Long-term fruit fly monitoring and impact of the systems approach on richness and abundance
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Canadian Entomologist (2021), page 1 of 20 Published on
behalf of the
doi:10.4039/tce.2021.32 Entomological
Society of
Canada
ARTICLE
Long-term fruit fly monitoring and impact of the
systems approach on richness and abundance
Mayara R. de Araujo1 , David dos S. Martins2 , Maurício J. Fornazier2 , Keiko Uramoto1 ,
Paulo S.F. Ferreira3 , Roberto A. Zucchi1 , and Wesley A.C. Godoy1*
1
Departamento de Entomologia e Acarologia, Universidade de São Paulo, Escola Superior de Agricultura Luiz de Queiroz
(ESALQ), 13418-900 Piracicaba, São Paulo, Brazil, 2Departamento de Entomologia, Instituto Capixaba de Pesquisa,
Assistência Técnica e Extensão Rural (INCAPER), Caixa Postal 01146, 29001-970, Vitória, Espírito Santo, Brazil, and
3
Departamento de Entomologia, Universidade Federal de Viçosa (UFV), Avenida Peter Henry Rolfs s/n, Campus
Universitário, 36570-900, Viçosa, Minas Gerais, Brazil
*Corresponding author. Email: wacgodoy@usp.br
(Received 25 October 2020; accepted 12 April 2021; first published online 19 July 2021)
Abstract
We investigated ecological patterns of richness and abundance of Anastrepha fruit flies, based on a
long-term series with a dataset from 1998 to 2010, subdivided into four subseries describing pest
management through the systems approach (integration of different measures, at least two of which
act independently, with cumulative effects), and its impact on abundance and diversity of fruit
flies. Richness and abundance were influenced by time and distance but to different extents.
Spatio-temporal analysis taking into account the implementation of the systems approach revealed
clear effects of the pest management on fruit fly richness and abundance. However, abundance was
affected by the systems approach three years before richness was. Abundance and richness also
showed different relationships with time and distance between orchards and forest. The Gompertz
model, used to describe the relationship between area and species richness, was the function that
showed the best fit to the data. The richness-partitioning analysis, which decomposes beta diversity,
indicated different distributions of richness values and predictions for additive partitioning that were
directly associated with the implementation of the systems approach. The spectral analysis projected
different trends for peaks, indicating that the systems approach is able to delay the time for new
population peaks of fruit flies.
Introduction
Papaya (Carica papaya Linnaeus) is an important tropical fruit marketed worldwide, with most
production occurring in a few countries – in particular, India and Brazil (Pathak et al. 2019).
Although Brazil has extensive planted areas and a favourable climate for the year-round
production of papaya, phytosanitary problems, including the occurrence of fruit flies (Diptera:
Tephritidae), can limit the export of fruits. Among the main species of fruit fly in Brazil,
121 members of the genus Anastrepha that are native to the Americas occur in the country
(Zucchi and Moraes 2021). Of note among the pests causing serious quarantine-related
problems for papaya exports are Anastrepha fraterculus (Wiedemann, 1830), a native
polyphagous species, and Ceratitis capitata (Wiedemann, 1824), an exotic species recorded in
Brazil in 1901 (Martins et al. 1993; Martins and Fornazier 2014). A successful management
programme using a systems approach was implemented in Brazil in the 1990s to enable the
Subject editor: Julia Mlynarek
© The Author(s), 2021. Published by Cambridge University Press on behalf of the Entomological Society of Canada
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.322 Araujo et al.
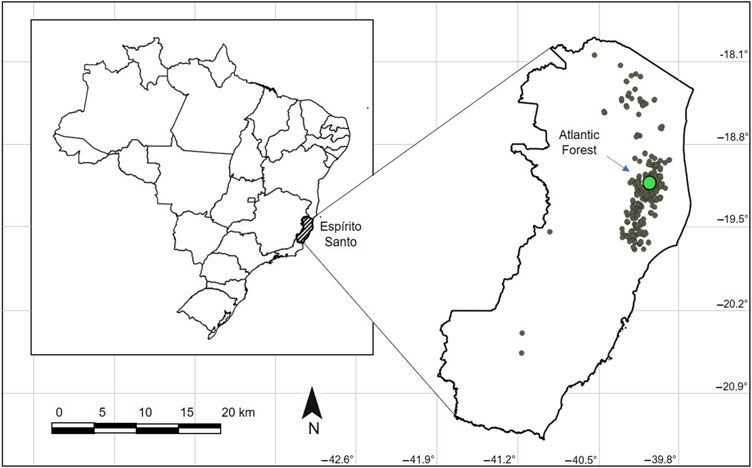
Fig. 1. Maps of fruit fly collection sites in the state of Espirito Santo, Brazil, 1998–2010.
export of papaya to the United States of America. The systems approach involves the integration
of actions during the pre- and postharvest periods to improve production, harvesting, packaging,
and transport of a commodity that meets quarantine-security requirements (Martins et al. 2012).
Essential to this type of programme is the monitoring of fruit fly abundance and richness.
Knowledge of the abundance of fruit flies and of the distribution patterns of species richness is
essential for pest management programmes because analysis of this information allows the
pest-infestation period to be evaluated, critical areas for planting to be identified, and
scenarios for implementation of control measures by harvest management, by release of
control agents, and by management and conservation of the diversity of natural enemies of
the flies, to be developed (Lundgren and Fausti 2015).
In this study, we investigated the patterns of richness and abundance of Anastrepha, analysed
through time, space – expressed by the distance of papaya orchards from an Atlantic forest
remnant – and area size. The patterns of richness and abundance of the fruit flies were also
analysed based on meteorological data. The analyses were based on a full time series (1998–2010)
and four partial series within the full series to investigate the impact of the systems approach
programme on fruit fly richness and abundance in commercial papaya production fields.
Materials and methods
Dataset
The dataset used in this study is part of a project entitled “Survey of fruit flies in the state of
Espirito Santo, Brazil”, coordinated by the Instituto Capixaba de Pesquisa, Assistência Técnica e
Extensão Rural. Data were obtained via weekly monitoring using one McPhail trap every two
hectares at different georeferenced points (18° 02' 35.7" to 20° 33' 58.3" S and 39° 13' 00.7" to
41° 09' 16.1" W) in papaya orchards in commercial production fields of Espirito Santo, Brazil
(Fig. 1) from 1998 to 2010. This is the main production area in the state for papaya export.
We investigated the richness and abundance of a pool of species of Anastrepha. The distance
between each orchard and the forest remnant was established, based on the same point within
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 3
the forest fragment (19° 59' 04" S and 39° 55' 24" W), a central area located in the Reserva
Natural Vale, which is a protected Atlantic forest remnant with high biodiversity within
approximately 23 000 ha. Annual mean rainfall, temperature, and relative humidity were
obtained from the Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural to
evaluate the influence of these factors on fruit fly richness and abundance.
Time series and the systems approach
The time periods consisted of a full time series of fruit flies in papaya orchards, from 1998 to
2010, and four partial time series, two of which include data from the beginning of the systems
approach implementation and the other two of which include data showing the impact
of the systems approach on fruit fly abundance and diversity. The first partial time series,
1998–2005, was analysed for fruit fly species richness. The second time series, 1998–2003, was
analysed for fruit fly abundance. The third time series, 2006–2010, was analysed for fruit fly
species richness, and the fourth partial time series, 2004–2010, was analysed for fruit fly
species abundance. In the four series, richness and abundance were analysed separately
because the systems approach affected species abundance and richness at different times. For
convenience, we named the 1998–2005 series – the first partial time series analysed and the
period during which management actions were implemented but were not yet yielding
observable effects on richness and abundance – “the continuous implementation of systems
approach”, and we named the two partial time series between 2004 and 2010 “impact on
richness” and “impact on abundance” because these periods showed visible impacts on
richness and abundance, respectively.
Data analysis
The results indicated that the relationship between richness or abundance and rainfall,
temperature, and humidity followed a normal distribution. A multiple regression then was
employed to analyse the significance of the relationships between richness or abundance and
those three environmental variables. The distribution of richness and abundance in time or at
distance between the papaya orchards and the Atlantic forest remnant did not follow a
normal distribution. We therefore used generalised linear models with Poisson errors to
analyse the relationship between fruit fly richness or abundance and time or distance between
papaya orchards and the forest remnant.
Nonlinear models fitted to the data
We used the R package “sars” to fit 19 SAR models describing convex and sigmoid shapes
(Matthews et al. 2019) aiming to find the best model shape to describe the relationship
between fruit fly richness and area, by comparing information criterion weights using a
barplot. Nonlinear and linear regressions (CRAN, version 1.1.1; R Core Team 2020) were also
used to investigate the relationship between the explanatory variables (time or distance) and
the response variables (abundance or richness) and to test the significance of the relationship
between them. The Akaike information criterion was used to compare the best fit of the
regression models to the data.
Richness-partitioning analysis
In this study, beta diversity was partitioned at three scales – area, ecoregion, and geographic
region – to understand the degree to which different area sizes could explain the changes in
richness over different time periods and to compare the impact of the systems approach on
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.324 Araujo et al.
beta diversity. Within insect communities, often only a fraction of species are capable during
dispersal of reaching regions that are more distant than others due to resource availability.
However, this distribution may not be obvious because of the intense movement of insects
among orchards, thereby requiring beta diversity – partitioning procedures to consider the
possibility of the richness being other than that directly observed via trapping. Other factors
besides area size may also significantly influence fly movement and, consequently, species
richness. For example, host plant diversity and spatial scale have been listed as causes of
richness variation in a tephritid community (Dinis et al. 2010).
The spatial scale used to analyse beta diversity can influence species-richness estimates. This
happens mainly when species diversity is linked to host richness, which would require insects to
travel more and colonise habitats at greater distances (Wagner and Edwards 2001; Kemp and
Ellis 2017). In addition, human-caused disturbances on landscapes at different scales have also
been shown to influence plant richness and tephritids (Dinis et al. 2010). The total number of
species in all areas of the present study was described as γ. The mean richness of the areas
studied was represented by ᾱ and was associated with area size. In that way, the reason why
species were missing in specific areas could be associated with area size. The term “βarea” was
an estimate of the mean difference between ᾱ and the diversity predicted for the largest area.
Generally, β contains both βarea and βreplace, with the latter term determining the mean
number of species missing due to factors other than area size (Henry and Stevens 2009). The
partitioning can then be described as β βarea βreplace , where βreplace is the mean number
of missing species that are not explained by patch size. In the present study, βreplace was
classified as βecoregion and βgeogr.region to evaluate the richness response to different spatial
scales. The parametric space “ecoregion” denotes a biogeographic regionalisation of terrestrial
biodiversity, and the parametric space “geographic region” is defined as a biodiversity
hotspot characterised by high species diversity (Reid 1998). In the context investigated in the
present study, the full partition can be viewed as γ = βarea βecoregion βgeogr.region
(Henry and Stevens 2009), where βreplace βecoregion βgeogr:region .
Time series and peaks
Periodograms were developed to evaluate the presence of potentially recurring peaks, based on
comparisons among spectrum values. Spectrum values are useful when comparing the potential
for new population peaks after a previous occurrence, based on frequencies rather than
fluctuations of numbers, thereby producing a periodogram (Crawley 2013). For specific cases
such as the dynamic series, 10-year cycles would have a frequency of 0.1 per year. The result
of the analysis is a plot-on-log scale, in units of decibels, with the frequency (x axis)
indicating the bandwidth, and 95% confidence interval in decibels shown by a vertical bar in
the top right-hand corner (Crawley 2013). The interpretation of the result indicates that,
at the maximum value where the spectrum takes place, cycles could occur. For example, if the
value is 0.1, the result would show cycles with a period of 1/0.1 = 10 years. Longer cycles for
frequency 0.05 would result in cycles of length 1/0.05 = 20 years.
Results
A total of 34 species was obtained during the 13-year series, with Anastrepha fraterculus and
Anastrepha obliqua (Macquart, 1835) showing the highest abundance, comprising 54.48% and
16% of the specimens obtained, respectively (Table 1). Among the variables (rainfall,
temperature, and relative humidity) evaluated with respect to their influence on richness and
abundance, only relative humidity showed a positive relationship with richness (t = 2.826,
P = 0.0199). Our results indicate an initial increase of richness and abundance during the
period of continuous implementation of the systems approach, followed by a decrease after
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 5
Table 1. Relative abundance of Anastrepha fruit fly species in papaya orchards in the state of Espírito Santo, Brazil, from
1998 to 2010.
Relative Relative Relative
Species abundance (%) Species abundance (%) Species abundance (%)
A. fraterculus 54.48 A antunesi 0.30 A. consobrina 0.04
A. obliqua 16.00 A. paralela 0.30 A. grandis 0.04
A. distincta 8.91 A. leptozona 0.19 A. minensis 0.04
A. serpentina 6.77 A. sororcula 0.19 A. sagittifera 0.04
A. pickeli 6.57 A. nascimentoi 0.15 A. sp. 2 0.04
A. pseudoparallela 2.08 A. bondari 0.13 A. dissimilis 0.02
A. bahiensis 1.82 A. sp. 1 0.13 A. furcata 0.02
A. barbiellinii 1.10 A. atlantica 0.09 A. lanceola 0.02
A. manihoti 0.71 A. bezzii 0.09 A. mixta 0.02
A. zenildae 0.61 A. zernyi 0.09 A. tumida 0.02
A. montei 0.41 A. amita 0.06
A. bivittata 0.37 A. quararibeae 0.06
A
1.5
Continuous implementation Impact on Richness
of Systems approach
1.0
Richness
0.5
0.0
1998 2000 2002 2004 2006 2008 2010
Year
B
Continuous implementation Impact on Richness
of Systems approach
Abundance
30
10
0
1998 2000 2002 2004 2006 2008 2010
Year
Fig. 2. A, Richness and B, abundance of Anastrepha fruit flies per hectare, 1998–2010, in papaya orchards in the state of
Espirito Santo, Brazil, based on the effect of the systems approach and its impact on the richness and abundance of flies.
2005 for richness and after 2003 for abundance – periods characterised by the impact on richness
and on abundance when observing the dispersal of points (Fig. 2A, B). Fruit fly richness and
abundance varied significantly over time from 1998 to 2010 (Table 2). Table 2 also shows the
variation in richness and abundance in response to distance between the papaya orchards and
the Atlantic forest remnant, with significant variation occurring only for abundance. The two
subseries, 1998–2005 and 1998–2003, which were analysed for richness and abundance over
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.326 Araujo et al.
Table 2. Regression with Poisson errors based on species of Anastrepha richness and abundance variation in response to
time and distance among papaya orchards and Atlantic forest remnant.
Coefficients Estimate Standard error z value Pr (> |z|)
Analysis of richness variation, 1998–2010, in response to time
Intercept 322.94 39.88 8.098 5.59e–16
Year –0.16 0.02 –8.12 4.68e–16
Analysis of abundance variation, 1998–2010, in response to time
Intercept 502.25 21.29 23.59 < 2e–16
Year –0.25 0.01 –23.57 < 2e–16
Analysis of richness variation in response to distance (1998–2010)
Intercept –0.82 0.15 –5.37 7.9e–18
Distance –0.001 0.001 –0.92 0.35
Analysis of abundance variation in response to distance (1998–2010)
Intercept 0.77 0.09 8.6 < 2e–16
Distance –0.004 0.0009 –4.73 2.17e–06
Table 3. Regression with Poisson errors based on species of Anastrepha richness and abundance variation in response to
time and distance among papaya orchards and Atlantic forest remnant.
Coefficients Estimate Standard error z value Pr (> |z|)
Analysis of richness variation, 1998–2005, in response to time
Intercept 94.63 78.28 1.2 0.23
Year –0.05 0.04 –1.21 0.23
Analysis of abundance variation, 1998–2003, in response to time
Intercept 223.5 44.97 4.96 6.82e–07
Year –0.11 0.022 –4.96 7.59e–07
Analysis of richness variation in response to distance (1998–2005)
Intercept –0.51 0.17 –3.05 0.002
Distance –0.002 0.001 –1.09 0.27
Analysis of abundance variation in response to distance (1998 to 2003)
Intercept 1.75 0.10 16.65 < 2e–16
Distance –0.009 0.001 –7.84 4.29e–15
time, respectively, indicated a significant relationship for abundance but not for richness (Table 3).
The same result was observed for the distance between papaya orchards and Atlantic forest
remnant (Table 3). The two subseries, 2006–2010 and 2004–2010, analysed for richness and
abundance, respectively, indicated a significant relationship with time for both richness and
abundance (Table 4), but with respect to distance, only abundance showed a significant
relationship (Table 4).
The information criterion weights were investigated for 19 mathematical functions describing
model curve shapes (Fig. 3). The barplot for the full series (1998–2010) with respect to species
richness per area shows the performance of the functions by comparing their respective
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 7
Table 4. Regression with Poisson errors based on species of Anastrepha richness and abundance variation in response to
time and distance among papaya orchards and Atlantic forest remnant.
Coefficients Estimate Standard error z value Pr (> |z|)
Analysis of richness variation, 2006–2010, in response to time
Intercept 994.6 168.2 5.9 3.40e–09
Year –0.49 0.008 –5.9 3.25e–09
Analysis of abundance variation, 2004–2010, in response to time
Intercept 948.2 103.63 9.15 < 2e–16
Year –0.47 0.05 –9.15 < 2e–16
Analysis of richness variation in response to distance (2006–2010)
Intercept –2.3 0.45 –5.4 6.35e–08
Distance 0.005 0.004 1.33 0.18
Analysis of abundance variation in response to distance (2004–2010)
Intercept –0.78 0.15 –5.2 2.03e–07
Distance 0.005 0.001 3.61 3e–04
A Model weights B Model weights
0.8
0.0 0.2 0.4 0.6
IC weights
IC weights
0.4
0.0
AR CR E1 Gom Lin Mon NegE P2 PowR CW3 AR CR E1 Gom Lin Mon NegE P2 PowR CW3
C Model weights
IC weights
0.10
0.00
AR CR E1 Gom Lin Mon NegE P2 PowR CW3
Fig. 3. Barplot of the IC weights of 19 different models for the time series A, 1998–2010, B, 1998–2005, and C, 2006–2010,
emphasising Anastrepha species richness per area size, with the Gompertz model (Gom) showing the highest information
criterion weight. AR, asymptotic; BP, beta-P; CR, Chapman–Richards; Loga, logarithmic; E1, extended power 1; E2, extended
power 2; Gom, Gompertz; Kob, Kobayashi; Lin, linear; Hel, logistic; Mon, monod; MMF, Morgan–Mercer–Flodin; NegE,
negative exponential; P1, Persistence function 1; P2, Persistence function 2; Pow, power; PowR, power Rosenzweig; RF,
rational; CW3, Weibull-3; CW4, Weibull-4.
information criterion weights (Fig. 3A). The Gompertz model was the function showing the
highest information criterion weight (Fig. 3A) and the lowest Akaike information criterion
(921), and the Weibull4 model showed the next highest information criterion weight, with
Akaike information criterion equalling 927. The distribution of richness values in response to
area sizes indicated that higher richness frequency was observed in areas below 50 ha. The
Gompertz function yielded 6.55 as the theoretical estimate of richness (Fig. 4).
The barplot for the first subseries, 1998–2005 (the period of continuous implementation of the
systems approach), shows the performance of the functions by comparing their respective
information criterion weights (Fig. 3B). The Gompertz model also showed the highest
information criterion weight, and its Akaike information criterion was 722, a lower value than
the asymptotic model, for which the Akaike information criterion was 727, and the Persistence
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.328 Araujo et al.
Gompertz
10
8
Species richness
6
4
2
0
0 50 100 150 200 250
Area
Fig. 4. The fit (solid line) of the Gompertz model to data for Anastrepha fruit fly richness (solid circles) in different area sizes
in the state of Espirito Santo, Brazil, 1998–2010.
Gompertz
10
8
Species richness
6
4
2
0
0 50 100 150 200 250
Area
Fig. 5. The fit (solid line) of the Gompertz model to data for Anastrepha fruit fly richness (solid circles) in areas of different
sizes in the state of Espirito Santo, Brazil, 1998–2005.
function 1, for which the Akaike information criterion was 728 (Fig. 3B). The distribution of
richness values in response to area sizes indicates that higher richness was also observed in
areas below 50 ha. The Gompertz model resulted in a theoretical richness of 6.39 (Fig. 5).
The barplot for the subseries 2006–2010 did not clearly indicate a specific model to fit the curve to
these data as observed in the previous series, although Weibull3 (CW3) showed the highest
information criterion weight (Fig. 3C). The Akaike information criterion values were close to each
other, showing the following values for the four models with the highest information criterion
weights: for Weibull3, the Akaike information criterion was –2.75; for Persistence function 1 (P1),
the Akaike information criterion was –2.69; for Weibull4, the Akaike information criterion was –
2.53; and for the Gompertz model (Gom), the Akaike information criterion was –2.34. Figure 3C
shows all functions’ fit, except that of Persistence function 1 because its model shape is convex
and not sigmoid as with Weibull3, Weibull4, and Gompertz. In addition, the Persistence function
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 9
A B
Fig. 6. A, Additive diversity partitioning to analyse the distribution of Anastrepha fruit fly species from 1998 to 2010 (full
series), in areas with different sizes, characterising different types of beta diversity, with the Gompertz curve fitted.
Predicted richness for the maximum area is 6.55. B, Partitioning from 1998 to 2005 (continuous implementation of the
systems approach), in areas with different sizes characterising different types of beta diversity, with the Gompertz
curve fitted. Predicted richness for the maximum area is 6.39.
1 exhibited theoretical richness that was less than 0.5 – a value much lower than the other models –
probably in response to its convex shape.
The diversity partitioning was investigated by analysing the distribution of fruit fly richness
according to the beta-diversity types (Fig. 6). Most of the richness values were within the
interval greater than 1 and less than 10, the β-area. The α-area contained the second-highest
concentration of richness values, followed by β-eco with a few richness values close to the line
between β-area and β-eco (Fig. 6). No richness value was observed in β-geo (Fig. 6).
Figure 6A also shows the Gompertz curve fitted to the full series, and Fig. 6B shows the same
curve fitted to the data influenced by the continuous implementation of the systems approach.
Comparing the distribution of richness values in Figures 6 and 7, a clear difference between the
figures can be observed, with the predicted richness decreasing from 6.39 to 1.57 (Weibull3), to
1.51 (Weibull4), and to 1.83 (Gompertz), and the decrease occurring from the continuous
implementation of the systems approach to the impact on richness subseries (Figs. 6B and 7).
A comparison among the series, full series (1998–2010), continuous implementation of the systems
approach (1998–2005), and impact on richness (2006–2010) also shows that the highest predicted fruit
fly richness occurred from 1998 to 2010 (Fig. 6A). However, the distribution of richness values was
almost the same for the full series and the series for the continuous implementation of the systems
approach, with most of the points in βarea (Fig. 6). A slight difference in terms of point
distribution can be observed exactly on the dotted line close to 10. In Fig. 6A, the points are
distributed above or below this line, but in Fig. 6B, a couple of points fall on the dotted line. Only
in the impact on richness series (2006–2010) were the points distributed in a narrower range, with
most of the richness values scattered between 0 and 5 for all fitted models (Fig. 7). For this
scenario, the β-area had a narrower vertical range than in the previous scenarios, and this
characteristic led to the points also being distributed in β-eco (Fig. 7).
The time-series analysis for both the full series and the two subseries, continuous
implementation of the systems approach (1998–2005) and impact on abundance (2004–2010),
indicated clear differences in terms of peaks, mainly between the continuous implementation
of the systems approach and the impact on abundance series, with respect both to peak
magnitude (fly axis) and to the time of occurrence (time axis; Fig. 8). A comparative spectral
analysis revealed a maximum spectrum value of 1295 for the full series, 2798 for the series for
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.3210 Araujo et al.
A B
C
Fig. 7. Additive diversity partitioning to analyse the distribution of fruit fly species from 2006 to 2010 (impact on richness),
in areas with different sizes, characterising different types of beta diversity, with the A, Gompertz, B, Weibull3, and
C, Weibull4 curves fitted. Predicted richness for the maximum value = 1.83 (Gompertz); 1.57 (Weibull3); 1.51 (Weibull4).
the continuous implementation of the systems approach, and a maximum spectrum value of 367
for the series for impact on abundance (Fig. 9). The difference between the three spectrum values
reflects the series analysed, and the lowest value (367) occurred for the impact on abundance
series. Comparing the potential peak production among the three scenarios, the full time
series (1998–2010) and the subseries for the continuous implementation of the systems
approach (1998–2005) would show a maximum peak with a period of 1/0.16 = 6.25 years.
However, the impact on abundance series (2004–2010) would show a maximum peak with a
period of 1/0.0008 = 125 years.
Discussion
Of the Anastrepha species obtained in the data series from 1998 to 2010, a high percentage (87%)
were previously recorded for the state of Espirito Santo (39 species; Zucchi and Moraes 2021). This
number of species was observed for the total dataset. However, for the analyses, the species richness
per area was used. The maximum value for richness per area was 10 species. Anastrepha fraterculus
and A. obliqua were the most abundant species in the dataset. Both have wide geographical
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 11
A B
C
Fig. 8. A, Full time series (1998–2010), B, continuous implementations of the systems approach subseries (1998–2005),
and C, impact on abundance (2004–2010) subseries, as times series object in R language.
distributions and are highly polyphagous. Anastrepha fraterculus is the species with the largest
number of host plants in Brazil (128), followed by A. obliqua (64; Zucchi and Moraes 2021).
Dominance of these species has been observed in several Brazilian states (Garcia et al. 2003;
Aguiar-Menezes et al. 2008; Lopes et al. 2015; Monteiro et al. 2019; Oliveira et al. 2019).
Usually, in commercial orchards one or two fruit fly species are dominant, comprising
a significant part of the total of captured specimens (Aluja et al. 1996). An array of biotic
and abiotic factors, including host plants, may simultaneously contribute to the establishment
of fruit flies as dominant and codominant species in specific areas (Aluja et al. 1996;
Rwomushana et al. 2009). In addition, both A. fraterculus and A. obliqua are considered
complexes of cryptic species sensu lato that are difficult to distinguish morphologically.
Effects of environmental variables
Our results suggest that only relative humidity influenced Anastrepha richness and that
abundance was not influenced by any of the three environmental variables analysed.
Environmental variables, particularly rainfall and soil moisture, can affect fruit fly life stages
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.3212 Araujo et al.
A B
C
Fig. 9. A, Spectral analysis for full series (1998–2010) with maximum spectrum of 1295 and frequency of 0.16, projecting a
1
period of 0:16 6:25 years for a new peak of this size to occur; B, continuous implementation of the systems approach
(1998–2005) with maximum spectrum of 2798 and frequency of 0.16, also projecting a period of 0:16 1
6:25 years; and
C, impact on abundance (2004–2010) with maximum spectrum of 367 and frequency of 0.008, projecting a period of
0:008 125 years between peaks.
1
in different ways (Montoya et al. 2008; Quesada-Moraga et al. 2012). Generally, rainfall,
temperature, and relative humidity affect both fruit fly diversity and abundance (Nascimento
et al. 1982; Garcia et al. 2003; Trassato et al. 2015; Gnanvossou et al. 2017), but the
correlation coefficients are usually low and some of them are statistically not significant. The
absence of effects of temperature and rainfall on richness and abundance found in the present
study is probably a function of a local aspect – that is, an area with orchards that are not far
from each other and are all influenced by the same conditions, resulting in apparent stability
of values during the observation period.
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 13
Effects of the systems approach
Fruit fly richness and abundance varied significantly from 1998 to 2010. The three subseries
investigated – the continuous implementation of the systems approach (1998–2005) and the
impacts on richness (2006 to 2010) and on abundance (2004–2010) – showed different
periods; the continuous implementation of the systems approach subset described an
increasing dispersal of points for both richness and abundance of flies, whereas the impact on
richness and abundance subsets showed the opposite. The high abundance and richness of
flies before 2006 probably occurred in response to the absence of pest management in
previous years. Even with the implementation of the systems approach before 2006, it takes
time for control actions to affect fruit fly abundance and species richness. However, pest
control implemented through the systems approach did result in a lower richness and
abundance of species of Anastrepha after a few years. The relationship between species
richness and abundance is generally associated with commonness and rarity, which are usually
cited as dependent on the heterogeneity of resources (Verberk 2011). In this study, fruit fly
richness reached its maximum value at 10 species per area and then decreased in response to
a set of actions, indicating successful pest management. The results analysed here were
obtained mainly from papaya orchards, although the data were likely influenced by nearby
urban and Atlantic forest remnant areas. The maximum richness found in the present study is
coherent with fruit fly surveys in tropical areas, which generally do not find richness levels
higher than those estimated in this study (Marsaro Júnior et al. 2012; Almeida et al. 2019).
The abundance of flies also changed during the study period. The most pronounced change in
the number of flies occurred after 2005, probably in part as a result of the actions taken in
the systems approach (Martins et al. 2012).
Generally, fruit flies infest papaya at the fruit’s advanced stages of maturity, but the Papaya
meleira virus reduces the fruits’ resistance to fruit flies, allowing them to be infested while still
unripe (Martins et al. 2012). When papaya fruits are harvested in early stages of maturity, the
risk of infestation by fruit flies is low due to the toxicity of benzyl-isothiocyanate, which is
present in the latex of the fruit (Tang 1973; Martins et al. 2012). Analysis of the data from
2006 to 2010 provided strong evidence of the positive effect of fruit harvest management at
the start of ripening.
Our results also indicate that fruit fly richness showed no significant variation in response to the
distance of the papaya orchards from the Atlantic forest remnant. However, fruit fly abundance
changed significantly in response to this factor for the full data series and for two subseries –
the continuous implementation of the systems approach and the impact on abundance. It is
possible that the distance analysed was not far enough to capture richness changes but was far
enough to affect fly abundance. Some authors have reported that native vegetation acts as a
reservoir for economically important Anastrepha species (Uramoto et al. 2008; Uchôa and
Nicácio 2010; Garcia et al. 2017). However, few studies have assessed the influence of nearby
vegetation on orchards’ fruit fly communities. A pioneer study in the state of Minas Gerais,
Brazil, showed that guava orchards adjacent to fragments of native vegetation harbour fruit fly
species in common (Querino et al. 2014), with the abundance of Anastrepha tending to be
higher near preserved native forest due to the availability of hosts for the flies.
Comparison of data for two of the subseries – continuous implementation of the systems
approach (1998–2005) and impact on richness (2006–2010) – showed a clear change in
species richness. The richness of flies was perceptibly higher from 1998 to 2005 than from
2006 to 2010; this decrease was probably due to the systems approach implementation.
Generally, crop environments have lower insect diversity than natural areas do because of the
lower diversity of host plants in crop areas (Aluja 1999; Gurr et al. 2003). However,
the richness analysis discussed here emphasises another aspect, this involving host plants and
the diversity of species exploiting the same resource, as some species of Anastrepha are among
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.3214 Araujo et al.
the main pests of fruit grown in Brazil (Araujo et al. 2019). The diversity of tephritids in fruits
depends on the association of each fly species with its host plants. Certainly, the attractiveness of
different plants may influence the presence of species in host plants, thereby impacting the
community structure of insects visiting the fruits (Almeida et al. 2019).
Model selection
The information criterion weights differed among the periods investigated in the present study,
especially when comparing the continuous implementation of the systems approach with the
impact on richness subseries, with the Gompertz model usually best describing the nonlinear
shape between species richness and area in each period. The ecological patterns produced by
the theoretical curves are similar to each other for the full series and the continuous
implementation of the systems approach subseries. These results were expected, because the
continuous implementation of the systems approach period is part of the full series. However,
the impact on richness period clearly shows very different richness values compared to the
continuous implementation of the systems approach period – indicating a strong and lagging
effect of the previously implemented management actions.
The sigmoid pattern observed in the relationship investigated here and described by the
Gompertz model takes place generally in more complex systems, differing from patterns
frequently observed in studies of this nature, including insects (Lomolino 2001). In the
present study, we tested 19 models that provided a wide range of possibilities for describing
the relationship between area and species. We found that the Gompertz function provided the
best shape for the full series and the continuous implementation of the systems approach
subseries. For the impact on richness subseries, no specific model described its trajectory,
probably in response to the low values of the series that resulted from the papaya pest
management programme. According to the information criterion weights, the Weibull3
model best matched the shape for impact on richness data, with the lowest Akaike
information criterion value of all models tested. The three functions – Weibull3, Weibull4,
and Gompertz – resulted in sigmoid curves; we then fitted the three functions to the impact
on richness subseries and found a sigmoidal pattern similar to those previously observed in
the full series and continuous implementation of the systems approach subseries but with
lower theoretical richness values.
The analysis of the relationship between richness values and area size indicates that higher
richness was predominantly observed in areas smaller than 50 ha. This result can be linked to
inter-habitat differences and influences, especially with respect to size and proximity of other
areas such as urban environments or forests, as observed in the presence of different taxonomic
groups (Lomolino 2001; Rico-Gray and Oliveira 2007). The papaya orchards investigated in the
present study exhibited different area sizes and were relatively close to each other, with the
predominance of aggregated areas varying their sizes. These conditions may have influenced
the concentration of values below 50 ha.
Richness partitioning
The diversity partitioning analysis indicated that most of the richness values were within βarea
(an estimate of the mean difference between and the diversity predicted for the largest area) for the
full series and the continuous implementation of the systems approach subseries. Despite the
similarity in terms of fruit fly richness within these series, comparison of all the series showed
a significant decrease in fruit fly richness when comparing the full or continuous
implementations of the systems approach series with the impact on richness subseries. This
decrease was likely influenced by the effects also derived from the systems approach. There is
an apparent similarity between Figures 6A and B due to the full and continuous
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 15
implementation of the systems approach series being the same except for the time periods.
However, the distribution of points differs slightly with respect to the asymptote.
The dotted line close to 10 shows a trend towards change in the point distribution as a
transition between the continuous implementation of the systems approach and the impact on
richness periods. In the impact on richness subseries (Fig. 7), the distribution of points differs
entirely from the continuous implementation of the systems approach (Fig. 6B). The decrease
in numbers from the systems approach affects the width of βarea. The parametric space for
βarea is narrower in the impact on richness subseries than in the continuous implementation
of the systems approach, probably in response to the decrease in number of fly species. The
changes in beta diversity of insects living in Atlantic forest remnants, including fruit flies,
were analysed by Novotny et al. (2007), who emphasised the relationship between diversity
and area and also discussed the role of host plant diversity in the relationship between
richness and area size in tropical areas, with results indicating a low rate of change in beta
diversity for tropical forests.
Although the preference for host plants defines the presence of several species in different
environments, the movement of species – especially polyphagous species – also is highly
important in the configuration of insect communities (Mazzi and Dorn 2012). The movement
of insect pests from one place to another is, on the one hand, associated with their origin,
abundance, and spatio-temporal distribution. But this movement is also capable of influencing
species richness at other localities at different scales because of the relationships to different
environments and to other species, as resource availability significantly influences the balance
of intra- and interspecific relationships (Senger et al. 2009; Mazzi and Dorn 2012).
The richness change in βarea in the context of diversity partitioning may also have important
implications for the expansion of fruit fly populations and species distributions, because some of
the insects affected by pest management techniques may attempt to establish in other locations,
including in new orchards. The maximum richness clearly decreased when the continuous
implementation of the systems approach subseries was compared to the impact on richness
subseries, with values decreasing from 6.55 to 1.83. This result highlights the importance of
using richness-partitioning analysis for problems in agriculture, mainly when species richness
is involved. The richness of fruit fly species depends directly on their host plant preference,
but this association is also influenced by the landscape, particularly when orchards are
scattered in different areas, including near Atlantic forest remnants (Monteiro et al. 2019).
Diversity partitioning has received intensive attention from ecologists who are interested in
the functionality of the diversity decomposition, describing different levels of coverage for the
number of species distributed at different spatial scales (Jost et al. 2010; Veech and Crist 2010;
Chao et al. 2012). This approach has been discussed intensely in terms of concepts and
theoretical aspects important for species diversity, particularly the use of additive or
multiplicative partitioning to quantify and interpret alpha, beta, and gamma diversity, at
multiple scales of sampling.
The study proposed by Chao et al. (2012) shows that both additive and multiplicative beta
diversity can be useful in quantifying and analysing community composition, but neither are
appropriate when comparing multi-site compositional similarity because of the dependence on
number of communities and alpha diversity, respectively, for multiplicative and additive beta
partitioning. However, the objective of the present study does not require the condition of
removing the dependence of the above-mentioned variables to compare species diversity
among areas. The data obtained were analysed to account for the influence of the area size on
the fruit fly species richness distributed in papaya orchards. We chose a way to investigate the
relationship between area and fruit fly richness at different beta-diversity scales, which are com-
patible with the main aspects of the diversity partitioning approach (Henry and Stevens 2009;
Jost et al. 2010; Veech and Crist 2010; Chao et al. 2012).
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.3216 Araujo et al.
The approach used in this study takes into account the type of data available to analyse fruit fly
diversity in commercial papaya orchards located within Brazilian Atlantic forest areas. The data
exhibit the condition required to use Henry and Stevens’s (2009) formalism, demanding that
gamma describes the total number of species and that alpha describes average richness of
patches, or orchards, in the present study. For the beta-diversity analyses, the partitioning was
necessary to consider the spatial structure of the orchards vis-à-vis nearby native forest,
making it possible to consider the presence of species not always instantly and locally detected.
The additive partitioning employed here makes possible a direct comparison of average
richness among samples, because the measures of diversity were expressed in the same units,
as required by Henry and Stevens’s (2009) approach. Therefore, the analysis used to investigate
fruit fly species richness patterns provided a way to better quantify the relationship between area
or distance and fruit fly species richness.
Losses and gains of species are events generally common in insect communities and depend
on many factors, with time being one of the main factors. In a spatial dimension, species
composition – understood as beta diversity – can vary significantly in response to time
among sites (Legendre and Gauthier 2014; Shimadzu et al. 2015), and statistical methods
have been proposed to evaluate changes in diversity through time at different patches
(Legendre 2019).
Although the approach employed in the present study does not specifically consider losses and
gains of species, the β-replace diversity value considered took into account a highly dynamic
scenario in terms of insect movement, with papaya orchards located close to each other, thereby
allowing species to migrate easily among orchards. This condition provides, at least in the
algorithm used in this study, the multi-site homogeneous replacement chance, which makes
insect losses and gains liable for recognition over time in the entire area, because replacements
are equally possible for all orchards.
Time significantly influenced the dynamics of losses and gains of species in the orchards
investigated, especially when different time-period subseries, comprising data without management
effects and data showing results after systems approach effects, were compared. These conditions
showed the temporal impact of the pest management measures on fruit fly species richness similar to
some extent to what Legendre (2019) discussed. Species richness is highly sensitive to multiple
factors, especially those that influence population demographics and species diversity, including
climate change or human activities (Legendre 2019).
Time series and peaks
The time series investigated in the present study showed several peaks occurring at different
time periods and with different population sizes, especially when the full series or continuous
implementation of the systems approach subseries were compared to the impact on abundance
subseries. Peaks may occasionally be associated with cycles in insect populations, with different
explanations for their occurrence. Immigrating populations may show substantially increased
growth rates in habitats that are closely connected to each other, reducing the time period
between peaks in situations where high connectivity exists between habitats. This condition
could result in more rapid changes from peaks to valleys than might occur in isolated
populations, depending on population density (Johnson et al. 2006). Cycles have been
analysed much more often under laboratory conditions, with different variables such as
temperature, humidity, or photoperiod controlled (Prout and McChesney 1985).
The existence of cycles or observation of recurrent peaks in natural conditions is less
common but, in insects, is of particular importance for agricultural landscapes, because of the
risk of outbreaks in crops. In these cases and also in cases of strong interactions, such as in
host–parasitoid systems, spectral analyses provide important information. In this context, the
occurrence or likelihood peaks can be taken into account in decisions regarding interventions
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32The Canadian Entomologist 17
in agricultural systems (Ferreira and Godoy 2014). In the present study, spectral analysis yielded
useful results, indicating that the time series for the full and continuous implementation of the
systems approach would show peaks with a 6.25-year periodicity and the impact on abundance
subseries would show peaks with a 125-year periodicity.
The present study combined three analyses in an attempt to better understand the ecological
patterns inherent in a long-term series. It also projected trends of potential infestations of fruit
flies, focusing on monitoring and papaya-harvesting management. The analyses essentially
complemented each other because they were capable of evaluating the relationships among
time periods, distance from an Atlantic forest remnant, harvesting, and pest management
based on the richness and abundance of Anastrepha fruit flies, which are economically
important for Brazilian fruit growers.
Conclusions
The systems approach implemented in papaya orchards resulted in a significant reduction in
both richness and abundance of Anastrepha species. The Gompertz model satisfactorily described
the sigmoidal relationship between area and species diversity. Additionally, its nonlinear nature
was useful for analysing the parametric space and estimating theoretical richness in the diversity
partition. The periodograms generated in the present study were useful for projecting the
occurrence of future population peaks of fruit flies with critical values for the production of
papaya in commercial orchards, indicating that the systems approach delays the time between
large fruit fly population peaks.
Acknowledgements. The authors thank CNPq, FAPESP (2014/16609-7), and FAPES for
scholarships and financial support. They also thank Janet W. Reid (JWR Associates,
Trumansburg, United States of America) for editing the English text. Two referees and the
editor made useful comments, which undoubtedly contributed to substantially improve
the clarity of the manuscript.
References
Aguiar-Menezes, E.L., Souza, S.A.S., Lima-Filho, M., Barros, H.C., Ferrara, F.A.A., Eurípedes, E.,
and Menezes, B. 2008. Análise faunística de moscas-das-frutas (Diptera: Tephritidae) nas
regiões norte e noroeste do Estado do Rio de Janeiro. Neotropical Entomology, 37: 8–14.
https://doi.org/10.1590/S1519-566X2008000100002.
Almeida, L.M.B., Coelho, J.B., Uchoa, M.A., and Gisloti, L.J. 2019. Diversity of fruit flies (Diptera:
Tephritoidea) and their host plants in a conservation unit from midwestern Brazil. Florida
Entomologist, 102: 562–570. https://doi.org/10.1653/024.102.0333.
Aluja, M. 1999. Fruit fly (Diptera: Tephritidae) research in Latin America: myths, realities and
dreams. Anais da Sociedade Entomológica do Brasil, 28: 565–579. https://doi.org/http://dx.
doi.org/10.1590/S0301-80591999000400001.
Aluja, M., Celedonio-Hurtado, H., Liedo, P., Cabrera, M., Castillo, F., Guillén, J., and Rios, E. 1996.
Seasonal population fluctuations and ecological implications for management of Anastrepha
fruit flies (Diptera: Tephritidae) in commercial mango orchards in southern Mexico. Journal
of Economic Entomology, 89: 654–667. https://doi.org/10.1093/jee/89.3.654.
Araujo, M.R., Uramoto, K., Ferreira, E.N.L., Mesquita Filho, W., Walder, J.M.M., Savaris, M., and
Zucchi, R.A. 2019. Fruit fly (Diptera: Tephritidae) diversity and host relationships in diverse
environments estimated with two sampling methods. Environmental Entomology,
48: 227–233. https://doi.org/10.1093/ee/nvy177.
Chao, A., Chiu, C.H., and Hsieh, T.C. 2012. Proposing a resolution to debates on diversity
partitioning. Ecology, 93: 2037–2051.
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.3218 Araujo et al.
Crawley, M.J. 2013. The R book. Second edition. Wiley Online Library, West Sussex, United
Kingdom.
Dinis, S., Prado, P.I., and Lewinsohn, T.M. 2010. Species richness in natural and disturbed
habitats: Asteraceae and flower-head insects (Tephritidae: Diptera). Neotropical Entomology,
39: 163–171.
Ferreira, C.P. and Godoy, W.A.C. 2014. Ecological modelling applied to entomology. Springer
Nature, Cham, Switzerland. https://doi.org/10.1007/978-3-319-06877-0_1.
Garcia, A.G., Araujo, M.R., Uramoto, K., Walder, J.M.M., and Zucchi, R.A. 2017. Geostatistics and
geographic information system to analyze the spatial distribution of the diversity of Anastrepha
species (Diptera: Tephritidae): the effect of forest fragments in an urban area. Environmental
Entomolomology, 46: 1189–1194. https://doi.org/10.1093/ee/nvx145.
Garcia, F.R.M., Campos, J.V., and Corseuil, E. 2003. Análise faunística de espécies de
moscas-das-frutas (Diptera: Tephritidae) na região oeste de Santa Catarina. Neotropical
Entomology, 32: 421–426. https://doi.org/10.1590/S1519-566X2003000300006.
Gnanvossou, D., Hanna, R., Goergen, G., Salifu, D., Tanga, C.M., Mohamed, S.A., and Ekesi, S.
2017. Diversity and seasonal abundance of tephritid fruit flies in three agro-ecosystems in
Benin, West Africa. Journal of Applied Entomology, 141: 798–809. https://doi.org/10.1111/
jen.12429.
Gurr, G.M., Wratten, S.D., and Michael Luna, J. 2003. Multi-function agricultural biodiversity:
pest management and other benefits. Basic Applied Ecology, 4: 107–116. https://doi.org/10.
1078/1439-1791-00122.
Henry, M. and Stevens, H. 2009. A primer of ecology with R. Springer, New York, New York,
United States of America.
Johnson, D.M., Bjørnstad, O.N., and Liebhold, A.M. 2006. Landscape mosaic induces travelling
waves of insect outbreaks. Oecologia, 148: 51–60. https://doi.org/10.1007/s00442-005-0349-0.
Jost, L., DeVries, P., Wall, T., Greeney, H., Chao, A., and Ricotta, C. 2010. Partitioning diversity for
conservation analyses. Diversity and Distribution, 16: 65–76.
Kemp, J.E. and Ellis, A.G. 2017. Significant local-scale plant–insect species richness relationship
independent of abiotic effects in the temperate Cape floristic region biodiversity hotspot. PLOS
One, 12: e0168033. https://doi.org/10.1371/journal.pone.0168033.
Legendre, P. 2019. A temporal beta-diversity index to identify sites that have changed in
exceptional ways in space–time surveys. Ecology and Evolution, 9: 3500–3514. https://doi.
org/10.1002/ece3.4984.
Legendre, P. and Gauthier, O. 2014. Statistical methods for temporal and space–time analysis
of community composition data. Proceedings of the Royal Society B: Biological Sciences,
281: 20132728. https://doi.org/10.1098/rspb.2013.2728.
Lomolino, M. 2001. The species–area relationship: new challenges for an old pattern. Progress in
Physical Geography: Earth and Environment, 25: 1–21.
Lopes, G.N., Souza-Filho, M.F., Gotelli, N.J., Lemos, L.J.U., Godoy, W.A.C., and Zucchi, R.A. 2015.
Temporal overlap and co-occurrence in a guild of sub-tropical tephritid fruit flies. PLOS One,
10: e0132124. https://doi.org/10.1371/journal.pone.0132124.
Lundgren, J.G. and Fausti, S.W. 2015. Trading biodiversity for pest problems. Science Advances,
1: e1500558. https://doi.org/10.1126/sciadv.1500558.
Marsaro Júnior, A.L., Nascimento, D.B., Ronchi-Teles, B., and Adaime, R. 2012. Faunistic analysis
of the species of Anastrepha Schiner (Diptera: Tephritidae) in three municipalities of the state of
Roraima, Brazil. Brazilian Journal of Biology, 72: 813–819.
Martins, D.S., Alves, F.L., and Zucchi, R.A. 1993. Levantamento de moscas-das-frutas (Diptera:
Tephritidae) na cultura do mamoeiro no Norte do Espírito Santo. Anais da Sociedade
Entomologica do Brasil, 22: 373–380.
Martins, D.S. and Fornazier, M.J. 2014. System approach: tecnologia que viabilizou a exportação
do mamão brasileiro para os Estados Unidos. Incaper em Revista, 4: 84–95.
Downloaded from https://www.cambridge.org/core. Universidade de São Paulo, USP, on 19 Jul 2021 at 11:38:48, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.4039/tce.2021.32You can also read

















































