NSrp70 is a lymphocyte-essential splicing factor that controls thymocyte development
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
5760–5778 Nucleic Acids Research, 2021, Vol. 49, No. 10 Published online 25 May 2021
https://doi.org/10.1093/nar/gkab389
NSrp70 is a lymphocyte-essential splicing factor that
controls thymocyte development
Chang-Hyun Kim 1,2,† , Sang-Moo Park1,2,† , Sun-jae Lee1,† , Young-Dae Kim1,2 ,
Se-Hwan Jang1 , Seon-Min Woo3 , Taeg-Kyu Kwon3 , Zee-Yong Park1 , Ik-Joo Chung4 ,
Hye-Ran Kim1,2,* and Chang-Duk Jun 1,2,*
1
School of Life Sciences, Gwangju Institute of Science and Technology (GIST), Gwangju 61005, Korea, 2 Immune
Synapse and Cell Therapy Research Center, Gwangju Institute of Science and Technology (GIST), Gwangju 61005,
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
Korea, 3 Department of Immunology, School of Medicine, Keimyung University, Daegu 42601, Korea and
4
Department of Hematology-Oncology, Immunotherapy Innovation Center, Chonnam National University Medical
School, Hwasun 58128, Korea
Received September 18, 2020; Revised April 4, 2021; Editorial Decision April 27, 2021; Accepted April 30, 2021
ABSTRACT gene expression (1). Proteins belonging to the nuclear
speckles include small nuclear ribonucleoprotein particles
Alternative pre-mRNA splicing is a critical step to (snRNPs) and many non-snRNP splicing factors such as
generate multiple transcripts, thereby dramatically SR (serine/arginine) proteins and heterogeneous nuclear ri-
enlarging the proteomic diversity. Thus, a common bonucleoproteins (hnRNPs) (2). Previously, our laboratory
feature of most alternative splicing factor knockout discovered a new SR-related protein named nuclear speckle-
models is lethality. However, little is known about related protein 70 (NSrp70), which co-localizes and inter-
lineage-specific alternative splicing regulators in a acts physically with SRSF1 and SRSF2 in nuclear speckles
physiological setting. Here, we report that NSrp70 through its RS-like region. NSrp70 modulates alternative
is selectively expressed in developing thymocytes, splicing site selection of some mRNAs, including Fas v6,
highest at the double-positive (DP) stage. Global Tra21 v2 and CD44 exon v5 (3). However, little is known
splicing and transcriptional profiling revealed that about the significance of NSrp70 in a physiological setting.
One clue is that, like other major splicing factors, a knock-
NSrp70 regulates the cell cycle and survival of thy-
out of NSrp70 resulted in an embryonic lethal phenotype
mocytes by controlling the alternative processing of (3), suggesting that this protein is essential for early devel-
various RNA splicing factors, including the onco- opment. Interestingly, NSrp70 is predominantly expressed
genic splicing factor SRSF1. A conditional-knockout in immune function-related tissues or organs (3), suggesting
of Nsrp1 (NSrp70-cKO) using CD4Cre developed se- its potential significance in the immune system. This fact led
vere defects in T cell maturation to single-positive us to investigate whether NSrp70 plays a role as a lineage-
thymocytes, due to insufficient T cell receptor (TCR) selective splicing regulator in the immune system.
signaling and uncontrolled cell growth and death. The immune system is where rapid cell division and com-
Mice displayed severe peripheral lymphopenia and plex developmental processes occur during hematopoiesis.
could not optimally control tumor growth. This study Notably, T cell development in the thymus is a highly inte-
establishes a model to address the function of grated process with distinct developmental stages defined by
the expression of the co-receptors CD4 and CD8 (4). These
lymphoid-lineage-specific alternative splicing factor
events are primarily driven by the expression and activation
NSrp70 in a thymic T cell developmental pathway. of the T cell receptor (TCR) complex. In particular, precise
control of TCR signaling is critical for complete thymocyte
INTRODUCTION development (5–7). Further, many factors that regulate the
cell cycle, apoptosis, survival, differentiation, and prolifera-
Nuclear speckles are irregular and non-membranous punc- tion are also cooperatively involved in thymocyte develop-
tuate structures that serve as storage/assembly sites for ment (8–11).
pre-mRNA splicing machinery, and therefore regulate
* To
whom correspondence should be addressed. Tel: +82 62 715 2506; Email: cdjun@gist.ac.kr
Correspondence may also be addressed to Hye-Ran Kim. Tel: +82 62 715 2567; Fax: +82 62 715 2546; Email: perfectfan@hanmail.net
†
The authors wish it to be known that, in their opinion, the first 3 authors should be regarded as joint First Authors.
Present address: Young-Dae Kim, Institute for Drug Discovery, CrystalGenomics, Inc., Korea Bio Park, 700 Daewangpangyo-ro, Bundang-gu, Seongnam-si,
Gyeonggi-do 13488, Korea.
C The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), which
permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.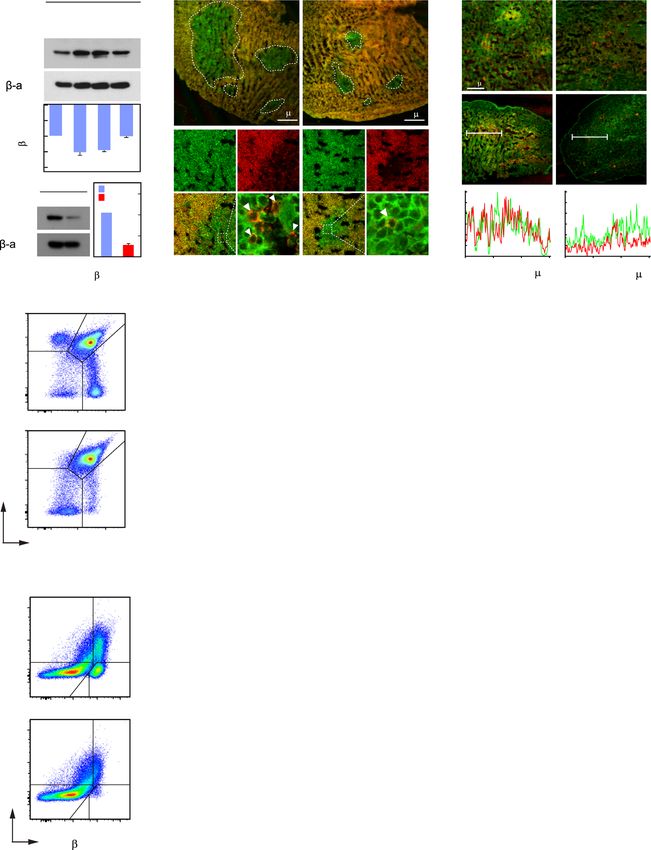
Nucleic Acids Research, 2021, Vol. 49, No. 10 5761
A previous report demonstrated that conditional deletion mCD4, CD62L, Foxp-3, peridinin chlorophyll protein com-
of the prototypical SR protein SRSF2 (also known as SC35) plex (PerCP) Cy5.5-conjugated CD8␣, CD44, TCR and
in the thymus causes a defect in T cell maturation (12). An for phycoerythrin (PE)-conjugated mouse CD25, CD24,
hnRNP L knockout resulted in decreased thymic cellular- CD69, CXCR4, CCR7, CD3ε and TCR , and for PE-
ity caused by a partial block at the transition between DN4 Cy7-conjugated mouse CD127 (IL7R␣), CD69 and Bcl-2
and DP thymocytes and higher proliferation rates in DN4 were purchased from eBioscience (San Diego, CA, USA).
thymocytes (13). However, the actions of SRSF2 and hn- Total RNA isolation reagent was purchased from Molec-
RNP L are not restricted to lymphocytes; indeed, they im- ular Research Center, Inc. (Cincinnati, OH, USA). PCR
pact almost all biological processes (14). To our surprise, premix and restriction enzyme were purchased from En-
NSrp70 was highly and selectively detected in developing zynomics (Daejeon, Korea). Plasmid DNA purification kit
lymphocytes––highest in DP thymocytes––, strongly sug- and WEST-ZOL western blot detection kit were purchased
gesting that NSrp70 may have lymphocyte-selective activity from Intron Biotechnology (Seongnam, Korea). PrimeS-
during the developmental process. TAR HS DNA polymerase was purchased from TaKaRa
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
To address a potential role of NSrp70 in T cell de- Bio (Shiga, Japan). Magna RIPTM Kit was purchased from
velopment, we created a conditional-knockout using the EMD Millipore (Billerica, MA, USA). All cell culture ma-
CD4Cre-loxP strategy. NSrp70 deletion (Nsrp1f/f CD4Cre) terials used in this work were from Life Technologies (Carls-
profoundly perturbed the late development of DP thy- bad, CA, USA). Tissue-Tek OCT was purchased from
mocytes, leading to a significant reduction of CD4+ and Sakura Finetek (Torrance, CA, USA). Complete protease
CD8+ single positive (SP) cells in the thymus and periph- inhibitors and phosphatase inhibitors were purchased from
eral lymphoid tissues. Global splicing profiling revealed that Roche Applied Science (Indianapolis, IN, USA). Aqua C18
NSrp70 controls the processing of cell cycle-related RNA (particle size 5 m) reverse phase column material was pur-
splicing factors, including Srsf1, Srsf7 and Son (15–17), chased from Phenomenex (Torrance, CA, USA). Strong
and spliceosome-related factors, including Sf3b1 and U2af2 cation exchange (particle size 5 m) column material was
(18). NSrp70 also organizes the nuclear speckles by physi- purchased from Whatman (Maidstone, Germany).
cally integrating the RNA splicing proteins and governs the
cell cycle and survival at the stage of TCR-mediated positive
Biological resources
selection to produce CD4+ or CD8+ SP T cell populations.
Cells. Mouse thymocytes were purified from the mouse
thymus. Mouse splenocytes and lymphocytes were dis-
MATERIALS AND METHODS persed and purified to CD4+ , CD8+ , CD19+ and CD11c+
populations by MACS cell separation (Miltenyi Biotec).
Reagents and antibodies
Naı̈ve CD3+ T cells were purified from the mouse spleen
Antibodies against NSrp70 (HPA015603) and mTCR and lymph nodes by negative selection using a T cell en-
(MABF931), tetramethylrhodamine (TRITC)-phalloidin, richment column (R&D Systems). To generate mouse T
poly-L-lysine (PLL), A23187, polybrene, and phorbol cell blasts, naı̈ve CD3+ T cells were incubated in 2 g/ml
12-myristate 13-acetate (PMA) were purchased from anti-CD3/28-coated culture plates with 100 U/ml rIL-2
Sigma-Aldrich (St. Louis, MO, USA). Anti-TCR -chain for 48 h and cultured for a further 5 days with 100 U/ml
(ab40804) antibody was purchased from Abcam (Cam- rIL-2. Mouse splenocytes were dispersed and purified into
bridge, MA, USA). Antibodies against PLC␥ 1 (sc-7290), CD4+ , CD8+ and CD19+ populations using the EasySep
phospho-Vav1 (sc-16408-R), SON (NREBP, sc-398508), magnetic separation system (Stemcell Technologies, Van-
HDAC1 (10E2), SRSF1 (SF2/ASF, sc-33652) were pur- couver, Canada) or MACS cell separation (Miltenyi Biotec,
chased from Santa Cruz Biotechnology (Dallas, TX, Bergisch Gladbach, Germany). The purity of each popula-
USA). Antibodies against phospho-TCR -chain (PA5- tion was confirmed as >95% by flow cytometry. HEK293T
37512), CD45ALL (30-F11), CD45RB and mCherry (PA5- cells (CRL-1573, ATCC), B16F10 cells (CRL-6475, ATCC)
34974) and Fluo-3/AM were purchased from Invitrogen and EO771 cells (CRL-3461, ATCC) were maintained in
(Carlsbad, CA, USA). Antibodies against -actin (8457), DMEM supplemented with 10% FBS, penicillin and strep-
Zap70 (2705S), phospho-Zap70 (2701S), phospho-PLC␥ 1 tomycin.
(2821L), PKC (12206S), phospho-PKC (9376), Erk1/2
(9102S), phospho-Erk1/2 (9101S), p38 kinase (9212S), Constructs. The mammalian expression vector of the
phospho-p38 kinase (9215S), rabbit IgG (7074) and mouse Fas minigene was a gift from Dr Michael Sattler (GSF-
IgG (7076) were purchased from Cell Signaling Technology National Research Center for Environment and Health,
(Danvers, MA, USA). Anti-CD2 antibody (130-100-617) Neuherberg, Germany). pEGFP-C1 NSrp70, and pEGFP-
was purchased from Miltenyi Biotec (Bergisch Gladbach, C1 RS1M constructs were generated using a previously
Germany). 4 ,6-Diamidino-2-phenylindole dihydrochloride reported method (3,19). The cDNA of SRSF1, U2AF1,
(DAPI) was purchased from Molecular Probes (Eugene, hnRNP U, U170K was obtained by RT-PCR using total
OR, USA). Anti-CD28 (ab205136) antibody was purchased RNA prepared from Jurkat T or HEK293T cells as a tem-
from BD Biosciences (San Jose, CA, USA). Hybridoma plate. Then, each cDNA was inserted into the pCS4-3Myc,
cell lines for anti-CD3ε (145-2C11; CRL-1975), and anti- pmCHerry-C1 and pHJ-1 vectors by in-frame fusion.
CD28 (PV1; HB-12352) were purchased from the Amer-
ican Type Culture Collection (Manassas, VA, USA). An- Animals. For generation of NSrp70-cKO mice, we
tibodies for fluorescein isothiocyanate (FITC)-conjugated used the gene-targeting vector with which the two-loxP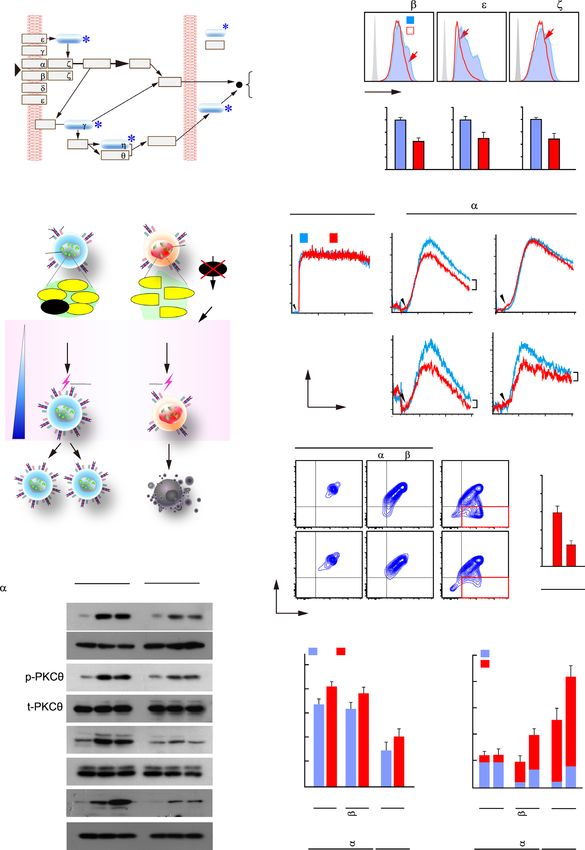
5762 Nucleic Acids Research, 2021, Vol. 49, No. 10
and two-frt strategy was employed to obtain homolo- nology). Beads were collected, washed with PBS, and re-
gous recombination in embryonic stem (ES) cells. After suspended in equal volumes of 5× SDS loading buffer. Im-
successful germ line transmission (Macrogen, Korea), munoprecipitated proteins were separated by SDS-PAGE
chimera mice (Nsrp1+/flox-frt-neo ) were generated, back- on 10, and 12% gels and analyzed by western blot analysis
crossed onto C57BL/6 background for six generations, as described above.
and the off-spring were interbred. The Nsrp1+/flox-frt-neo
mice were crossed with -actin-driven Saccharomyces
cerevisiae-enhanced FLP1 recombinase variant (FLPe) mRNA-seq and data analysis
deleter mice (The Jackson Laboratory) to facilitate in Total RNA was purified from sorted-DP thymocytes us-
vivo frt-neo deletion. The Nsrp1flox/flox (Nsrp1f/f will be ing TRIzol reagents (Molecular Research Center, Cincin-
used herein to indicate Nsrp1flox/flox for simplicity) mice nati, OH, USA). A NanoDrop spectrophotometer (Thermo
were further crossed with CD4cre transgenic mice (The Fisher Scientific Inc., USA) was used to confirm total RNA
Jackson Laboratory) to generate Nsrp1f/f CD4cre mice. quantity and integrity. Purified samples were then prepared
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
All mice were housed in specific pathogen-free conditions. using a standard polyA-enriched library preparation proto-
All experimental methods and protocols were approved col (NEBNext Ultra II Directional RNA-Seq Kit standard
by the Institutional Animal Care and Use Committee of protocols) implemented by ebiogen (Seoul, Korea). Ribo-
the School of Life Sciences, Gwangju Institute of Science somal RNA was removed prior to proceeding. Sequencing
and Technology (Gwangju, Korea), and carried out in was performed using the Illumina NovaSeq 6000 that gen-
accordance with their approved guidelines. erates paired end reads of 101 bp. Illumina Casava1.8 soft-
ware used for basecalling. Sequenced reads were trimmed
for adaptor sequence, and masked for low-complexity or
Cell transfection and lentiviral infection
low-quality sequence using fastx trimmer, then mapped to
HEK293T cells were transfected with various constructs UCSC mm10 whole genome (Mus musculus genome) us-
(pHJ-1 NSrp70, and pHJ-1 SRSF1) by Lipofectamine ing Tophat. The mapped reads (FPKM) were further ana-
2000 (Life Technologies) according to the manufacturer’s lyzed with Cufflink to calculate the level of gene expression.
instructions. To establish stable cell lines, the pHJ-1 vec- The sequencing data were deposited into GEO repository
tor was co-transfected with lentiviral packaging vectors into with the accession number GSE168379. For detailed differ-
HEK293T cells. After 48 h, the supernatants were collected entially expressed gene (DEG) analysis, raw gene counts of
and spin-infected into HEK293T cells by centrifugation at the transcriptome were normalized by size factors and com-
800 × g for 90 min in the presence of polybrene (8 g/ml). pared between wild-type and Nsrp70-KO samples by neg-
ative binomial tests using the R DESeq2 package. Based
on significantly differentially expressed genes (adjusted P-
Western blot analysis
value < 0.05), we performed gene ontology enrichment
Cells or homogenized tissues from a C57BL/6 mouse were tests among up-regulated or down-regulated genes using the
lysed in ice-cold lysis buffer (50 mM Tris–HCl pH 7.4, con- modified Fisher’s Exact tests with the RDAVIDWebService
taining 150 mM NaCl, 1% Triton X-100, one tablet of com- package (P-value < 0.01). The alternative splicing events
plete protease inhibitors and phosphatase inhibitors) for were analyzed using the MISO package with the annotation
30 min on ice. Cell lysates were centrifuged at 16 000 × of all known alternative splicing events (20). The significant
g for 25 min at 4◦ C, and the supernatant was eluted with differentially spliced events were determined by Bayes’ fac-
SDS sample buffer (100 mM Tris–HCl pH 6.8, 4% SDS tor (BF) and Percent spliced in values (PSI, ) (BF ≥ 10
and 20% glycerol with bromophenol blue) and boiled for and PSI ≥ 0.2). Gene ontology and KEGG analysis were
5 min. The proteins were separated through 10–12% SDS- performed using the functional annotation tool of DAVID
PAGE gels and were transferred to a nitrocellulose mem- (21) to search for enriched pathways of associated alterna-
brane by means of a Trans-Blot SD semidry transfer cell tive splicing events. The functional associations of NSrp70
(Bio-Rad, California, USA). The membrane was blocked in targets were analyzed using protein interaction dada from
5% skim milk for 1 h, rinsed, and incubated with intended the STRING database (22), generating a set of functional
antibodies in TBS containing 0.1% Tween 20 (TBS-T) and interaction networks.
3% skim milk overnight. Excess primary antibody was then
removed by washing the membrane four times in TBS-T.
Reverse transcription PCR (RT-PCR) and real-time quanti-
The membrane was then incubated with peroxidase-labeled
tative RT-PCR (qRT-PCR)
secondary antibody (0.1 g/ml, anti-rabbit or -mouse) for 1
h. After three washes with TBS-T, bands were visualized by Total RNA was isolated from cells or homogenized tis-
WEST-ZOL western blot detection kit (Intron Biotechnol- sues of C57BL/6 mice with TRIzol reagents (Molecular
ogy) and the membranes were then exposed to X-ray film. Research Center, Cincinnati, OH, USA) and reverse tran-
scribed using RT-Premix (Intron Biotechnology). PCR was
performed with the respective forward and reverse sequence
Immunoprecipitation (IP)
pairs as indicated in Supplementary Table S1. The expres-
Cell lysates were precleared, and supernatants were incu- sion levels of mouse Cdk1, Cdca3, Rpl26, Uba52, Atp5pd,
bated overnight with antibodies at 4◦ C, followed by incuba- Ndufs5, Fyn, Plcg1, Ccr7, Junb, Ptk2b and Bag6 were
tion with protein A/G agarose beads (Santa Cruz Biotech- evaluated by qRT-PCR. Amplification was performed in a
Nucleic Acids Research, 2021, Vol. 49, No. 10 5763
StepOne real-time PCR system (Applied Biosystems, Nor- Flow cytometry analysis
walk, CT, USA) for continuous fluorescence detection in a
Thymus, spleen and lymph nodes were dissected and passed
total volume of 10 l of cDNA/control and gene-specific
through a 40 m cell strainer (BD Biosciences). The passed
primers using SYBR Premix Ex Taq (TaKaRa Bio). The
cells were washed with PBS and remaining erythrocytes
mRNA levels of the target genes were normalized relative to
were lysed with a commercially available hemolysis buffer
those of Gapdh using the following formula: relative mRNA
(Morphisto, Frankfurt am Main, Germany). Isolated cells
expression = 2− (Ct of target gene − Ct of Gapdh ), where
were washed and stained for flow cytometry with antibodies
Ct is the threshold cycle value. In each sample, the expres-
against CD4, CD8, CD25, CD24, Foxp-3, TCR-, CD69,
sion of the analyzed gene was normalized to that of Gapdh
TCR-ε and TCR- for 30 min at 4◦ C. Data were acquired
and described as the mRNA level relative to Gapdh.
on a FACSCanto (BD Biosciences, San Jose, CA, USA)
and analyzed with FlowJo software (Tree Star, Ashland,
OR, USA). Appropriate isotype controls were included
RNA immunoprecipitation
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
and gates were set according to isotype-matched controls.
RNA immunoprecipitation (RIP) experiments were per- For intracellular staining, cells were fixed and permeabi-
formed using a Magna RIP RNA-Binding Protein Im- lized with intracellular fixation and permeabilization buffer.
munoprecipitation Kit according to the manufacturer’s in- Fixed cells were stained for flow cytometry with antibodies
structions (Millipore) using a modified version of a pre- against Bcl-2 and Ki67 for 30 min at 4◦ C. Total cellular-
viously described method (23). Briefly, HEK293T cells ity was determined by counting the live cells. Absolute cell
(1 × 107 ) transfected with the indicated constructs were numbers were calculated on the basis of the percentage of
lysed on ice in complete RIP lysis buffer (Millipore). each population and represented as the mean ± SD. The
GFP or IgG control antibodies were pre-coated onto mag- mice in these experiments were 6∼8 weeks of age.
netic beads protein A/G for 30 min at room temperature.
Antibody-coated beads were washed two times with RIP Isolation of oocyte, morula, and embryos and immunofluores-
wash buffer (Millipore) using a magnetic separator. Pro- cence staining
tein lysates were then incubated with rabbit anti-GFP- or
C57BL/6 mice were injected with 10 IU/mouse of PMSG
control IgG-coated beads overnight at 4◦ C. The immuno-
(pregnant mare serum gonadotrophin). After 48 h, mice
precipitated samples were then centrifuged and washed
were re-stimulated with 10 IU/mouse of hCG (human
with ice-cold RIP wash buffer six times using a mag-
chorionic gonadotropin). After 13 h, mice were terminated
netic separator. After the final wash, the RNA–protein
to obtain oocytes. Mouse embryonic day is related to the
complexes were dissociated from the beads by incubat-
female presence of a vaginal plug indicating that mating oc-
ing in proteinase K buffer (Millipore) for 30 min at 55◦ C.
curred. Day 1 of pregnancy was defined as the day on which
The supernatants were collected using a magnetic separa-
the vaginal plug was observed. At 2 d post coitum (dpc),
tor. RNA extraction from the supernatant was performed
mice were terminated to obtain morula; in the same way we
with phenol:chloroform:isoamyl alcohol (125:24:1) (Sigma-
could obtain E11.5 (11.5 dpc) to E18.5 (18.5 dpc) stages of
Aldrich) and precipitated using ethanol supplemented with
embryos.
salt solution I, salt solution II, and precipitation enhancer
overnight. The RNA pellets were then washed and sus-
Freeze sectioning and confocal microscopy
pended with RNAse-free water. The cDNA was transcribed
from total RNA using Reverse Transcript PCR Premix Embryos were snap-frozen in isopentane precooled close to
(Intron Biotechnology). PCR was performed as described its freezing point in liquid nitrogen and then 5-m thick sec-
above. tions were prepared and mounted onto poly-L-lysine coated
glass slides. Sections were air-dried under a fan for 2–24
h, after which the embryos were incubated first with anti-
In vivo splicing assays NSrp70 antibody and then with the FITC-conjugated sec-
ondary antibody for NSrp70 and TRITC-phalloidin for
In vivo splicing assays were performed as previously de-
actin, for 30 min at 37◦ C. For frozen sections of immune
scribed (24). Briefly, a splicing reporter minigene was
related tissues, mice were anaesthetized, and the thymus,
co-transfected with an increasing amount of wild-type
spleen and lymph node of each mouse was collected and
GFP NSrp70 construct, together with mCherry-tagged
fixed with 4% paraformaldehyde for 4 h, followed by treat-
SRSF1 in HEK293T cells. Empty plasmids were added
ment with a 30% sucrose solution in PBS overnight at 4◦ C.
to ensure that the same amount of DNA was transfected.
Tissue samples were embedded in optimal cutting tempera-
Forty-eight hours after transfection, total RNA was ex-
ture (OCT) medium and stored at –80◦ C. The samples were
tracted from cells using the Total RNA isolation reagent
sectioned to a 7-m thickness. The slides were examined us-
(Molecular Research Center). The cDNA was transcribed
ing a FV1000 confocal laser scanning microscope (Olym-
from 2 g of total RNA using Reverse transcript PCR pre-
pus, Tokyo, Japan) equipped with 40×, 60× and 100× ob-
mix (Intron Biotechnology). PCR for the FAS minigene was
jectives.
performed as previously described (19). The PCR products
were analyzed with a 1–1.5% agarose gel and the splicing
PCR genotyping assay of progenies
pattern was quantified using ImageJ (ImageJ is a public do-
main java image processing program inspired by National Tails of progenies were lysed in 1.5 ml tubes contain-
Institutes of Health Image). ing tail lysis buffer (DirectPCR Lysis Reagent, VIAGEN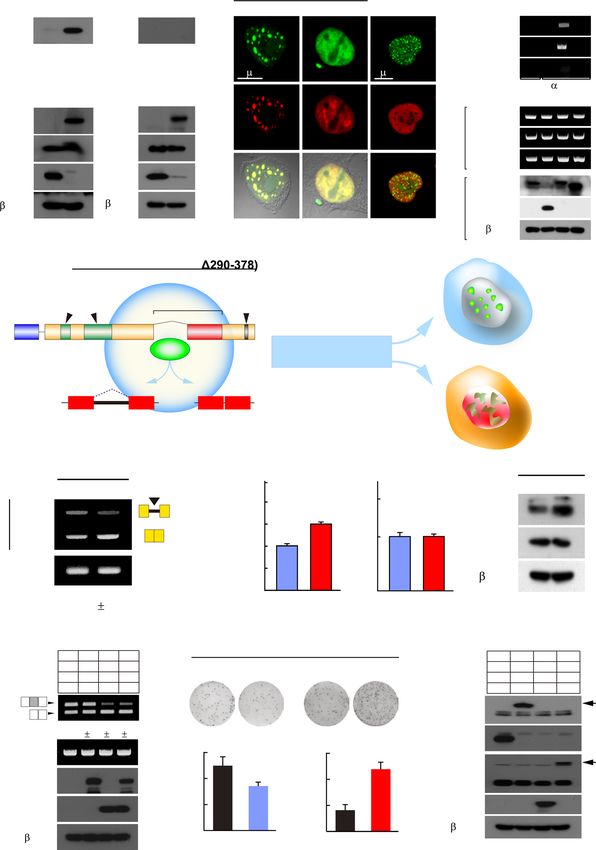
5764 Nucleic Acids Research, 2021, Vol. 49, No. 10
Biotech), and 2 g lysate was used for PCR genotyping. Tumor animal models
The PCR primers for the wild-type allele were located in
Control B16F10 melanoma cells (2 × 106 cells/0.1 ml me-
exon 1 (21T 93–1: 5 -ATATACACGTCGGCGTCAGC-3 )
dia per mouse) were injected subcutaneously into the two
and exon 2 (21T93-3R: 5 -CAAAATAAGCCCATACCTG
dorsal flank regions of Nsrp1f/f (WT) and Nsrp1f/f CD4Cre
CGTAA-3 ), and the PCR primers for the trap allele were
(KO) mice (6–8 weeks old). After 2 weeks, mice were sacri-
located between exon 1 and exon 2 (21T93-9: 5 -GAAG
ficed, tumors were dissected, photographed, and weighed.
AGAGGCCCATTGGTTG-3 , SA-5SA: 5 -GGGCAAGA
ACATAAAGTGACC-3 ). Primers 21T93-1 and 21T93-3R
were used to amplify 1,737 base pairs of intron 1 (35 cy- Computational resources
cles of 1 min at 94◦ C, 1 min at 58◦ C and 2 min at 72◦ C), TCGA database and cBioPortal. cBioPortal for Cancer Ge-
and primers 21T93-9 and SA-5AS were used to amplify 597 nomics (http://cbioportal.org) provides an open-access web
bp that consisted of the partial trap vector and the intron 1 resource for exploring, visualizing and analyzing multidi-
sequence (35 cycles of 1 min at 94◦ C, 15 s at 56◦ C, and 30 s mensional cancer genomic data from TCGA (27). In the
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
at 72◦ C). present study, TCGA PanCancer Atlas studies provided by
default on the cBioPortal were selected for further analysis
of NSRP1 gene alterations. The cancer types summary and
Ca2+ flux analysis survival tabs were applied according to the online instruc-
Thymocytes and lymphoblasts were first labeled for 1 h tions of cBioPortal.
at 37◦ C with 4 g/ml Fluo-3/AM (Invitrogen) and then
washed and resuspended with ice-cold PBS. Cells (3 × Statistical analyses
106 ) were labeled on the surface for 30 min on ice with Mean values were calculated using data taken from at least
PE-conjugated anti-CD4 (H129.19; BD Biosciences) and three independent experiments conducted on different days.
PE-Cy3-conjugated anti-CD8 (53–6.7; BD Biosciences), Where significance testing was performed, unpaired Stu-
and then incubated with biotinylated anti-CD3 (5 g/ml; dent’s t tests and one-way analysis of variance tests were
145–2C11) and anti-CD28 (5 g/ml; 37.51). Labeled cells used. Differences between groups were considered signifi-
were warmed for 20 min at room temperature and then cant at P-value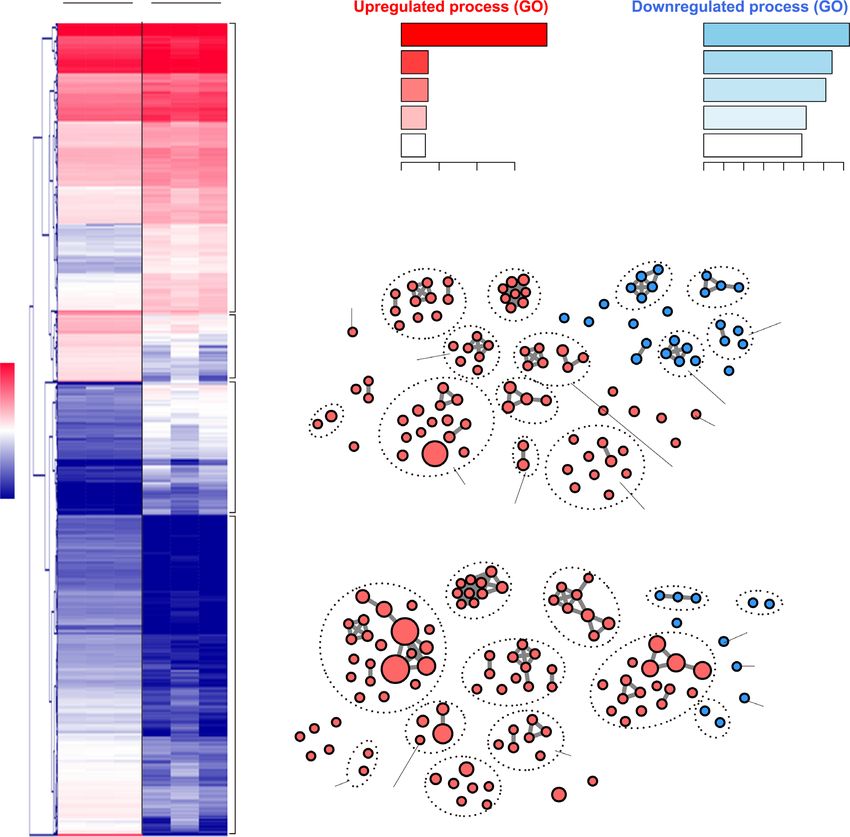
Nucleic Acids Research, 2021, Vol. 49, No. 10 5765
A B C
E 11 13 15 17 200
Actin NSrp70 Nsrp1
(240 bp) 160 n = 147
Number of litters
mGapdh
(170 bp) 120
Oocyte
2
Relative Nsrp1
to mGapdh
DAPI Merge 1.5 80
NS NS NS
1 n = 38
40
0.5 n=0
10 µm 0
11 13 15 18
+
T
T
+/
G
G
+/
T/
Actin NSrp70
G
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
D *
S. ach
* *
Sp us
Th cle
Kid en
Liv y
L.N .
L.N
Morula
M. s
ne
ym
*
Mu rt
om
He n
Te g
er
sti
le
.
s
ai
a
n
I.
.
L.I
Lu
Br
St
M
DAPI Merge
100
NSrp70
70
10 µm 55
-Actin
42
E11.5 a
b
CD11c+
E
CD11b+
CD11c+
CD19+
CD19+
CD4+
CD8+
CD4+
CD8+
SP
SP M
Embryo
50 µm 100 Nsrp1
NSrp70
a 70 (240 bp)
b ZOOM
-Actin 42 mGapdh
0 (170 bp)
protein to -actin
Relative NSrp70
0
Relative Nsrp1
1
to mGapdh
2 1
3 2
*
p=2E-02
4 3 * p=4E-03
p=2E-04 *
NSrp70 DAPI Actin 5 * *
p=2E-04 p=1E-03
4
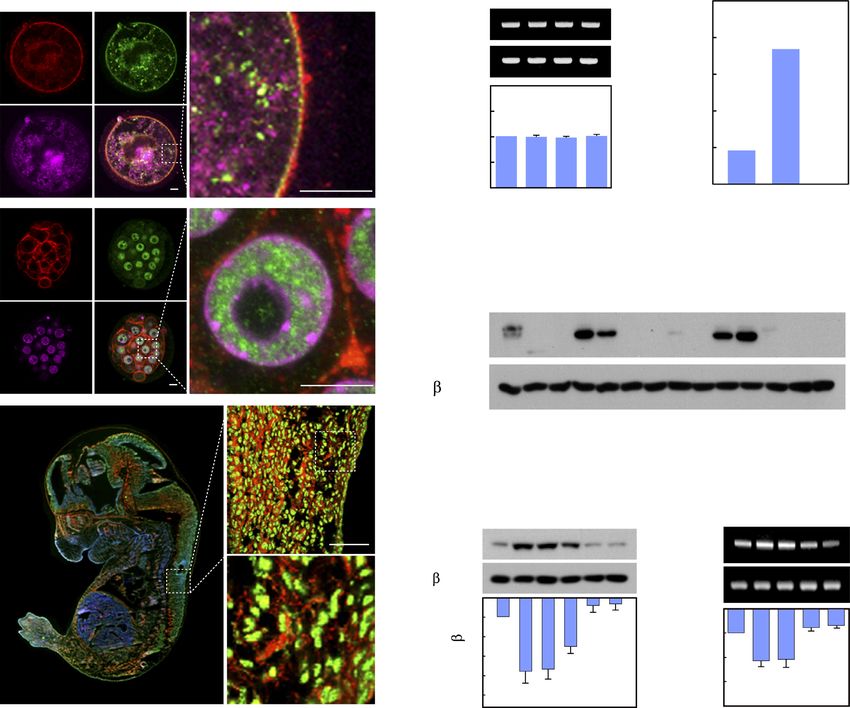
Figure 1. NSrp70 is preferentially expressed in early embryonic tissues and in lymphoid cells. (A) Immunohistochemistry for NSrp70. Oocyte (top), morula
(middle), and fetal embryo E11.5 (bottom) were isolated from WT mice, stained with anti-NSrp70 antibody followed by FITC-conjugated 2nd antibody
(green), TRITC-phalloidin (red), and DAPI (magenta and blue), and then visualized by confocal microscopy. (B) Fetal embryos from gestation day 11.5,
13.5, 15.5 and 17.5 mice were isolated. Nsrp1 mRNA was detected by RT-PCR (blots) and density was represented by bar graph. mGapdh was used as a
loading control. E, embryonic day; bp, base-pair. (C) Intercrossing the heterozygous Nsrp1+/GT mouse produced no offspring homozygous for the allele
containing the gene-trap vector. GT, gene-trap vector; +/+, wild-type; +/GT, heterozygous Nsrp1+/GT ; GT/GT, homozygous Nsrp1GT/GT . (D) Tissue
distribution of NSrp70 was determined by western blot analysis in 8-week old mice. M.L.N, mesenchymal lymph node; L.N., lymph node; S.I., small
intestine; L.I., large intestine; M, molecular mass (KDa). (E) Western blot (left) and RT-PCR (right) for NSrp70 in mouse immune cells. -actin and
mGapdh were shown as loading controls. SP, splenocytes. All data shown are representative of three independent experiments. The bar graphs indicate the
mean ± standard deviation of the indicated protein blot or RNA gel densitometry presented with respect to -actin or mGapdh (B and E).
no overt signs of weight difference, nor differences in the We assessed the populations of thymocytes to determine
sizes of immune organs such as thymus, spleen, and lymph the populations and numbers of CD4+ and CD8+ cells. In
node as compared to those of the wild-type (WT, Nsrp1f/f ) agreement with the results of the thymic tissue section, a
(Supplementary Figure S1B and C). NSrp70 was upregu- significant reduction of CD4 and CD8 SP thymocytes in
lated at the DP stage, but significant reduction of NSrp70 NSrp70-cKO mice was observed as compared with that of
was confirmed in NSrp70-cKO thymocytes (Figure 2A). Al- WT mice, while total numbers of thymocytes were not al-
though there was no sign of morphological changes, inter- tered substantially (Figure 2D and E). In addition, the pop-
estingly, smaller areas of the medulla accompanied by sig- ulations and numbers of DN and DP thymocytes were not
nificant reduction of CD4+ and CD8+ T cell zones were dramatically altered (Figure 2D and E).
shown in NSrp70-cKO thymus (Figure 2B, arrowheads, and We observed a considerable decrease in the number of
Supplementary Figure S1C). In addition, CD4+ and CD8+ CD4+ and CD8+ SP thymocytes in NSrp70-cKO mice (Fig-
SP signals were low in the spleen and lymph nodes (Figure ure 2E). To clarify at which stage thymocyte development
2C), suggesting that thymocytes are not properly developed was blocked, we quantified cells at four distinct develop-
at the DP stage.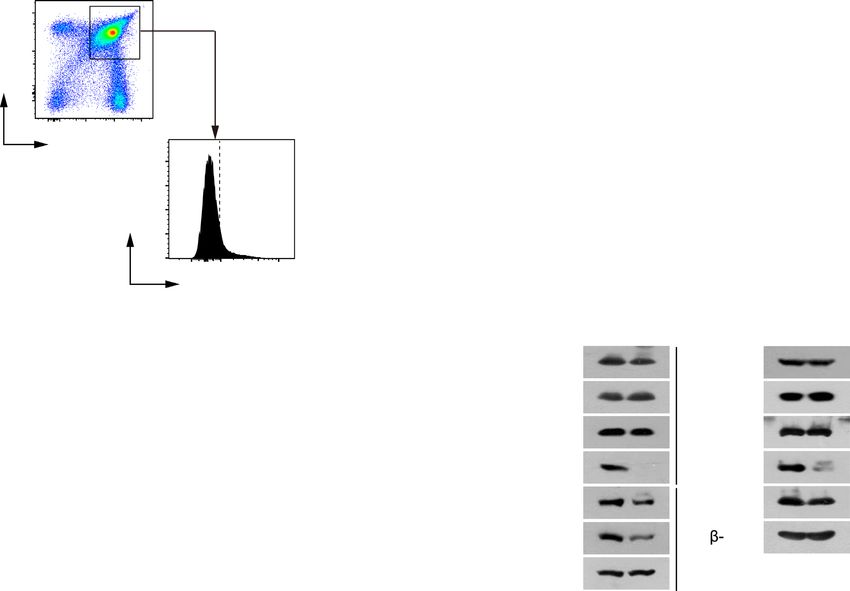
5766 Nucleic Acids Research, 2021, Vol. 49, No. 10
A WT B WT KO C WT KO
CD4 CD4CD8
+ SP+ SP M
C CD8
N P D 4 D8 M M
D D C C
Spleen
100 M C
NSrp70 70 M
M
55
200 m
ctin
42 M M
CD4CD8
Relative NSrp70
0 M
Lymph node
200 m 200 m
10X 10X
to -actin
0.5 a b a b
1 CD4 CD8 CD4 CD8 psi
NS
1.5 *
2 * p=2E-05
p=5E-03
DP WT 60X 60X 60X 60X
1.5 30
Intensity (X100)
30
p=1.3E-04
WT KO KO Merged Merged
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
1 20 20
NSrp70
0.5 10 10
ctin *
60X 60X
0 0 0
Relative NSrp70 a b a b
to -actin Distance (450 m) Distance (450 m)
D E DN DP CD4SP CD8SP
WT105 3.28 84.6 12 120 20 5
WT p=0.012
Population (%)
10 NS 100 *
104 KO 15 4
8 80
p=0.0015 3 p=1E-05
103 6 60 10
4 40 * 2 *
5
2 20 1
0
2.70 9.32 0 0 0 0
0 103 104 WT KO WT KO WT KO WT KO
105 Total
KO 0.80 90.9 8 25 8 200
WT NS 150
104 KO NS
Cells (x106)
6 20 6 150 NS
100 p=5E-04
CD8
103
15 p=3E-04
4 4 100
50
10 * *
0 2 5 2 50
6.42 1.77 0 0 0 0 0
0 103 104 WT KO WT KO WT KO WT KO WT KO
CD4
F G Gated on H
hi
Gated on I Gated on II Gated on III Gated on IV
Total III + IV (TCR )
35 200 40 40 40
WT II 4 III
Cells (x106)
10
Population (%)
10.7 16.6 25 150 30 30 30
NS NS
103 p=3.1E-05
15 100 20 20 p=0.003 20
* *
5 50 10 10 10 p=0.03
0
I 58.3 IV 14.4 *
0 0 0 0 0
0 103 104 105 WT KO WT KO WT KO WT KO WT KO
KO 104 II III I 30 6
12.2 5.51
CD4+CD69hi (x106)
CD8+CD69hi (x106)
103 p=0.01
20 4
*
CD69
0
10 2 p=0.01
I 81.8 IV 0.76 *
0 103 104 105
0 0
TCR WT KO WT KO
Figure 2. Development of CD4+ and CD8+ SP thymocytes is impaired in Nsrp1f/f CD4Cre mice. (A) Western blot of NSrp70 expression in thymocyte
subsets from Nsrp1f/f (WT) and Nsrp1f/f CD4Cre (cKO) mice. DN, double negative; DP, double positive; SP, single positive; WT, Nsrp1f/f mouse; KO,
Nsrp1f/f CD4Cre mouse. The bar graphs indicate the mean ± standard deviation (SD) of the indicated protein blot densitometry presented with respect
to -actin. (B and C) Staining for CD4 (green) and CD8 (red) in thymus (B) and in spleen and lymph node (C) from 6-week-old WT and KO mice.
White arrowheads: CD8+ SP thymocytes in medulla. M, medulla; C, cortex. Original magnification, ×10. The fluorescent intensity of CD4+ and CD8+
thymocytes was quantified (C, bottom). (D) Flow cytometric analysis of CD4 and CD8 on thymocytes. (E) Quantification of average percent (top) and
cell numbers (bottom) of DN, DP, CD4+ or CD8+ SP thymocyte subpopulations. Each black and white circle in graphs of cell numbers represents an
individual mouse. The bar graphs (top) and small horizontal lines (bottom) indicate the mean ± SD. *, meaningful P-value; NS, non-significant P-value.
(F) Surface staining of TCR and CD69 on total thymocytes. (G–I) Quantification of TCRhi thymocytes population gated on III and IV (G), total cell
numbers gated on I–IV gate plots (H), and CD4+ CD69hi and CD8+ CD69hi cells (I) from panel (F). The bar graphs indicate the mean ± SD of populations.
hi, high expression. Each black and white circle in graphs represents an individual mouse. The small horizontal lines indicate the mean ± SD. All data
shown are representative of three independent experiments.Nucleic Acids Research, 2021, Vol. 49, No. 10 5767
mental stages based on the expression of the maturation tive splicing in the spliceosome of nuclear speckles (3,19).
marker TCR and the positive selection marker CD69. Al- NSrp70-regulated splicing targets were heavily connected
though the population and numbers of TCR-lo CD69lo with mRNA processing and splicing interaction networks,
cells (Gate I: mostly DN and preselected DP cells) and as judged by the Search Tool for the Retrieval of In-
TCRint CD69int cells (Gate II: thymocytes undergoing se- teracting Genes/Proteins (STRING) (Figure 3F). Intrigu-
lection) were unchanged, the population and numbers of ingly, NSrp70 targets are enriched with well-known cell
TCRhi CD69hi cells (Gate III: post-positive selection thy- cycle-related splicing factors, such as Srsf1, Srsf7 and Son
mocytes) and TCRhi CD69lo cells (Gate IV: mature SP (16,17,28), and spliceosome-related factors, such as Sf3b1
cells ready for export to the periphery) were significantly re- and U2af2 which are important for splice site recognition
duced in NSrp70-cKO mice (Figure 2F – H). Accordingly, (18). The read tracks of Srsf1 and Srsf7 and other splicing
the numbers of CD4+ CD69hi and CD8+ CD69hi thymocytes target genes are shown as examples (Figure 3G and H).
were reduced in NSrp70-cKO mice as compared to those
in control mice (Figure 2I). As a result, mature (CD24lo )
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
TCRhi CD4+ or TCRhi CD8+ SP thymocytes were signif- NSrp70 sequesters pre-mRNA splicing factors, including
icantly reduced (Supplementary Figure S2A and B). Taken SRSF1, into the nuclear speckles and counteracts SRSF1
together, these results indicate that NSrp70 deletion results Interestingly, we previously demonstrated that NSrp70
in severe defects in the late development of DP thymocytes physically interacts with SRSF1 and SRSF2 (3). In the
during positive selection. However, despite such a marked present study, we further corroborated the binding of
reduction in SP thymocytes, the reason for the small change NSrp70 to SRSF1, but not to histone deacetylase 1
in the size (Supplementary Figure S1C) and cellularity (Fig- (HDAC1) (29), a transcriptional regulator known to be
ure 2E) of the thymus is probably due to the fact that CD4 involved in T cell development (Figure 4A). In addition,
and CD8 SP thymocytes represent only approximately 10% NSrp70 directly binds to the mRNA of Srsf1 and Srsf7,
of the total thymocytes in the thymus. as determined by an RNA immunoprecipitation (RIP) as-
say (Figure 4B). In a previous report, we identified that the
protein-protein interaction site of NSrp70 is the RS1 re-
NSrp70 regulates alternative splicing (AS) events of pre-
gion, as shown in the schematic cartoon in Figure 4C (3,19).
mRNA splicing factors in DP thymocytes
Deletion of the RS1 region (RS1M) caused complete loss
Previously, we reported that NSrp70 modulates the alter- of NSrp70 localization in the nuclear speckles and showed
nate splice site selection of several splicing reporter mini- a disturbed pattern in the nucleus (3,19). Interestingly, co-
genes (3). To gain further insights into NSrp70-regulated transfection of RS1M caused failure of speckle localization
AS events related with its physiological functions, we per- of SRSF1 (Figure 4A). Disturbed nuclear speckle localiza-
formed high-throughput sequencing of mRNA (mRNA- tion is also observed with other NSrp70-binding proteins,
seq) with deep sequencing coverage (∼60 million, 100-nt such as SON, U2AF1, and hnRNP U, but not with U170K,
paired-end reads) after sorting of DP thymocytes from a nuclear speckle protein that does not bind NSrp70 (Sup-
WT and NSrp70-cKO mice. Strikingly, we observed that plementary Figure S3). Since spatiotemporal subnuclear or-
the number of isoforms and genes were markedly reduced ganization of the pre-mRNA processing machinery is essen-
(6.13% and 5.5% reductions, respectively; Wilcoxon rank- tial for the proper maturation of protein-coding mRNAs
sum tests, one-sided, P-values ≤ 0.05) in DP thymocytes (1), these results suggest that NSrp70 is associated with the
from NSrp70-cKO mice (Figure 3A), implying remarkable expression and function of other pre-mRNA splicing fac-
changes in the alternative splicing landscape. For a better tors, they leading to altered expression of many downstream
characterization of the changes in the AS events, we ana- genes involved in T cell development (Figure 4C).
lyzed AS changes of individual transcripts using mixture- Among various NSrp70-regulated target splicing factors
of-isoforms (MISO) models (20) and identified each 400, (Figure 3E and F), SRSF1 is particularly interesting, be-
163 and 321 NSrp70-regulated AS events with an obvious cause it is a well characterized oncogenic factor that pro-
change in PSI values (PSI ≥ 0.2) (Figure 3B). Of these, 160 motes tumorigenesis through multiple pathways (29). In
genes that commonly appeared in two or more sets were addition, interestingly, retention of intron (RI) is most of-
selected (Figure 3B). The alternatively spliced genes are ten associated with down-regulation of gene expression via
listed in Supplementary Table S2. We observed 473 NSrp70- non-sense-mediated decay (NMD) (30,31). Since the RI
regulated AS events, including exon skipping (ES), alterna- form of Srsf1 mRNA was significantly reduced in NSrp70-
tive 3 /5 splice sites (A3SS/A5SS), mutually exclusive ex- cKO DP thymocytes in the present study (Figures 3G
ons (MXE), and retained intron (RI) (Figure 3C). Subse- and 4D), we expected that Srsf1 mRNA and its canon-
quent analysis revealed that most AS events were negatively ical protein isoform would be increased in NSrp70-cKO
regulated in NSrp70-cKO DP thymocytes (decreased PSI DP thymocytes. As expected, the increased Srsf1 mRNA
value by NSrp70 KO) (Figure 3D). and protein were seen in NSrp70-cKO DP thymocytes
When analyzing the cellular functions of NSrp70- (Figure 4E).
regulated AS events using gene ontology (GO), we found Since SRSF1 is an oncogenic factor, we further inves-
that NSrp70 affects genes in the mRNA processing and tigated whether NSrp70 has an opposite role to SRSF1
splicing, DNA repair, and T cell differentiation pathways in cells. Overexpression of NSrp70 suppressed SRSF1-
(Figure 3E). These results are direct evidence that NSrp70 mediated alternative splicing of the Fas minigene (Figure
is an RNA binding factor known to regulate alterna- 4F). In addition, colony formation assay revealed that over-5768 Nucleic Acids Research, 2021, Vol. 49, No. 10
A B 1
C
0 50 100 150 200
Number of present isoforms
Number of present genes
14.5k
265 ES
20.5k
A3SS
20.0k 14.2k 12 95
28 A5SS
14.0k
19.5k 98 25 173
MXE
13.8k 2 3
19.0k RI
13.5k
I - 0.2 and BF >
I> - 10
WT KO WT KO =PSI
D F
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
PSI increased mRNA processing & splicing
PSI decreased
Hnrnph1
p=3E-05 p=3E-08 DNA repair Sugp2
p=7E-05 *
100 *
p=4E-04 * Nabp1 Cdk9 Ptbp1 Rbm39
Percent (%)
75 * Srsf7
Mum1
Ddb2
50
Ercc5 Acin1
Pms2 Sf3b1
25 Son
Mms19 Xab2
0 Tra2b Wdr83
ES A3SS A5SS RI Srsf1
E p-value T cell differentation Ddx39
0 1E-2 1E-4 1E-6 1E-8 in thymusRorc
Jmjd6 Tia1
U2af2
mRNA processing
Ncaph2
mRNA splicing
mRNA splicing via spliceosome
via spliceosome
DNA repair Srsf1
G
600 593
WT 25
T cell differentation = 0.36
RPKM
in thymus 400 17 [0.32, 0.40]
200 8
0
0 0 0.5 1
H MISO
ES A3SS 600 KO 35
RPKM
400 404 = 0.09
23
[0.07, 0.12]
Tia1 Hnrnph1 Rbm39 Cdk9 Rorc 200 11
BF=1E+12 BF=1E+12 BF=1E+12 BF=1E+12 BF=1E+12 0 0
100 0 0.5 1
60 60 60 60
MISO
*100 (%)
80
40 60 40 40 40
Srsf7
40
20 20 20 20
20
0 0 0 0 0 600 WT
WT KO WT KO WT KO WT KO WT KO 404 25
250
RPKM
400 = 0.61
17 [0.56, 0.65]
200 8
A5SS RI 0 0
0 0.5 1
198 MISO
Sugp2 Ercc5 Sf3b1 Ddx39 Ptbp1 600 KO
RPKM
BF=6.8E+11 BF=110 BF=1E+12 BF=1E+12 BF=1E+12 400 130 28 = 0.23
48 [0.20, 0.27]
50 40 60 60 60 18
*100 (%)
200
40 30 9
30 40 40 40 0 0
20
20 262 0 0.5 1
10 20 20 20 MISO
10
0 0 0 0 0
WT KO WT KO WT KO WT KO WT KO
Figure 3. Global alternative RNA splicing changes in NSrp70-cKO DP thymocytes. (A) The number of isoforms and genes present in the given samples.
We checked the number of isoforms and genes present in given wild-type and KO samples and found significant reductions of the numbers of isoforms and
genes in DP thymocytes (6.13% and 5.5% reductions, respectively; Wilcoxon rank sum tests, one-sided, P-values ≤ 0.05). (B) Venn diagram of the number
of significantly alternative spliced-genes from three mRNA-seq datasets of Nsrp1f/f (WT) and Nsrp1f/f CD4Cre (KO) DP thymocytes. PSI, Percent spliced
in; BF, Bayes’ factor. (C) Quantification of the different alternative splicing events affected by NSrp70. (D) The relative percent of each alternative splicing
event affected by NSrp70. The small horizontal lines indicate the mean ± standard deviation *, meaningful P-value. (E, F) Gene ontology analysis (E) and
functional association network (STRING) (F) of alternative spliced-targets affected by NSrp70-cKO. Fisher exact P-values were plotted for each enriched
functional category. (G) Sashimi plot of alternative exons affected by NSrp70. Genes were chosen to represent both an increase and a decrease of PSI, and
the numbers of exon junction reads are indicated. (H) PSI and BF values of different types of NSrp70-regulated alternative splicing events based on MISO
analysis.Nucleic Acids Research, 2021, Vol. 49, No. 10 5769
A B
GFP: - + - -
GFP: + - GFP: + - NSrp70 : SRSF1 NSrp70 G_NSrp70: + - + -
G_NSrp70: - + G_NSrp70: - + M WT Mutant HDAC1 G_RS1M: - - - +
70 NSrp70 RS1M NSrp70 Srsf1
SRSF1 HDAC1 (309 bp)
GFP
55 Srsf7
IP: GFP IP: GFP (335 bp)
WB: SRSF1 WB: HDAC1 Hdac1
1 m 1 m (300 bp)
Abs: IgG -GFP
SRSF1 SRSF1 HDAC1
mCherry
Total lysates
G_ 130 Srsf1
G_ ( 309 bp)
NSrp70 NSrp70 100
PCR
Srsf7
70 (335 bp)
SRSF1 HDAC1
55 Hdac1
(300 bp)
Merged
GFP GFP M
25
G_NSrp70 100
55
-actin
WB
-actin
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
GFP 25
35
Total lysates Total lysates -actin 55
35
C
NSrp70 WT and RS1M (
Coiled-coil RS-like NLS
1 289 379 558 WT
GFP RS1
SRSFs 1. Speckle organization
2. RNA alternative splicing Cell cycle and survival?
RI WT < KO
Deletion
Srsf1 mRNA exon’ exon’’ 5’ exon’ exon’’ 3’ or mutation
5’ 3’
(Cell cycle increase?)
D E
DP NSrp70
Srsf1 Hdac1
2.5 WT KO M
WT KO Intron 3
mRNA to GAPDH
p=3E-04 2.0
Relative target
2.0
Long SRSF1 35
Srsf1
(490bp) *
1.5 NS
Short 1.0 70
(284bp) 1.0 HDAC1
55
Gapdh 0.5 55
-actin
0 0.0 35
Normalized 1 1.6 WT KO
S/L 0.2 * WT KO
F Fas miniG G EO771
Colony formation: EO771
GFP O O GFP (G) O
mCherry O O EV NSrp70 EV SRSF1 mCherry (mC) O
G_NSrp70 O O G_NSrp70 O
mC_SRSF1 O O mC_SRSF1 O
567
G_NSrp70 Endo
57
Normalized 100 94 23 35
5-6-7/5-7 (%) 3 4* 5 *
GFP
Gapdh p=0.0007
Colony number
240
Colony number
450
p=2.4E-07 *
mC_SRSF1
G_NSrp70 160 * 300 Endo
mC_SRSF1 80 150 mCherry
0 0 -actin
-actin
0
EV
EV
1
p7
SF
Sr
SR
N
Figure 4. NSrp70 controls the expression of SRSF1 and counteracts mutually. (A) Immunoprecipitation (left) and immunofluorescence images (right) of
NSrp70 and SRSF1. HEK293T cells were transfected with GFP (empty vector), GFP NSrp70 (G NSrp70), or RS1M mutant. In some cases, mCherry-
fused SRSF1 and HDAC1 were co-transfected. Samples were immunoprecipitated and blotted with antibodies against the indicated proteins (left). IP,
immunoprecipitation; WB, western blotting; M, molecular mass (kDa). Fluorescence signals were visualized under a confocal microscope (right). Magnifi-
cation, 100×. Results are representative of three independent experiments. (B) RIP assay for validation of NSrp70-binding mRNA. The samples in (A) were
immunoprecipitated with the indicated antibodies (anti-rabbit IgG or anti-GFP). RNAs purified from the IP samples and total lysates were determined
by RT-PCR. bp, base pair. (C) Schematic diagram of wild-type NSrp70 and RS1M mutant. Potential regulation of alternative splicing and speckle organi-
zation by NSrp70 and its effect on T cell development. NLS, nuclear localization sequence. (D) RT-PCR analysis of different SRSF1 isoforms in Nsrp1f/f
(WT) and Nsrp1f/f CD4Cre (KO) DP thymocyte. (E) Real-time quantitative PCR (left) and western blot analysis (right) for SRSF1 and HDAC1 from WT
and KO DP thymocytes. Gapdh and -actin were shown as the loading controls. *, meaningful P-value. (F) NSrp70 counteracts SRSF1 in vivo splicing
assay. HEK293T cells were co-transfected with the indicated constructs and Fas minigene. Exon inclusion or exclusion was determined by RT-PCR (top).
The ratio of exclusion or inclusion of Fas exon 6 is shown as the normalized ratio (%) (middle). mCherry, empty vector; mC SRSF1, mCherry SRSF1.
(G) NSrp70 effects on the proliferation of EO771 cancer cells. The cells were stably transfected with NSrp70, SRSF1, or a control vector and analyzed
by colony formation assays with mean ± standard deviation of relative colony numbers plotted (left). Expression of NSrp70 and SRSF1 were confirmed
(right). All data shown are representative of three independent experiments. *, meaningful P-value.5770 Nucleic Acids Research, 2021, Vol. 49, No. 10
expression of NSrp70 significantly reduced colony forma- WT (Figure 6B), demonstrating an active cell proliferation
tion in E0771 breast cancer cells, whereas SRSF1 increased at the CD69+ DP stage. Higher numbers of Ki-67-positive
it (Figure 4G). cells were also seen in NSrp70-cKO CD69– DP thymocytes
(Figure 6B), demonstrating that loss of NSrp70 in thymo-
Aberrant gene expression related to the cell cycle, translation, cytes triggers uncontrolled cell proliferation. Interestingly,
and subcellular locations associated with alternative splicing large populations of NSrp70-cKO CD69+ DP thymocytes
in NSrp70-cKO DP thymocytes underwent apoptotic cell death (Figure 6C). This event was
accompanied by reduced expression of p21 (Figure 6D),
We subsequently analyzed how NSrp70-cKO affects global
which inhibits cell cycle progression and apoptosis (33), and
gene expression. Based on hierarchical clustering of gene
Bcl-2 and Mcl-1 (Figure 6D), which play a role in survival
expressions, we found distinct clusters of up- or down-
and apoptosis (34).
regulated expressions in the Nsrp70-cKO group (Fig-
Knockdown of NSrp70 also showed upregulation of can-
ure 5A; genes with more than 2-fold changes were
didate cell cycle regulators in non-immune HEK293T cells
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
shown). Based on the R DESeq2 package, we identi-
(Supplementary Figure S4A), suggesting that the role of
fied 834 significantly up-regulated genes and 295 down-
NSrp70 generally contributes to the expression of cell cycle
regulated genes (adjusted P-value < 0.05) (Supplementary
regulators in cells. In contrast, NSrp70-cKO did not affect
Table S3) (32).
the expression of genes known to be associated with specific
Interestingly, based on GO enrichment tests of significant
stages of T cell development, as corroborated by RT-PCR
up- or down-regulated genes (Figure 5B and Supplemen-
(Supplementary Figure S4B and C). Collectively, these re-
tary Table S4), we found that biological processes related
sults demonstrate that NSrp70 is a critical regulator of cell
to cell proliferations (e.g. cell cycle) and the synthesis of
cycle arrest and survival during TCR-mediated positive se-
cellular building blocks and energy (e.g. translation, DNA
lection.
replication and repair, and mitochondrial metabolism) were
significantly up-regulated, whereas transcription, protein
phosphorylation, T cell receptor signaling pathway, and the Deletion of NSrp70 results in defective survival signals fol-
negative regulation of apoptotic signaling were significantly lowing TCR activation in CD69+ DP thymocytes
down-regulated (Figure 5C and Supplementary Table S4;
In the mRNA-seq analysis, we found that genes involved
P-value < 0.01). Furthermore, we also identified subcel-
in the TCR signaling pathway are markedly downregu-
lular locations significantly enriched among up- or down-
lated, as depicted in the KEGG pathway (Figure 7A). A
regulated genes (Figure 5C and Supplementary Table S5; P-
dramatic reduction of CD4+ or CD8+ SP thymocytes in
value < 0.01). For example, we found that subcellular loca-
NSrp70-cKO mice suggests that the survival signals follow-
tions for macromolecular synthesis and cell cycle processes
ing TCR activation may be impaired in DP thymocytes.
(e.g. ribosome, cytoskeleton, and centrosome) were signifi-
As shown in Figure 4C, the spatiotemporal organization of
cantly up-regulated, whereas major subcellular locations as-
pre-mRNA splicing factors and their normal protein iso-
sociated alternative splicing, such as nuclear chromatin and
forms may be critical to optimally express molecules re-
nuclear speckle were down-regulated.
quired for T cell differentiation. An NSrp70 defect can in-
Quantitative RT-PCR confirmed the results of RNA-seq
duce unbalanced, polarized expression of certain protein
analysis and validated the increased expression of Cdk1,
isotypes, which may be coupled with the low expression of
Cdca3, Rpl26, Uba52, Atp5pd and Ndufs5, which are re-
survival factors such as Bcl-2, p21, and Mcl-1 (Figure 6D).
lated to the cell cycle, translation, and metabolism, and the
Interestingly, the expression of TCR, CD3ε, and CD3 -
decreased expression of Fyn, Plcg1, Ccr7, Junb, Ptk2b and
chain was significantly reduced in NSrp70-cKO CD69+ DP
Bag6, which are related to TCR signal transduction and
thymocytes (Figure 7B). Attenuated TCR signaling, which
anti-apoptosis (Figure 5D). Taken together, these results
is essential for thymocyte survival and differentiation, may
strongly suggest that NSrp70 function is closely linked to
result in retarded differentiation of SP thymocytes by cou-
cell cycle progression and building block synthesis during
pling enhanced cell growth and reduced cell survival in
the transition of DP to SP thymocytes.
NSrp70-deficient thymocytes, as schematically outlined in
Figure 7C. To test this concept, we determined the Ca2+ flux
Deletion of NSrp70 induces uncontrolled cell proliferation
induced by anti-CD3/CD28 or phorbol ester/ionomycin
followed by apoptotic cell death
(P/I). Although the Ca2+ flux evoked by P/I was simi-
Next, we investigated whether the increased expression of lar in both cKO and WT cells, the Ca2+ flux evoked by
cell cycle regulators in NSrp70-cKO DP thymocytes is re- anti-CD3/CD28 was significantly reduced in NSrp70-cKO
lated to increased cell cycle activity in vivo. To this end, CD69+ DP thymocytes, but not in CD69– DC thymocytes
total DP thymocytes were gated by CD69– (before posi- (Figure 7D). Accordingly, TCR down-stream signals were
tive selection) and CD69+ DP (post-positive selection), and clearly attenuated (Figure 7E).
then the cells were assessed for cell cycle activity and apop- After the initiation of positive selection, DP thymocytes
totic cell death. Most CD69– DP thymocytes showed G0 /G1 down-modulate CD4 and CD8 expression on the surface,
arrest of the cell cycle (Figure 6A). In contrast, NSrp70- entering the CD4+ CD8int transitional stage before commit-
cKO CD69+ DP thymocytes showed increased populations ting to the CD4+ or CD8+ SP lineage (35,36). Therefore, we
of S and G2 /M phases as compared to WT (Figure 6A). performed a two-stage differentiation assay in vitro (25) and
Consistently, NSrp70-deficient CD69+ DP thymocytes con- found that only small numbers of NSrp70-cKO DP thymo-
tained higher numbers of Ki-67-positive cells than that of cytes were entering into the CD4+ CD8int stage. As a result,Nucleic Acids Research, 2021, Vol. 49, No. 10 5771
A WT KO B
1 2 3 1 2 3
Translation Covalent
chromatic modification
Cell cycle DNA-templated
transcription
Cell division Protein phosphorylation
Genes satisfying with fc2 = 834 Antigen processing Phosphorylation
Mitotic Regulation
nuclear division of transcription
0 20 40 60 0 1 2 3 4 5 6 7
-log10 p-value -log10 p-value
C ion
Mitochondrial Protein folding ein lat
metabolism & localization ot ry Response to cAMP
Pr pho
os
ph
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
Mitophagy T cell receptor
signaling
Mitotic spindle
14
Normalized Expression
& chromosome
Transcription
Cell cycle negative regulation of
7
apoptotic signaling
Splicing
DNA replication
& repair
Translation
0
Transport &
Biological processes Proteolysis metabolism
Spliceosome Proteosome
Ribosome Cell projection Vesicle
Centrosome Growth cone
Nuclear speckle
Plasma membrane
Nuclear
Mitochondria chromatin
Cytoskeleton
Protein
complex Extracellular
exosome Nucleus
Subcellular locations
D Cell cycle Ribosome Oxidative phosphorylation
Cdk1 Cdca3 Rpl26 Uba52 Atp5pd Ndufs5
Relative mRNA level
Relative mRNA level
2.5 2.5 p=2.4E-05 2.5 2.5 p=2E-03 2.5 2.5
p=4.4E-04 p=9E-03 p=2E-02 p=3E-03
to GAPDH
to GAPDH
2 2 2 2 2 2
* * *
1.5 * 1.5 1.5 * 1.5 1.5 * 1.5
1 1 1 1 1 1
0.5 0.5 0.5 0.5 0.5 0.5
0 0 0 0 0 0
WT KO WT KO WT KO WT KO WT KO WT KO
TCR & signal transduction Cell cycle control (apoptosis)
Relative mRNA level
Relative mRNA level
Fyn Plcg1 Ccr7 Junb Ptk2b Bag6
p=4E-02 p=2E-03 p=4E-04 p=4E-03 p=2E-05 p=6E-06
to GAPDH
to GAPDH
1 1 1 1 1 1
0.5 * 0.5 * 0.5 * 0.5 * 0.5 * 0.5 *
0 0 0 0 0 0
WT KO WT KO WT KO WT KO WT KO WT KO
Figure 5. Aberrant gene expression related to the cell cycle and TCR intracellular signal transduction in NSrp70-cKO DP thymocytes. (A) Hierarchical
clustering of gene expressions with more than two-fold changes. We found distinct gene clusters of up- or down-regulation in Nsrp70-cKO DP thymocytes.
The heat-map for gene expression patterns was generated using the Multi Experiment Viewer (MeV) software. Genes with red and blue colors indicate
higher and lower expressions, respectively. fc2, 2-fold change. (B) Top-5 most significantly up-regulated or down-regulated biological process terms from
significantly differentially expressed genes (negative binomial tests) based on gene ontology enrichment tests (modified Fisher’s exact tests from DAVID).
(C) The global map of significantly up-regulated (pink) or down-regulated (skyblue) gene ontology terms (biological processes and subcellular locations,
top and bottom, respectively) by gene ontology enrichment tests (P-value < 0.01). Like in the Enrichment Map (54), we clustered enriched biological
processes and subcellular locations as connected in the map if they shared genes in significant numbers (Jaccard > 0.25). (D) Real-time quantitative PCR
analysis for the validation of NSrp70-regulated targets. mGapdh was used as the loading control. *, meaningful P-value. All data shown are representative
of three independent experiments.5772 Nucleic Acids Research, 2021, Vol. 49, No. 10
A CD69- DP CD69+ DP
100 p=4E-05
10 4
WT * G0/G1
600
G0/G1: 98.8 G0/G1: 69.2
80 S
103
400
S: 0.22 S: 8.24
CD8
G2/M: 0.8 G2/M: 22.3 G2/M
Cells (%)
0 200 60 p=1E-06
*
0 3
10 10 4
10 5 0
CD4 40 p=3E-04
CD69- DP CD69+ DP KO
Cell counts
2.0K 600
G0/G1: 96.7 G0/G1: 59.8 *
Counts
1.5K 400 S: 0.62 S: 9.98 20
1.0K G2/M: 2.4 G2/M: 29.6
Downloaded from https://academic.oup.com/nar/article/49/10/5760/6284184 by guest on 05 November 2021
200
500 0
WT
KO
WT
KO
WT
KO
0 0
0 103 0 1.0K 2.0K 3.0K 1.0K 2.0K 3.0K
CD69+ DP
CD69 7-AAD
B C D
CD69- DP CD69+ DP CD69- DP CD69+ DP WT KO WT KO
WT WT 0.36 17.8
Bcl-2 CDK2
103
Bcl-2 family
Bcl-xL CDK4
55.9 53.3
102
0 Bim Cyclin A
93.7 5.94 75.2 7.02
Ki-67
KO KO 0.50 30.8 Mcl-1 p21
7-AAD
3
10
103 cIAP1 p27
IAP family
72.4 78.7
102 Actin
102 cIAP2
0
0
90.4 9.14 61.3 7.86
0 1K 2K 3K 0 1K 2K 3K 0 103 XIAP
Annexin V
7-AAD
50 Apoptotic cells p=2E-05
Dead cells * 1.5 WT KO
1200 p=2E-06
protein to -actin
p=2E-04
Population (%)
p=1E-04
p=5E-04
40
Relative target
Ki-67 (MFI*10)
p=3E-05
1000 * NS NS NS NS NS
30 1 p=4E-04
p=2E-06
800 p=5E-03
* * *
p=2E-06
600 * 20 *
NS 0.5 *
400
10 *
200 *
0 0
0
WT KO WT KO WT KO WT KO l-2 -xL im l-1 P1 P2 AP K2 K4 inA 21 27
Bc Bcl B Mc cIA cIA XI CD CD ycl p p
CD69- DP CD69+ DP CD69- DP CD69+ DP C
Figure 6. Deletion of NSrp70 induces uncontrolled cell proliferation followed by apoptotic cell death. (A) Cell cycle analysis of DP thymocytes from
Nsrp1f/f (WT) and Nsrp1f/f CD4Cre (KO) mice. Populations of CD69– or CD69+ thymocytes were gated from CD4 and CD8 DP thymocytes (left) and
were analyzed for DNA content by 7-ADD intensity (right). The bar graphs indicate average ± SD of different cell cycle stages population. *, meaningful
P-value. (B, C) Analysis of cell proliferation by Ki-67 staining and apoptotic cell death by annexin V and 7-ADD. DP thymocytes from (A) were stained
with anti-Ki-67 antibody and 7-ADD (B) or annexin V and 7-ADD (C) and analyzed by flow cytometry. Bar graphs indicate mean fluorescence intensities
(MFI) (B, bottom). Annexin V+ populations represent early apoptotic cells and annexin V+ and 7-ADD+ populations represent dead cells (C). The bar
graphs indicate average ± SD of apoptotic and dead thymocyte populations. NS, non-significant P-value. (D) Western blot of cell cycle and cell death-
related proteins in samples extracted from WT and KO thymocytes. -actin served as a loading control. The bar graphs indicate average ± SD of indicated
protein blot densitometry presented relative to -actin. All data shown are representative of three independent experiments.
the populations of CD4+ CD8int were significantly lower in Since rapid proliferation is coupled with impaired SP
NSrp70-cKO cells than in WT cells (Figure 7F). Similar to maturation in NSrp70-deficient CD69+ DP thymocytes, we
the in vivo condition, NSrp70-cKO CD69+ DP thymocytes next questioned whether blocking of the cell cycle may
showed a higher proliferative phenotype (Ki-67+ cells) than overcome the developmental defects. To determine this,
that of WT cells (Figure 7G). In addition to the prolifer- NSrp70-cKO DP thymocytes were treated with the CDK1
ative phenotype, these cells underwent increased apoptotic inhibitor purvalanol A and a two-stage differentiation as-
cell death, presumably due to the reduced expression of sur- say was performed. We found that the NSrp70-cKO DP
vival factors (Figure 7H). thymocytes treated with purvalanol A did not differenti-You can also read

















































