats [16]. In addition, some studies have
reported differences in the fecal microbiota of obese dogs after a weight-loss program [17–20]. Taken
together, these studies highlight the importance of the fecal microbiota in sustaining canine health and
controlling obesity.
Previous studies have also reported differences in the gut microbiota among obese and lean individuals
in humans and mice [21–25]. These changes are characterized by a reduced diversity of gut microbiota
[21, 22], increased Firmicutes/Bacteroidota ratio [23, 24], and better energy harvesting [25] in the obese
group.
However, only a few studies have investigated the differences in the fecal microbiota between obese and
lean dogs [17, 20, 26]. The composition of the gut microbiota in dogs can differ based on several factors,
including diet, age, breed, and disorders [27–29]. Previous studies had several limitations in interpreting
the relationship between obesity and gut microbiota, including the lack of unification in terms of breeds
or ages [17, 20, 26]. Therefore, the present study investigated the effects of obesity on the gut microbiota
and microbial functional features of dogs of a similar age and breed. We hypothesized that fecal
microbiota, microbial functional profiles, and networks among microbiota would differ between obese
and normal dogs.
Therefore, this study aimed to investigate differences in fecal microbiota composition, microbial
functional profiles, and specific networks between obese and normal BCS groups after maintenance
energy requirements were met.
Methods
Animals and experimental design
Page 3/25This study was conducted using neutered beagles owned by the National Institute of Animal Science in
Korea. This experiment was conducted in accordance with the methods approved by the Animal Care and
Use Committee National Institute of Animal (NIAS-2019-370). All methods for animal care are performed
in accordance with the ARRIVE guidelines. Twenty beagle dogs (males = 12, initial body weight [BW]: 10.5
± 1.08 kg; females = 8, initial BW: 11.3 ± 1.71 kg; all 2-year-old) were enrolled in this study. The beagles
lived individually in an indoor space (1.8 m × 2.6 m) maintained at a temperature of 22–24 °C and
relative humidity from 60% to 80% using automatic temperature control and continuous ventilation. All
dogs were fed twice a day (at 10:00 and 16:00), had access to drinking water ad libitum, and engaged in
approximately 3 h of outdoor activity every day. The dogs were fed an extruded pellet-type commercial
dry food (Iskhan All-life33®; Wooriwa Ltd., Korea) that met the maintenance energy requirements
estimated based on the formula suggested by the Association of American Feed Control Officials
(AAFCO; metabolizable energy requirement = 132 kcal × kg BW0.75/day) [30] (Table 1). The experiment
was conducted for 18 weeks. In the first and final day of the experiment, the BCS was measured on a
nine-point scale according to the criteria of Laflamme D [12], and 12 neutered beagles (eight males and
four females, all 2-years-old) were selected based on the final BCS 24 h before collecting feces and
divided into two groups, each consisting of four males and two females. The normal BCS group (NBCS)
was composed of six healthy dogs with a normal BCS (4–6), and the high BCS group (HBCS) was
composed of six dogs with a high BCS (7–9) (Table 2).
Fecal sample collection and DNA extraction
Twenty-four hours before collecting feces, feeding was restricted. Fecal samples were collected directly
from the rectum before breakfast (9:00 to 10:00), immersed in liquid nitrogen, and stored in a deep freezer
at -80 °C until analysis. DNA was extracted from the fecal samples using the NucleoSpin DNA Stool kit
(Macherey-Nagel, Germany) according to the manufacturer’s instructions. Briefly, fecal samples (180–220
mg) were lysed using the MN Bead Tube Holder, and the genomic DNA was harvested through
NucleoSpinDNA stool column centrifugation at 13,000 × g for 1 min. In the final step, genomic DNA was
eluted into the elution buffer after centrifugation at 13,000 × g for 1 min and stored at -20 ℃.
16S rRNA gene amplicon sequencing and data processing
For PCR amplification targeting the V3 and V4 variable regions of 16S rRNA, the following primer
sets were used [31]: forward primer 5′-TCG TCG GCA GCG TCA GAT GTG TAT AAG AGA CAG CCT ACG
GGN GGC WGC AG-3′ and reverse primer 5′-GTC TCG TGG GCT CGG AGA TGT GTA TAA GAG ACA GGA
CTA CHV GGG TAT CTA ATC C-3′. Illumina adapter overhang nucleotide sequences were added to the
gene-specific sequences. The locus-specific sequences were as follows: forward overhang 5′-TCG TCG
GCA GCG TCA GAT GTG TAT AAG AGA CAG-3′ and reverse overhang 5′-GTC TCG TGG GCT CGG AGA TGT
GTA TAA GAG ACA G-3′. To purify the PCR products, KAPA HiFi HotStart ReadyMix (KAPA Biosystems,
USA) and Agencourt AMPure XP system (Beckman Coulter Genomics, USA) were used. The libraries were
sequenced using the Illumina MiSeq instrument (2 × 300 paired-end sequencing; Illumina, CA, USA).
Page 4/25The barcode sequences were trimmed using Cutadapt (version 1.11) [32]. Then, the raw amplicon
sequences were analyzed using built-in plugins within Quantitative Insights into Microbial Ecology 2
(QIIME2) (version 2019.04) [33]. First, the DADA2 plugin was used to remove primer sequences, denoise
low-quality reads (Q < 25), merge forward and reverse reads, and remove chimeric sequences. The raw
16S rRNA gene sequences are available in the National Center for Biotechnology Information sequence
read archive (BioProject accession: PRJNA818097). The amplicon sequence variants (ASVs) were
taxonomically classified using the Silva 16S rRNA gene database (version SSU138) [34]. Taxonomically
unassigned ASVs were excluded. The alpha diversity of each sample was examined with respect to ACE
(abundance-based coverage estimator) and Chao1 estimates, observed ASVs, evenness, Shannon
diversity index, and Simpson diversity index based on rarefied ASV tables using randomly selected 67,151
ASVs per sample. Principal coordinate analysis (PCoA) was conducted based on unweighted and
weighted UniFrac distance matrices to compare the overall dissimilarity of microbiota between the HBCS
and NBCS groups and visualized using Emperor implemented in QIIME2. The number of common and
exclusively identified bacterial taxa between the HBCS and NBCS groups at the phylum, family, and genus
levels were visualized using Venn diagrams. Functional metagenomic features from the 16S rRNA gene
sequences were predicted based on ASVs and the corresponding biological observation matrix (BIOM)
table using the Phylogenetic Investigation of Communities by Reconstruction of Unobserved State
(PICRUSt2) (version 2.3.0-b) with default options [35]. Principal component analysis (PCA) was
performed to compare the overall differences in predicted functional profiles based on the Kyoto
Encyclopedia of Genes and Genomes (KEGG) orthologs between the HBCS and NBCS groups. The PCA
plot was visualized using the ggfortify R package [36]. FastSpar was used to examine the correlations
among the major bacterial genera in the HBCS and NBCS groups [37], and the correlations were
computed based on the sparse correlations for compositional data [38]. Only significant microbial
interactions (P < 0.05) were further analyzed using co-expression differential network analysis
(CoDiNA) [39] to identify specific microbial networks in the HBCS and NBCS groups.
Statistical analysis
Before statistical analysis, all data were tested for normal distribution using the Kolmogorov–Smirnov
test in SAS 9.4 (SAS Institute Inc., NC, USA). Then, normally distributed data, including those on animal
performance (BW, BCS, and BW gain) and alpha diversity indices (ACE, Chao1, observed ASVs, evenness,
Shannon index, and Simpson diversity index) were analyzed using a t-test in SAS 9.4. Data that were not
normally distributed, including major microbial taxa and predicted microbial functional features from
KEGG categories and modules, were analyzed using a non-parametric Wilcoxon rank-sum test.
Permutational multivariate analysis of variance (PERMANOVA) was used to further analyze the PCoA and
PCA results to assess whether the overall microbial community and predicted functional profiles (KEGG
orthologs) differed significantly between the HBCS and NBCS groups. Statistical significance was
declared at P < 0.05, and a statistical trend was speculated at 0.05 ≤ P ≤ 0.10.
Results
Page 5/25Relation between BCS and animal performance
Both the initial BW and BCS were higher in the HBCS group than those in the NBCS group (Table 2). After
feeding to meet maintenance energy requirements for individual dogs, the HBCS group had a significantly
higher final BW and BCS than the NBCS group (P < 0.05), resulting in a higher BW gain in the HBCS group
(Table 2).
Quality assessment and sample statistics
A total of 1,826,502 sequences were obtained from 16S rRNA amplicon sequencing analysis, with an
average of 152,209 ± 30,573 sequences per fecal sample (Table S1). After quality filtering using QIIME 2
(Q score > 25), 1,193,079 sequences (65% of the raw reads) were generated, with an average of 99,423 ±
19,814 sequences per sample. Good’s coverage was greater than 99% for all fecal samples.
Relation between BCS and the alpha and beta diversity of fecal microbiota
In fecal microbiota, the difference in BCS did not result in significant changes in any of the alpha diversity
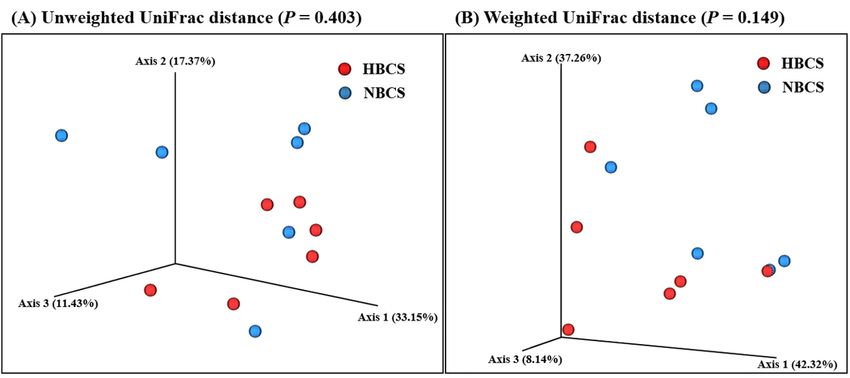
measurements (Table 3). The PCoA results based on both unweighted and weighted UniFrac
distances also did not present any significant difference in the fecal microbiota of the HBCS and NBCS
groups (Figure 1).
Relation between BCS and the fecal microbiota
In this study, only bacterial taxa have been presented because archaeal reads could not be detected. The
bacterial taxa that were detected in over 50% of the samples and had a relative abundance of over 0.05%
in at least one of the BCS groups were defined as major microbial taxa.
The Venn diagram shows common and exclusively identified prokaryotic taxa in the HBCS and NBCS
groups (Figure 2). Eight of the nine phyla were shared between the HBCS and NBCS groups, and only
Desulfobacterota was exclusively detected in the NBCS group (Figure 2A); moreover, Desulfobacterota
was detected in only one of the six samples. At the family level, 50 families (approximately 75.8%) were
shared between the HBCS and NBCS groups and no major families were found exclusively in the BCS
group (Figure 2B). At the genus level, more than 100 genera (approximately 72.7%) were shared between
the HBCS and NBCS groups, and only 14 and 24 genera were exclusively detected in the HBCS and NBCS
groups, respectively (Figure 2C). Succinivibrio and an unclassified genus within Atopobiaceae were the
only major genera (present in at least 50% of the samples in each group) found exclusively in the HBCS
and NBCS groups, respectively; however, they only accounted for a minor proportion (relative abundance
< 0.01%).
The major phyla, families, and genera with relative abundances of over 0.5% in at least one group are
shown in Figure S1. Among them, the major taxa that showed statistical tendency (P < 0.10) are shown in
Figure 3 (relative abundance ≥ 0.05% in at least one group). Firmicutes was the most predominant
phylum (accounting for at least 77% of the total microbiota), followed by Fusobacteriota (Figure S1A). Of
Page 6/25the five predominant phyla, the relative abundance of Bacteroidota tended to be higher in the HBCS group
than that in the NBCS group (HBCS, 7.61% vs. NBCS, 2.32%; P = 0.0656), whereas Actinobacteria showed
an opposite trend (HBCS, 2.62% vs. NBCS, 4.32%; P = 0.0927) (Figure 3A). The relative abundance of
Deferribacterota, accounting for a minor proportion, was significantly higher in the HBCS group than that
in the NBCS group (HBCS, 0.051% vs. NBCS, 0.008%; P = 0.0278) (Figure 3A). At the family level,
Lachnospiraceae, Peptostreptococcaceae, and Lactobacillaceae were the three most abundant families
in the HBCS group, whereas Peptostreptococcaceae, Erysipelotrichaceae, and Lactobacillaceae were the
most abundant in the NBCS group (Figure S1B). Among the major classified families, the relative
abundances of Ruminococcaceae, Bacteroidaceae, Acidaminococcaceae, Selenomonadaceae, and
Deferribacteraceae were higher in the HBCS group than those in the NBCS group (Figure 3B). At the genus
level, Peptoclostridium was the most predominant genus in both BCS groups (HBCS, 14.90% vs. NBCS,
23.79%), and more than 84% of the entire fecal microbiota was assigned to 54 major classified genera
(representing > 0.05% of relative abundance in at least one BCS group). At the genus level, the relative
abundances of Bacteroides, Mucispirillum, and the four genera within Firmicutes (Faecalibacterium,
Phascolarctobacterium, Megamonas, and UG_Ruminococcaceae) were significantly higher in the HBCS
group than those in the NBCS group (Figure 3C, P < 0.05). Meanwhile, the relative abundances of
Sutterella, Prevotellaceae Ga6A1 group, and four genera within Firmicutes (UG_Erysipelotrichaceae,
Lachnospiraceae NK4A136 group, Catenibacterium, and Fournierella) tended to be higher in the HBCS
group than those in the NBCS group (Figure 3C, P < 0.10).
Relation between BCS and the predicted functional profiles from fecal microbiota
The overall distribution of the microbial functional profiles predicted based on KEGG orthologs was
visualized using PCA plots (Figure 4A). There was no significant difference in the overall distribution of
KEGG orthologs between the HBCS and NBCS groups based on the PERMANOVA analysis. Among the
top 10 KEGG categories (Level 2), only carbohydrate metabolism was significantly higher in the NBCS
group than that in the HBCS group (Figure 4B, P < 0.05). Among the major KEGG modules, three modules
related to amino acid metabolism (lysine biosynthesis [M00030], serine biosynthesis [M00020], and
tryptophan biosynthesis [M00023]) and five modules related to the metabolism of cofactors and vitamins
(cobalamin biosynthesis [M00122], pyridoxal-P biosynthesis [M00124], phylloquinone biosynthesis
[M00932], and lipoic acid biosynthesis [M00881, M00884]) were markedly enriched in the HBCS group
(Figure 4B, P < 0.10). In contrast, three modules related to carbohydrate metabolism (galactose
degradation [M00632], nucleotide sugar biosynthesis [M00549], and D-galactonate degradation
[M00552]), two modules related to energy metabolism (reductive pentose phosphate cycle [M00165] and
methanogenesis [M00357]), and one related to cysteine biosynthesis [M00609] tended to be higher in the
NBCS group (Figure 4C, P < 0.10).
Differential network analysis based on BCS
The statistics for the differential networks of the major genera in the HBCS and NBCS groups are listed in
Table 4. The total number of nodes for the HBCS and NBCS groups was 31 and 39, respectively, and the
Page 7/25total number of edges was 74 and 71, respectively. In the differential network of the HBCS group, positive
edges were detected more frequently than negative edges (Table 4; 51 positive edges vs. 23 negative
edges), whereas the number of positive edges was similar to that of negative edges in the differential
network of the NBCS group (Table 4; 36 positive edges vs. 35 negative edges). In the differential networks
of each treatment, the γ link, which was specific to one of the networks, was assigned to 29% of the HBCS
group and 23.1% of the NBCS group (Table 4). The specific genera (γ link) in the differential network of
the HBCS group were Allobaculum, Lachnospiraceae NK4A136 group, UG_Ruminococcaceae, Collinsella,
UG_Erysipelotrichaceae, Phascolarctobacterium, Muribaculaceae, Megamonas, and Fusobacterium
(Figure 5A), and those of NBCS group were Veillonella, Prevotella, Negativibacillus, Bifidobacterium,
Bacillus, Oribacterium, Turicibacter, Faecalibacterium, and Helicobacter (Figure 5B). Based on the two
centrality measurements (degree and eigenvector centrality) and hub score within each differential
network, UG_Erysipelotrichaceae and Phascolarctobacterium were the keystone genera in HBCS, while
Negativibacillus and Bifidobacterium were those in the NBCS group (Figure 5 and Figure S2).
Discussion
Several human obesity studies have reported that obesity is generally linked to altered gut microbiota,
presenting lower diversity in obese humans than that in lean humans [21, 22]. In addition, a recent
metagenomic study revealed that the dog microbiota was similar to human microbiota in comparison
with pig or mouse microbiota, and the alteration of microbiota in response to dietary changes was also in
agreement with that observed in human studies [40]. Therefore, we expected the HBCS group to have a
lower microbiota diversity than the NBCS group. However, the present study revealed no significant
differences in the alpha and beta diversity measurements between the HBCS and NBCS groups. Similar
to the results of microbial diversity in this study, Handl S, German AJ, Holden SL, Dowd SE, Steiner JM,
Heilmann RM, Grant RW, Swanson KS and Suchodolski JS [17] reported no significant differences in
alpha diversity (Shannon’s index, evenness, and the number of operational taxonomic units, i.e. OTUs)
and beta diversity between lean and obese companion dogs. In addition, a recent study that evaluated
differences in the fecal microbiota among obese, lean, and weight loss dogs showed that there were no
significant differences in alpha diversity (Faith’s phylogenetic diversity, Shannon’s index, and the number
of OTUs) and beta diversity, whereas the evenness index was significantly higher in the obese group than
that in the lean and weight loss groups [20].
Previous studies have reported that the five predominant phyla in the fecal microbiota of dogs are
Firmicutes, Bacteroidota, Proteobacteria, Actinobacteriota, and Fusobacterium [27, 40] as shown in the
present study. The Firmicutes/Bacteroidota ratio is generally considered an indicator of obesity because
several studies in humans and mice have shown that the obese group has a higher
Firmicutes/Bacteroidota ratio than the lean group [23, 24]. In addition, a previous study proposed that
Firmicutes was more effective at harvesting energy from a diet than Bacteroidetes; therefore, a higher
Firmicutes/Bacteroidota ratio may promote weight gain [41]. However, in the present study, the relative
abundance of Bacteroidota tended to be higher in the HBCS group, whereas there was no significant
difference in the relative abundance of Firmicutes, thereby reducing the Firmicutes/Bacteroidota ratio in
Page 8/25the HBCS group (data not shown). Meanwhile, some studies on gut microbiota composition in obese
humans or mice have also reported conflicting results, such as a lower Firmicutes/Bacteroidota ratio [42]
or no difference in the Firmicutes/Bacteroidota ratio [43]. In addition, several studies have observed no
significant differences in both Firmicutes and Bacteroidota between obese and lean dogs [15, 17, 44].
Park HJ, Lee SE, Kim HB, Isaacson R, Seo KW and Song KH [26] reported that the relative abundance of
Firmicutes was significantly lower in the obese group than that in the lean group, but no difference was
observed in Bacteroidota. In a recent study, Macedo HT, Rentas MF, Vendramini THA, Macegoza MV,
Amaral AR, Jeremias JT, de Carvalho Balieiro JC, Pfrimer K, Ferriolli E and Pontieri CFF [20] showed a
significantly higher proportion of Bacteroidota and a lower proportion of Firmicutes in obese dogs than
that in lean dogs, which is similar to our findings. Therefore, a high relative abundance of Firmicutes, low
relative abundance of Bacteroidota, and a high Firmicutes/Bacteroidota ratio might not be reliable
biomarkers to identify obese dogs.
Butyrate is a major short-chain fatty acid in the gut environment because it is the main fuel for epithelial
cells in the colon and exerts anti-inflammatory and anti-carcinogenic effects [45]; therefore, butyrate-
producing bacteria are considered important microbiota [46]. In this study, Faecalibacterium and
Lachnospiraceae NK4A136 group were enriched in the HBCS group. Faecalibacterium is the predominant
genus belonging to Clostridial cluster IV and can produce butyrate and D-lactate by degrading a wide
range of carbohydrate substrates [47]. Previous studies have reported conflicting results regarding the
relative abundance of Faecalibacterium between obese and lean humans, such as a lower [48, 49] or
higher [50–52] abundance in obese humans. In the fecal microbiome of dogs, there are few reports on the
relationship between members of Faecalibacterium and obesity. Recently, Macedo HT, Rentas MF,
Vendramini THA, Macegoza MV, Amaral AR, Jeremias JT, de Carvalho Balieiro JC, Pfrimer K, Ferriolli E
and Pontieri CFF [20] reported that the relative abundance of Faecalibacterium was significantly higher in
obese dogs than that in lean dogs, and it decreased significantly after the obese group underwent a
weight loss program. Thus, further studies are needed to confirm the role of Faecalibacterium in dogs
with obesity. Lachnospiraceae NK4A136 group is a potential butyrate producer [53], and Companys J,
Gosalbes MJ, Pla-Pagà L, Calderón-Pérez L, Llauradó E, Pedret A, Valls RM, Jiménez-Hernández N,
Sandoval-Ramirez BA and Del Bas JM [52] previously reported that the relative abundance of the
Lachnospiraceae NK4A136 group was enriched in obese humans, which is supported by our results.
Hence, the presence of butyrate-producing bacteria in the fecal microbiome of dogs may be related to
obesity. The abundance of the family Ruminococcaceae was higher in the HBCS group. Many genera
under Ruminococcaceae belong to cluster IV and possess enzymes that degrade recalcitrant
carbohydrate sources and produce acetate as a major fermentation product that can be used as an
energy source by the host [47]. In addition, four major genera related to acetate production as the major
fermentation product, specifically Fournierella [54], Catenibacterium [55], Megamonas [56], and
unclassified genera within Ruminococcaceae [47], were enriched in the HBCS group. Faecalibacterium
prausnitzii, which is a major species belonging to Faecalibacterium, requires acetate for growth [57].
Therefore, the increase in Faecalibacterium may be linked to the high abundance of acetate-producing
bacteria. Furthermore, several studies have suggested a relationship between Sutterella spp. and obesity
Page 9/25in humans [58, 59]. A previous study reported a higher relative abundance of Sutterella in obese dogs [20],
which is supported by our results. Thus, we speculate that the increase in Sutterella may be related to
obesity in dogs. Serena C, Ceperuelo-Mallafré V, Keiran N, Queipo-Ortuño MI, Bernal R, Gomez-Huelgas R,
Urpi-Sarda M, Sabater M, Pérez-Brocal V and Andrés-Lacueva C [60] revealed that obesity in humans is
associated with elevated levels of circulating succinate concomitant with specific changes in succinate-
related microbiota, especially higher succinate producers (Prevotellaceae and Veillonellaceae) and lower
succinate consumers (Odoribacteraceae and Clostridiaceae). In the present study, there were no
significant differences in these families between the HBCS and NBCS groups. Instead, we found that the
relative abundances of Bacteroides [61, 62] and Mucispirillum [63], which are considered to be succinate
producers [63], and that of Phascolarctobacterium, which is a propionate producer through the succinate
pathway, were higher in the HBCS group at the genus level. We thus suggest that obese dogs may have a
higher relative abundance of succinate producers and utilizers than lean dogs.
The alteration in fecal microbiota composition is linked to changes in microbial functional features. In
this study, the difference in BCS did not affect the overall predicted functional features, as expected from
the lack of differences in the overall bacterial diversity (Table 3). An excessive energy supply to the host
by increasing carbohydrate metabolism is an important factor in obese animals [64]. In addition, several
studies have reported an increase in the functional profile of carbohydrate metabolism in obese humans
[65, 66]. Therefore, we expected an increased relative abundance of functional profiles of carbohydrate
metabolism in the HBCS group. However, carbohydrate metabolism among the top 10 KEGG categories
showed a significant increase only in the NBCS group. Some KEGG modules involved in carbohydrate
metabolism (galactose degradation [M00632], nucleotide sugar biosynthesis [M00549], and D-
galactonate degradation [M00552]) were significantly enriched in the NBCS group. The unexpected
increase in carbohydrate metabolism can be explained by two possible reasons: 1) the functional profile
was predicted based on DNA; therefore, the RNA-based results (i.e.. RNA sequencing) in carbohydrate
metabolism may be different from this result, and 2) the lack of a dog microbiome-specific PICRUSt2
database. In this study, some KEGG modules related to B vitamins biosynthesis (cobalamin biosynthesis
[M00122] and pyridoxal-P biosynthesis [M00124]) and amino acid biosynthesis (lysine biosynthesis
[M00030], serine biosynthesis [M00020], and tryptophan biosynthesis [M00023]) were enriched in the
HBCS group. The microbiota produces many types of micronutrients that play an important role in host
homeostasis [67]. B vitamins acts as an enzymatic cofactor or a precursor of cofactors [68] and can be
exchanged and shared between microbiota, thereby stabilizing the gut microbiota by cross-feeding [69]. A
recent study that investigated the effects of co-culturing butyrate producers revealed that
Faecalibacterium prausnitzii, a major species belonging to Faecalibacterium, requires several B vitamins
and tryptophan for growth (auxotrophs) [70]. They also demonstrated that co-culturing Faecalibacterium
prausnitzii with species belonging to Lachnospiraceae, which are prototrophic (capable of de novo
synthesis) for all amino acids and several vitamins, stimulates the growth of Faecalibacterium prausnitzii
through cross-feeding [70]. In this study, the HBCS group showed a higher relative abundance of butyrate
producers (Faecalibacterium and Lachnospiraceae NK4A136 group) and obesity-related genera
(Lachnospiraceae NK4A136 group and Sutterella). Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V,
Page 10/25Mardis ER and Gordon JI [25] previously suggested that the microbiota of obese mice may be more
efficient in energy harvesting than that of lean mice. Therefore, we speculate that obesity in dogs may be
influenced by the enhancement of energy-harvesting efficiency through cross-feeding among fecal
microbiota, especially among auxotrophs and prototrophs.
We also compared the overall interactions within the major fecal bacterial genera belonging to each BCS
group. Unclassified genera within Erysipelotrichaceae and Phascolarctobacterium were the keystone
genera within exclusive networks in the HBCS group. Previous studies have reported that several species
belonging to the family Erysipelotrichaceae are associated with obesity in mouse models [71] and
humans [72]. Additionally, an association between Erysipelotrichaceae and host lipid metabolism has
been suggested [73]. In the present study, UG_Erysipelotrichaceae had an exclusive positive interaction
with five major genera, namely Allobaculum, Fusobacterium, Lachnospiraceae NK4A136 group,
Megamonas, and Phascolarctobacterium. Among these genera, Lachnospiraceae NK4A136 group [52]
and Megamonas [74], which were higher in abundance in the HBCS group, were also positively
associated with obesity and weight gain, respectively. Phascolarctobacterium consumes succinate and
produces propionate during carbohydrate fermentation [75]. Succinate is not only a metabolic end-
product of some bacteria but also a primary cross-feeding metabolite in microbial propionate synthesis
[76]. Phascolarctobacterium showed a strong positive correlation with 10 major genera, whereof three
genera, namely Fusobacterium [77], Bacteroides [61, 62], and Mucispirillum [63] are considered to be
succinate producers. Interestingly, Muribaculaceae, which showed no difference in the relative abundance
between the HBCS and NBCS groups, exhibited exclusively negative correlations with four major genera,
namely the Lachnospiraceae NK4A136 group, Megamonas, Phascolarctobacterium, and Prevotellaceae
Ga6A1 group, which had significantly higher abundances in the HBCS group. Although little is known
about the relationship between Muribaculaceae and obesity, a previous study revealed the resistance
effect of Muribaculaceae in lean mice fed with a high-fat diet [78]. Therefore, we speculate that
Muribaculaceae may play an anti-obesity role in dogs because of its negative correlation with obesity-
related genera. In the exclusive network of the NBCS group, Bifidobacterium and Negativibacillus were the
keystone genera. Bifidobacterium is well known as an important probiotic that exerts beneficial health
effects, such as regulating microbial homeostasis, reducing intestinal lipopolysaccharide levels, and
improving the mucosal barrier function [79–82]. In addition, several studies have reported that the relative
abundance of Bifidobacterium is lower in mice fed a high-fat diet [83, 84]. In this study, Bifidobacterium
exhibited a strong negative correlation with Peptoclostridium and Blautia, which have previously been
associated with obesity in dogs [20, 85], and a strong positive correlation with Lactobacillus, Sellimonas,
and Negativibacillus. A previous study suggested that Lactobacillus may serve as a potential probiotic
for dogs [27] and Sellimonas plays an important role in gut recovery homeostasis [86]. Therefore, the co-
occurrence interactions among potential probiotics and mutually exclusive interactions among obesity-
related genera might have maintained the BW of the NBCS group.
The majority of previous studies investigating the fecal microbiome in obese dogs focused on the impact
of dietary changes or physical training to reduce weight rather than on natural differences in the
microbiome and microbial interactions. In this study, we investigated essential differences in the fecal
Page 11/25microbiome and their interactions between normal and obese dogs. However, we found several results,
especially regarding microbiota related to succinate metabolism and functional profiles of carbohydrate
metabolism, that were inconsistent with the results of previous studies on obese mice and humans.
Therefore, to better understand obesity in dogs, future studies should be conducted to further investigate
the differences in microbial functional metabolism and metabolites between normal and obese dogs.
Conclusions
Differences in BCS did not affect the overall distribution of the microbiota and predicted functional
features. However, several genera related to short-chain fatty acid production were enriched in the HBCS
group. In the predicted microbial functional profiles, some KEGG modules related to B vitamins and
amino acid biosyntheses were higher in the HBCS group, indicating enhanced energy efficiency through
cross-feeding between auxotrophs and prototrophs. In addition, specific co-occurrence and mutually
exclusive interactions among the different genera were detected among the HBCS and NBCS groups.
Further studies are needed to investigate how the specific networks can be interpreted in the context of
fecal fermentation characteristics and obesity in dogs.
Abbreviations
BW
body weight
BCS
body condition score
KEGG
kyoto encyclopedia of genes and genomes
AAFCO
association of american feed control officials
QIIME2
quantitative insights into microbial ecology 2
ASV
amplicon sequence variant
ACE
abundance-based coverage estimator
PCoA
principal coordinate analysis
BIOM
biological observation matrix
PICRUSt2
phylogenetic investigation of communities by reconstruction of unobserved state
PCA
Page 12/25principal component analysis
CoDiNA
co-expression differential network analysis
PERMANOVA
permutational multivariate analysis of variance
OTU
operational taxonomic unit
Declarations
Ethics approval and consent to participate
The animal study was reviewed and approved by the National Institute of Animal Science in Korea. This
experiment was conducted in accordance with the methods approved by the Animal Care and Use
Committee National Institute of Animal (NIAS-2019-370).
Consent for publication
Not applicable.
Availability of data and materials
The raw 16S rRNA sequences have been deposited in the NCBI SRA ubder BioProject PRJNA818097. All
analyzed microbioal datasets in the present study are available from the corresponding author on
reasonable request.
Competing interests
The authors declare that they have no competing interests.
Funding
This work was carried out with the support of the Cooperative Research Program of Center for
Companion Animal Research (Project No. PJ013984012022), Rural Development Administration, Korea.
This study was supported by the 2022 RDA Fellowship Program of the National Institute of Animal
Science, Rural Development Administration, Korea.
Authors' contributions
The study design was performed by JS and KK. KS, HC, and JC conducted the experiment and analyzed
the animal data. HK and JS did the sequencing based analysis. All the visualization of data and
statistical analyses were performed by HK. HK and JK drafted the manuscript. KK reviewed and edited the
manuscript. All authors read and approved the final manuscript.
Page 13/25Acknowledgements
Not applicable
References
1. Houpt KA, Goodwin D, Uchida Y, Baranyiová E, Fatjó J, Kakuma Y: Proceedings of a workshop to
identify dog welfare issues in the US, Japan, Czech Republic, Spain and the UK. Appl Anim Behav Sci
2007, 106:221–233.
2. Mizelle HL, Edwards TC, Montant J-P: Abnormal cardiovascular responses to exercise during the
development of obesity in dogs. Am J Hypertens 1994, 7:374–378.
3. Bach JF, Rozanski EA, Bedenice D, Chan DL, Freeman LM, Lofgren JL, Oura TJ, Hoffman AM:
Association of expiratory airway dysfunction with marked obesity in healthy adult dogs. Am J Vet
Res 2007, 68:670–675.
4. Mattheeuws D, Rottiers R, Kaneko J, Vermeulen A: Diabetes mellitus in dogs: relationship of obesity
to glucose tolerance and insulin response. Am J Vet Res 1984, 45:98–103.
5. Kealy RD, Lawler DF, Ballam JM, Mantz SL, Biery DN, Greeley EH, Lust G, Segre M, Smith GK, Stowe
HD: Effects of diet restriction on life span and age-related changes in dogs. J Am Vet Med Assoc
2002, 220:1315–1320.
6. German AJ: The growing problem of obesity in dogs and cats. J Nutr 2006, 136:1940S-1946S.
7. Edney A, Smith P: Study of obesity in dogs visiting veterinary practices in the United Kingdom. Vet
Rec 1986, 118:391–396.
8. Mason E: Obesity to pet dogs. Vet Rec 1970, 86:612–616.
9. Kienzle E, Bergler R, Mandernach A: A comparison of the feeding behavior and the human–animal
relationship in owners of normal and obese dogs. J Nutr 1998, 128:2779S-2782S.
10. Burkholder WJ: Precision and practicality of methods assessing body composition of dogs and cats.
Compend Contin Educ Pract Vet 2001, 23:1–10.
11. Pendergrass PB, Bartley CM, Nagy F, Ream LJ, Stuhlman R: A rapid method for determining normal
weights of medium-to‐large mongrel dogs. J Small Anim Pract 1983, 24:269–276.
12. Laflamme D: Development and validation of a body condition score system for dogs. Canine Pract
(Santa Barbara, Calif: 1990)(USA) 1997.
13. Huang Z, Pan Z, Yang R, Bi Y, Xiong X: The canine gastrointestinal microbiota: early studies and
research frontiers. Gut Microbes 2020, 11:635–654.
14. Li Q, Lauber CL, Czarnecki-Maulden G, Pan Y, Hannah SS: Effects of the dietary protein and
carbohydrate ratio on gut microbiomes in dogs of different body conditions. mBio 2017, 8:e01703-
01716.
15. Xu J, Verbrugghe A, Lourenço M, Cools A, Liu DJ, Van de Wiele T, Marzorati M, Eeckhaut V, Van
Immerseel F, Vanhaecke L: The response of canine faecal microbiota to increased dietary protein is
Page 14/25influenced by body condition. BMC Vet Res 2017, 13:1–11.
16. Moinard A, Payen C, Ouguerram K, André A, Hernandez J, Drut A, Biourge VC, Suchodolski JS,
Flanagan J, Nguyen P: Effects of high-fat diet at two energetic levels on fecal microbiota, colonic
barrier, and metabolic parameters in dogs. Front Vet Sci 2020, 7:699.
17. Handl S, German AJ, Holden SL, Dowd SE, Steiner JM, Heilmann RM, Grant RW, Swanson KS,
Suchodolski JS: Faecal microbiota in lean and obese dogs. FEMS Microbiol Ecol 2013, 84:332–343.
18. Salas-Mani A, Jeusette I, Castillo I, Manuelian CL, Lionnet C, Iraculis N, Sanchez N, Fernández S,
Vilaseca L, Torre C: Fecal microbiota composition changes after a BW loss diet in Beagle dogs. J
Anim Sci 2018, 96:3102–3111.
19. Sanchez SB, Pilla R, Sarawichitr B, Gramenzi A, Marsilio F, Steiner JM, Lidbury JA, Woods GR,
German AJ, Suchodolski JS: Fecal microbiota in client-owned obese dogs changes after weight loss
with a high-fiber-high-protein diet. PeerJ 2020, 8:e9706.
20. Macedo HT, Rentas MF, Vendramini THA, Macegoza MV, Amaral AR, Jeremias JT, de Carvalho
Balieiro JC, Pfrimer K, Ferriolli E, Pontieri CFF: Weight-loss in obese dogs promotes important shifts
in fecal microbiota profile to the extent of resembling microbiota of lean dogs. Anim Microbiome
2022, 4:1–13.
21. Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, Sogin ML, Jones WJ, Roe
BA, Affourtit JP: A core gut microbiome in obese and lean twins. Nat 2009, 457:480–484.
22. Le Chatelier E, Nielsen T, Qin J, Prifti E, Hildebrand F, Falony G, Almeida M, Arumugam M, Batto J-M,
Kennedy S: Richness of human gut microbiome correlates with metabolic markers. Nat 2013,
500:541–546.
23. Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI: Obesity alters gut microbial
ecology. Proc Natl Acad Sci 2005, 102:11070–11075.
24. Ley RE, Turnbaugh PJ, Klein S, Gordon JI: Human gut microbes associated with obesity. Nat 2006,
444:1022–1023.
25. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI: An obesity-associated gut
microbiome with increased capacity for energy harvest. Nat 2006, 444:1027–1031.
26. Park HJ, Lee SE, Kim HB, Isaacson R, Seo KW, Song KH: Association of obesity with serum leptin,
adiponectin, and serotonin and gut microflora in beagle dogs. J Vet Intern Med 2015, 29:43–50.
27. Grześkowiak Ł, Endo A, Beasley S, Salminen S: Microbiota and probiotics in canine and feline
welfare. Anaerobe 2015, 34:14–23.
28. Lee W-J, Hase K: Gut microbiota–generated metabolites in animal health and disease. Nat Chem Biol
2014, 10:416–424.
29. Simpson J, Martineau B, Jones W, Ballam J, Mackie RI: Characterization of fecal bacterial
populations in canines: effects of age, breed and dietary fiber. Microb Ecol 2002, 44:186–197.
30. AAFCO: Association of American Feed Control Officials. The Official Publication of the Association of
American Feed Control Officials 2019.
Page 15/2531. Klindworth A, Pruesse E, Schweer T, Peplies J, Quast C, Horn M, Glöckner FO: Evaluation of general
16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity
studies. Nucleic Acids Res 2013, 41:e1-e1.
32. Martin M: Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J
2011, 17:10–12.
33. Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, Al-Ghalith GA, Alexander H, Alm EJ,
Arumugam M, Asnicar F: Reproducible, interactive, scalable and extensible microbiome data science
using QIIME 2. Nat Biotechnol 2019, 37:852–857.
34. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO: The SILVA
ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids
Res 2012, 41:D590-D596.
35. Douglas GM, Maffei VJ, Zaneveld JR, Yurgel SN, Brown JR, Taylor CM, Huttenhower C, Langille MG:
PICRUSt2 for prediction of metagenome functions. Nat Biotechnol 2020, 38:685–688.
36. Tang Y, Horikoshi M, Li W: ggfortify: unified interface to visualize statistical results of popular R
packages. R J 2016, 8:474.
37. Watts SC, Ritchie SC, Inouye M, Holt KE: FastSpar: rapid and scalable correlation estimation for
compositional data. Bioinform 2019, 35:1064–1066.
38. Friedman J, Alm EJ: Inferring correlation networks from genomic survey data. 2012.
39. Morselli Gysi D, de Miranda Fragoso T, Zebardast F, Bertoli W, Busskamp V, Almaas E, Nowick K:
Whole transcriptomic network analysis using co-expression differential network analysis (CoDiNA).
PLoS One 2020, 15:e0240523.
40. Coelho LP, Kultima JR, Costea PI, Fournier C, Pan Y, Czarnecki-Maulden G, Hayward MR, Forslund SK,
Schmidt TSB, Descombes P: Similarity of the dog and human gut microbiomes in gene content and
response to diet. Microbiome 2018, 6:1–11.
41. Krajmalnik-Brown R, Ilhan ZE, Kang DW, DiBaise JK: Effects of gut microbes on nutrient absorption
and energy regulation. Nutr Clin Pract 2012, 27:201–214.
42. Schwiertz A, Taras D, Schäfer K, Beijer S, Bos NA, Donus C, Hardt PD: Microbiota and SCFA in lean
and overweight healthy subjects. Obe 2010, 18:190–195.
43. Duncan SH, Lobley G, Holtrop G, Ince J, Johnstone A, Louis P, Flint HJ: Human colonic microbiota
associated with diet, obesity and weight loss. Int J Obe 2008, 32:1720–1724.
44. Chun JL, Ji SY, Lee SD, Lee YK, Kim B, Kim KH: Difference of gut microbiota composition based on
the body condition scores in dogs. J Anim Sci Technol 2020, 62:239.
45. Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost F, Brummer RJ: The role of butyrate on
colonic function. Aliment Pharmacol Ther 2008, 27:104–119.
46. Louis P, Flint HJ: Diversity, metabolism and microbial ecology of butyrate-producing bacteria from
the human large intestine. FEMS Microbiol Lett 2009, 294:1–8.
Page 16/2547. Duncan SH, Louis P, Flint HJ: Cultivable bacterial diversity from the human colon. Lett Appl Microbiol
2007, 44:343–350.
48. Yasir M, Angelakis E, Bibi F, Azhar EI, Bachar D, Lagier JC, Gaborit B, Hassan AM, Jiman-Fatani AA,
Alshali KZ, et al: Comparison of the gut microbiota of people in France and Saudi Arabia. Nutr
Diabetes 2015, 5:e153-e153.
49. Andoh A, Nishida A, Takahashi K, Inatomi O, Imaeda H, Bamba S, Kito K, Sugimoto M, Kobayashi T:
Comparison of the gut microbial community between obese and lean peoples using 16S gene
sequencing in a Japanese population. J Clin Biochem Nutr 2016, advpub.
50. Chávez-Carbajal A, Nirmalkar K, Pérez-Lizaur A, Hernández-Quiroz F, Ramírez-del-Alto S, García-Mena
J, Hernández-Guerrero C: Gut Microbiota and Predicted Metabolic Pathways in a Sample of Mexican
Women Affected by Obesity and Obesity Plus Metabolic Syndrome. Int J Mol Sci 2019, 20:438.
51. Salah M, Azab M, Ramadan A, Hanora A: New Insights on Obesity and Diabetes from Gut
Microbiome Alterations in Egyptian Adults. Omics 2019, 23:477–485.
52. Companys J, Gosalbes MJ, Pla-Pagà L, Calderón-Pérez L, Llauradó E, Pedret A, Valls RM, Jiménez-
Hernández N, Sandoval-Ramirez BA, Del Bas JM: Gut microbiota profile and its association with
clinical variables and dietary intake in overweight/obese and lean subjects: A cross-sectional study.
Nutrients 2021, 13:2032.
53. Stadlbauer V, Engertsberger L, Komarova I, Feldbacher N, Leber B, Pichler G, Fink N, Scarpatetti M,
Schippinger W, Schmidt R, Horvath A: Dysbiosis, gut barrier dysfunction and inflammation in
dementia: a pilot study. BMC Geriatr 2020, 20:248.
54. Togo AH, Durand G, Khelaifia S, Armstrong N, Robert C, Cadoret F, Di Pinto F, Delerce J, Levasseur A,
Raoult D: Fournierella massiliensis gen. nov., sp. nov., a new human-associated member of the
family Ruminococcaceae. Int J Syst Evol Microbiol 2017, 67:1393–1399.
55. Kageyama A, Benno Y: Catenibacterium mitsuokai gen. nov., sp. nov., a gram-positive anaerobic
bacterium isolated from human faeces. Int J Syst Evol Microbiol 2000, 50:1595–1599.
56. Chevrot R, Carlotti A, Sopena V, Marchand P, Rosenfeld E: Megamonas rupellensis sp. nov., an
anaerobe isolated from the caecum of a duck. Int J Syst Evol Microbiol 2008, 58:2921–2924.
57. Duncan SH, Hold GL, Harmsen HJ, Stewart CS, Flint HJ: Growth requirements and fermentation
products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii
gen. nov., comb. nov. Int J Syst Evol Microbiol 2002, 52:2141–2146.
58. Hou Y-P, He Q-Q, Ouyang H-M, Peng H-S, Wang Q, Li J, Lv X-F, Zheng Y-N, Li S-C, Liu H-L: Human gut
microbiota associated with obesity in Chinese children and adolescents. BioMed Res Int 2017, 2017.
59. Gao X, Zhang M, Xue J, Huang J, Zhuang R, Zhou X, Zhang H, Fu Q, Hao Y: Body mass index
differences in the gut microbiota are gender specific. Front Microbiol 2018, 9:1250.
60. Serena C, Ceperuelo-Mallafré V, Keiran N, Queipo-Ortuño MI, Bernal R, Gomez-Huelgas R, Urpi-Sarda
M, Sabater M, Pérez-Brocal V, Andrés-Lacueva C: Elevated circulating levels of succinate in human
obesity are linked to specific gut microbiota. ISME J 2018, 12:1642–1657.
Page 17/2561. Miller TL: The pathway of formation of acetate and succinate from pyruvate by Bacteroides
succinogenes. Arch Microbiol 1978, 117:145–152.
62. Johnson JL, Moore W, Moore LV: Bacteroides caccae sp. nov., Bacteroides merdae sp. nov., and
Bacteroides stercoris sp. nov. isolated from human feces. Int J Syst Evol Microbiol 1986, 36:499–
501.
63. Loy A, Pfann C, Steinberger M, Hanson B, Herp S, Brugiroux S, Neto JCG, Boekschoten MV, Schwab C,
Urich T, et al: Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri,
a Core Member of the Murine Gut Microbiota. mSystems 2017, 2:e00171-00116.
64. Wang T-Y, Zhang X-Q, Chen A-L, Zhang J, Lv B-H, Ma M-H, Lian J, Wu Y-X, Zhou Y-T, Ma C-C: A
comparative study of microbial community and functions of type 2 diabetes mellitus patients with
obesity and healthy people. Appl Microbiol Biotechnol 2020, 104:7143–7153.
65. Pedrogo DAM, Jensen MD, Van Dyke CT, Murray JA, Woods JA, Chen J, Kashyap PC, Nehra V: Gut
microbial carbohydrate metabolism hinders weight loss in overweight adults undergoing lifestyle
intervention with a volumetric diet. In Mayo Clinic Proceedings. Elsevier; 2018: 1104–1110.
66. Del Chierico F, Abbatini F, Russo A, Quagliariello A, Reddel S, Capoccia D, Caccamo R, Ginanni
Corradini S, Nobili V, De Peppo F: Gut microbiota markers in obese adolescent and adult patients:
age-dependent differential patterns. Front Microbiol 2018, 9:1210.
67. Agus A, Clément K, Sokol H: Gut microbiota-derived metabolites as central regulators in metabolic
disorders. Gut 2021, 70:1174–1182.
68. Madigan MT, Martinko JM, Parker J: Brock biology of microorganisms. Pearson Prentice Hall Upper
Saddle River, NJ; 2006.
69. Sharma V, Rodionov DA, Leyn SA, Tran D, Iablokov SN, Ding H, Peterson DA, Osterman AL, Peterson
SN: B-vitamin sharing promotes stability of gut microbial communities. Front Microbiol 2019:1485.
70. Soto-Martin EC, Warnke I, Farquharson FM, Christodoulou M, Horgan G, Derrien M, Faurie J-M, Flint
HJ, Duncan SH, Louis P: Vitamin biosynthesis by human gut butyrate-producing bacteria and cross-
feeding in synthetic microbial communities. mBio 2020, 11:e00886-00820.
71. Woting A, Pfeiffer N, Loh G, Klaus S, Blaut M: Clostridium ramosum promotes high-fat diet-induced
obesity in gnotobiotic mouse models. MBio 2014, 5:e01530-01514.
72. Zhang H, DiBaise JK, Zuccolo A, Kudrna D, Braidotti M, Yu Y, Parameswaran P, Crowell MD, Wing R,
Rittmann BE: Human gut microbiota in obesity and after gastric bypass. Proc Natl Acad Sci 2009,
106:2365–2370.
73. Kaakoush NO: Insights into the role of Erysipelotrichaceae in the human host. Front Cell Infect
Microbiol 2015, 5:84.
74. Kieler IN, Shamzir Kamal S, Vitger AD, Nielsen DS, Lauridsen C, Bjornvad CR: Gut microbiota
composition may relate to weight loss rate in obese pet dogs. Vet Med Sci 2017, 3:252–262.
75. Del Dot T, Osawa R, Stackebrandt E: Phascolarctobacterium faecium gen. nov, spec. nov., a novel
taxon of the Sporomusa group of bacteria. Syst Appl Microbiol 1993, 16:380–384.
Page 18/2576. Fernández-Veledo S, Vendrell J: Gut microbiota-derived succinate: Friend or foe in human metabolic
diseases? Rev Endocr Metab Disord 2019, 20:439–447.
77. McDonald NC, White RL: Reduction of fumarate to succinate mediated by Fusobacterium varium.
Appl Biochem Biotechnol 2019, 187:163–175.
78. Cao W, Chin Y, Chen X, Mi Y, Xue C, Wang Y, Tang Q: The role of gut microbiota in the resistance to
obesity in mice fed a high fat diet. Int J Food Sci Nutr 2020, 71:453–463.
79. Sela D, Price N, Mills D: Bifidobacteria: genomics and molecular aspects. Book Chap. 2010:45–70.
80. Wang Z, Xiao G, Yao Y, Guo S, Lu K, Sheng Z: The role of bifidobacteria in gut barrier function after
thermal injury in rats. J Trauma Acute Care Surg 2006, 61:650–657.
81. Griffiths EA, Duffy LC, Schanbacher FL, Qiao H, Dryja D, Leavens A, Rossman J, Rich G, Dirienzo D,
Ogra PL: In vivo effects of bifidobacteria and lactoferrin on gut endotoxin concentration and
mucosal immunity in Balb/c mice. Dig Dis Sci 2004, 49:579–589.
82. Wang Z-T, Yao Y-M, Xiao G-X, Sheng Z-Y: Risk factors of development of gut-derived bacterial
translocation in thermally injured rats. World J Gastroenterol 2004, 10:1619.
83. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo
C: Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56:1761–1772.
84. Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, Gibson G, Delzenne NM: Selective
increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a
mechanism associated with endotoxaemia. Diabetologia 2007, 50:2374–2383.
85. Thomson P, Santibáñez R, Rodríguez-Salas C, Flores-Yañez C, Garrido D: Differences in the
composition and predicted functions of the intestinal microbiome of obese and normal weight adult
dogs. PeerJ 2022, 10:e12695.
86. Muñoz M, Guerrero-Araya E, Cortés-Tapia C, Plaza-Garrido A, Lawley TD, Paredes-Sabja D:
Comprehensive genome analyses of Sellimonas intestinalis, a potential biomarker of homeostasis
gut recovery. Microb Genom 2020, 6.
87. Council NR: Nutrient requirements of dogs and cats. National Academies Press; 2006.
88. Blondel VD, Guillaume J-L, Lambiotte R, Lefebvre E: Fast unfolding of communities in large networks.
J Stat Mech: Theory and Experiment 2008, 2008:10008.
Tables
Page 19/25Table 1
Analyzed chemical composition of the experimental diet.
Chemical composition (%DM or as stated)
DM, % 88.0
CP 37.5
EE 22.7
CF 3.41
NFE1 21.6
Ash 14. 8
Ca 1.00
P 0.80
ME2, Mcal/kg 3.68
DM, dry matter; CP, crude protein; EE, ether extract; CF, crude fiber; NFE, nitrogen-free extract; Ca,
calcium; P, phosphorus; ME, metabolizable energy.
1NFE% = 100 - (CP + EE + CF + Ash + moisture)
2Estimated based on National Research Council [87].
Table 2
Differences in body weight and body condition score between obeses and normal dogs.
Treatment1
Parameters HBCS NBCS SEM P-value
Initial BW, kg 11.88 10.05 0.590 0.0261
Final BW, kg 13.10 10.07 0.772 0.0045
BW gain, kg 1.22 0.02 0.375 0.0287
Initial BCS 7.0 5.2 0.601 0.0262
Final BCS 8.0 5.0 0.365 < 0.0001
SEM, standard error of the mean.
1HBCS, high body condition score (BCS) group (BCS range: 7–9, 4 males and 2 females, all 2-year-
old); NBCS, normal BCS group (BCS range: 4–6, 4 males and 2 females, all 2-year-old).
Page 20/25Table 3
Differences in alpha diversity measurements of the fecal microbiota by obesity.
Treatments1
Parameters HBCS NBCS SEM2 P-value
Observed ASVs 180 174 19.1 0.8220
Chao1 estimates 181 175 19.5 0.8268
ACE 180 175 19.4 0.8344
Evenness 0.698 0.636 0.0261 0.1044
Shannon’s index 5.22 4.72 0.251 0.1830
Simpson’s index 0.934 0.897 0.0181 0.1361
ASV, amplicon sequencing variant; ACE, abundance-based coverage estimator.
1
HBCS, high body condition score (BCS) group (BCS range: 7–9, 4 males and 2 females, all 2-year-
old); NBCS, normal BCS group (BCS range: 4–6, 4 males and 2 females, all 2-year-old).
2SEM, standard error of the mean.
Page 21/25You can also read

















































