Inferring longevity from advanced rib remodelling in insular dwarf deer
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Biological Journal of the Linnean Society, 2022, 136, 41–58. With 6 figures.
Inferring longevity from advanced rib remodelling in
insular dwarf deer
JUSTYNA J. MISZKIEWICZ1,2,*, and ALEXANDRA A.E. VAN DER GEER3,*
1
School of Archaeology and Anthropology, Australian National University, Canberra 2601, Australian
Capital Territory, Australia
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
2
School of Social Science, University of Queensland, St Lucia 4072, Queensland, Australia
3
Vertebrate Evolution, Development and Ecology, Naturalis Biodiversity Center, 2300 RA Leiden, The
Netherlands
Received 3 January 2022; revised 13 February 2022; accepted for publication 14 February 2022
Dwarfed insular mammals are of significant interest to evolutionary biology because of their morphological and
physiological adaptations triggered by variables inherent to geographical isolation. One life history trait that is
expected to be modified in relation to body size reduction is extended longevity facilitated by ecological parameters
such as predator and competitor release and fluctuating resource availability. However, systematic examination
of longevity in extreme cases of dwarfism is difficult as such cases are largely represented by fossils of extinct
species. We studied seven rib fragments of the extraordinary endemic Pleistocene dwarf deer Candiacervus (size 2)
to test whether its bone matrix shows evidence for extensive bone remodelling typically seen in large and long-living
mammals. We found that all ribs had well-remodelled Haversian bone, implying several generations of resorption
and re-deposition processes. Osteon density was high relative to small bone cross-section size, with examples of
porosity similar to that seen in elderly animals. We report the first combined micro-morphological and micro-
morphometric record of Candiacervus rib histology, agreeing with previous literature discussing the evolutionary
biology of dwarf ruminants. Further experimental validation of osteon density in extant deer will confirm whether
dwarfed Candiacervus indeed lived longer than expected for its body mass.
ADDITIONAL KEYWORDS: bone histology – Candiacervus – Cervidae – Crete – Haversian tissue – island
biogeography – life history – palaeobiology – ribs.
INTRODUCTION favourable environments where ecologically relevant
predation and competition are minimized or absent,
Mammals typically express a suite of morphological
and food resources can be monopolized (e.g. Drake
and physiological adaptations in response to insular
et al., 2002; Köhler, 2010). In response, mammals may
environments (MacArthur & Wilson, 2001; Lomolino,
evolve smaller or larger body masses to ensure energy
2005; Whittaker & Fernández-Palacios, 2006;
investment allocation suits their insular life histories
Lomolino et al., 2017). Cases of insular dwarfism and
(McNab, 1994, 2002; McNab & Bonaccorso, 2001;
gigantism are well documented and provide evidence
Raia et al., 2003; Raia & Meiri, 2006; Köhler & Moyà-
for evolutionary mechanisms, such as relaxed
Solà, 2009).
selection, ecological release, adaptive radiation
Extraordinary examples of an insular dwarf are four
and anagenetic speciation, acting on island fauna
species of the now extinct deer genus Candiacervus that
(e.g. Berry, 1996; Dayan & Simberloff, 1998; de Vos,
evolved during the Pleistocene on Crete, the largest
2000; Losos & Ricklefs, 2009; Lomolino et al., 2012,
Greek island (8450 km²) (de Vos, 1979, 1984, 2000;
2013; Álvarez-Castañeda & Nájera-Cortazar, 2020;
Kuss, 1975). The smallest species (C. ropalophorus) is
van der Geer, 2020). Insular conditions can produce
estimated to have reached only about 40 cm shoulder
height (de Vos, 1984), with an estimated body mass
*Corresponding authors. E-mail: j.miszkiewicz@uq.edu.au; of 22 kg, while the largest, giant species (C. major)
alexandra.vandergeer@naturalis.nl weighed about 316 kg (van der Geer et al., 2014). Since
© 2022 The Linnean Society of London. 41
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial
License (https://creativecommons.org/licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and
reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact
journals.permissions@oup.com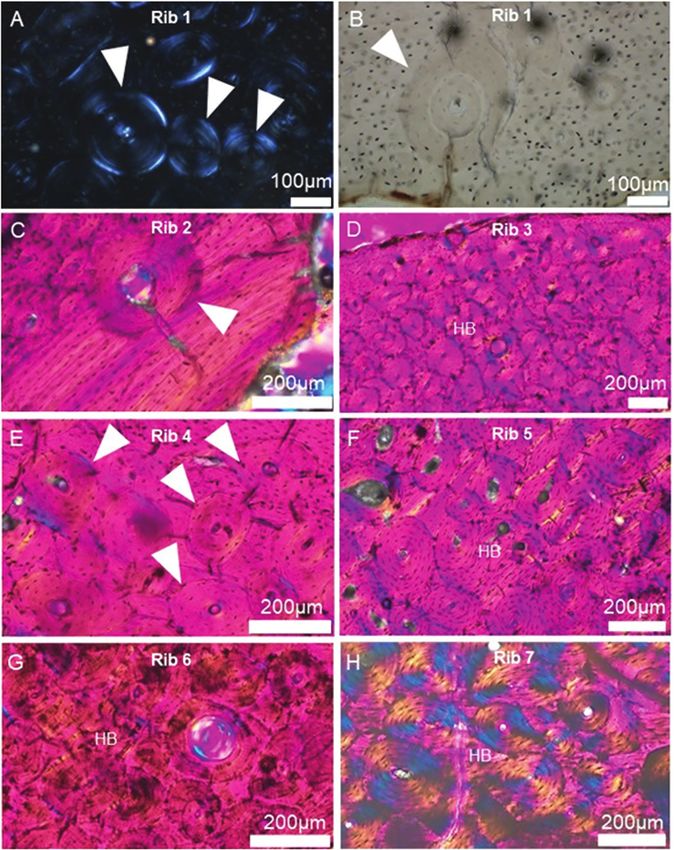
42 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
the discovery of Candiacervus fossils in the late 19th extinct island mammals (deer, proboscideans, mice,
and early 20th century (Bate, 1905; Simonelli, 1907), otters and hippopotamuses) evolving dwarfism or
they have been studied to investigate various topics of gigantism away from their mainland counterparts
relevance to artiodactyl and ungulate evolution (van (Lyras et al., 2021). Kuss (1975) proposed the genus
der Geer et al., 2021). Studies have, for example, focused Candiacervus for the Cretan deer, which included
on questions relating to brain external morphology a larger (C. rethymnensis) and a smaller phenotype
and size reduction relative to body mass shifts under species (C. cretensis). In the years thereafter, the
predator release (Angelelli, 1980; Palombo et al., 2008; following species were described: C. ropalophorus,
Lyras, 2018); antler evolution (Kuss, 1975; de Vos, C. major, C. dorothensis, C. devosi, C. listeri and
1984; van der Geer, 2018); incidence of bone pathology C. reumeri (de Vos, 1984; Capasso Barbato & Petronio,
(Lyras et al., 2016, 2019; Palombo & Zedda, 2016, 1986; Capasso Barbato, 1992; van der Geer, 2018). The
2022); survival and mortality patterns (van der Geer taxonomy of the large biodiversity of Cretan deer has
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
et al., 2014); or ontogenetic growth rates in relation to seen other species proposals, making it a controversial
body size (van der Geer et al., 2006; Kolb et al., 2015a). area in palaeontology (Caloi & Palombo, 1996). This
Using skeletochronology, bone histology, radiography high species diversity makes Candiacervus one of
and dental wear patterns of fossil material, previous the most interesting taxa of Mediterranean endemics
research noted the possibility that the dwarf forms regarding evolutionary radiations (de Vos, 1996, 2000;
of Candiacervus enjoyed long lifespans despite their van der Geer, 2014). Kuss (1975) hypothesized an
dwarfed size (van der Geer et al., 2014; Kolb et al., 2015a; anagenetic trend in Candiacervus experiencing body
Lyras et al., 2016, 2019; Palombo & Zedda, 2016). This size decrease through time. de Vos (1979) suggested
agrees with slow life history trends reported for other an adaptive radiation instead, and recognized six
dwarfed insular ruminants including the Pleistocene size classes and different types of antlers, skulls and
Balearic mouse-goat (Myotragus balearicus) (Köhler teeth. Only the first two size classes (1 and 2) are
& Moyà-Solà, 2009; Jordana & Köhler, 2011; Jordana unequivocally recognized as dwarfed forms (van der
et al., 2012), the black-tailed deer (Odocoileus hemionus Geer et al., 2013). Size 1 is represented by the species
columbianus) (Long et al., 2019) and the anoa (Bubalus C. ropalophorus, and size 2 by the species C. devosi,
depressicornis) (Burton et al., 2005; Clauss et al., 2014). C. listeri and C. reumeri, each with a distinct antler,
The anoa, a dwarf water buffalo, is in fact considered skull and tooth morphology. The three species of size
an example of a ruminant with a particularly high 2 together are often referred to as Candiacervus sp. II
longevity for its body size, ascribed to secondary body in the literature. This informal species nomenclature
size reduction (Müller et al., 2011). However, the range was used in de Vos (1979) and subsequent literature to
of Candiacervus specimens and samples selected as refer to the three size 2 species combined, which were
part of the earlier work (van der Geer et al., 2014; Kolb initially named sp. IIa, IIb and IIc (now respectively
et al., 2015a; Lyras et al., 2016, 2019; Palombo & Zedda, C. listeri, C. devosi and C. reumeri). Diagnostic features
2016) included biomechanically susceptible long bones are limited to the skull, antlers and teeth, while the
(Lanyon et al., 1982), and teeth which are under strong postcranial material cannot be classified other than to
dietary influences (Ungar, 2015). These factors can size class, and hence the need for a group name. No
cloud the ageing-related signal retained by fossils. In complete and articulated skeletons have ever been
this study, we undertook a more controlled approach excavated of any of the species (van der Geer et al.,
to the selection of bones for examination by assessing 2006), meaning that the fossils are represented by
dwarfed Candiacervus ribs. Ribs are thought to best disarticulated and somewhat commingled skeletal
reflect bone metabolic activity without significant elements.
biomechanical influences (Crowder & Rosella, 2007; Our current knowledge about the palaeobiology of
Stewart et al., 2021), and are increasingly being used in dwarf Candiacervus stems from a series of bone and
palaeontological studies of life history in amniotes (e.g. tooth examinations. Lyras et al. (2016) examined the
Waskow & Sander, 2014; Canoville et al., 2016; Waskow histology of bone tissue (femora, antebrachii, metacarpi,
& Mateus, 2017; Waskow, 2019). We hypothesized that metatarsi) and found Haversian remodelling, but no
dwarfed Candiacervus ribs would show evidence for evidence of bone metabolic abnormalities despite
extensive bone remodelling, typically seen in large and observing characteristics of osteoarthritis in nine
long-living mammals, indicating its longevity under individuals. They also remarked that multiple
predator-free island conditions. individuals of the same site (Liko Cave) exhibited
severely worn-down dentition which indicated
the presence of ‘elderly’ deer in the sample (Lyras
Candiacervus spp. fossils and histology et al., 2016: 44). A later examination by Lyras et al.
Pleistocene Crete produced a significant number of (2019) involved radiographic, gross anatomical and
mammalian fossils, which are classic examples of histological analyses of healthy metatarsals from Liko
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58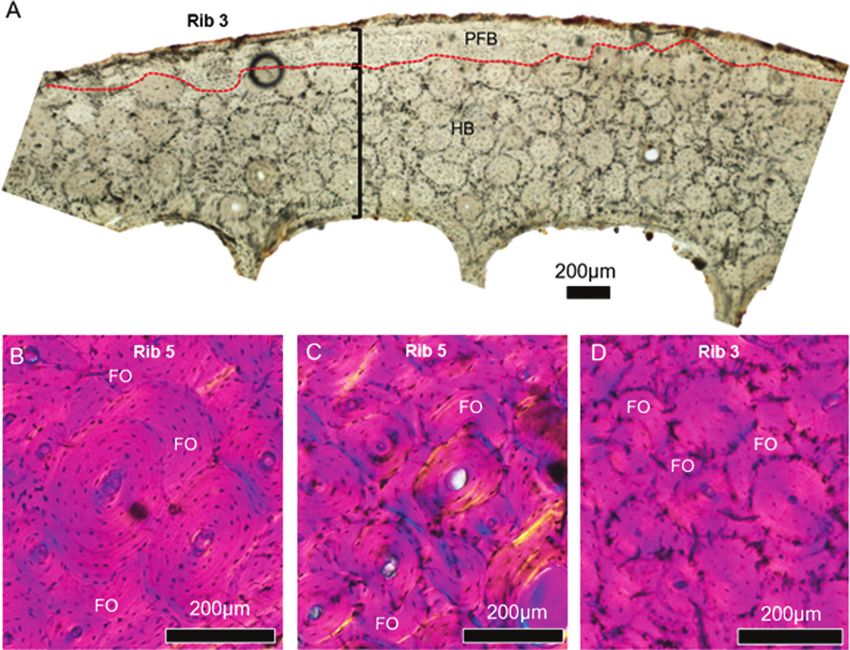
RIB REMODELLING IN INSULAR DWARF DEER 43
Cave, and abnormal metapodials from Mavromouri. best reconstructed from bones that do not experience
Part of the abnormal Mavromouri material had variable and high levels of mechanical load (Canoville
been examined previously to show increased bone et al., 2016; Stewart et al., 2021), which can introduce
porosity with unusual expansion of the marrow dimensional scaling issues (Cullen et al., 2021).
cavity (Sondaar, 1977; Braber, 1981). Dermitzakis Several studies have now shown that ribs tend to
et al. (2006) hypothesized these changes to be due to reflect bone metabolic activity that is not clouded by
metabolic disease. The analyses by Lyras et al. (2019) biomechanics because the only, albeit non-weight-
agreed with this hypothesis proposing secondary bearing and cyclical, mechanical stimulation of ribs
hyperparathyroidism due to either malnutrition arises from muscles attaching to the pleural rib surface
or environmental stress causing the bone lesions. and from regular respiration (e.g. Skedros et al., 2003;
Considerable levels of densely remodelled Haversian Crowder & Rosella, 2007; Canoville et al., 2016; Lad
bone were noted as well. Kolb et al. (2015a) analysed et al., 2019). In reptiles, some mechanical stimulation
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
long bone samples, including femora, and tooth of bone growth may impact cervical ribs involved in
cementum, as part of a larger ontogenetic assessment neck movement (Preuschoft & Klein, 2013), but not
of bone size and body mass growth in several deer taxa. those located near the mid-cage and more ventrally
Their study sectioned bone specimens from newborn, and caudally.
juvenile and adult dwarfed forms of Candiacervus Ribs have also been noted to show strong
(Kolb et al., 2015a) and found that bone remodelling remodelling, valuable in skeletochronological studies
began in the juvenile specimen, and extended lifespans through their retention of annuli, and a different
could be inferred from an allometric perspective. growth and remodelling rate relative to limb bones
Kolb et al. (2015a) reported to have sectioned one (Hillman et al., 1973; Waskow & Mateus, 2017). In
adult Candiacervus rib, but they did not refer to it deer in particular, Hillman et al. (1973) showed that
when describing histology. Kolb confirmed the rib rib tissue from 2-year-old mule deer (Odocoileus
showed extensive bone remodelling which had erased hemionus) experienced cyclical and advanced bone
any evidence of growth marks (C. Kolb, pers. comm. remodelling and active resorption compared to the
December 2021). Based on growth layers in tooth tibia, metacarpus and the metatarsus intra-skeletally,
cementum, they reconstructed the adult form to have which was linked to periods of antler growth. Hillman
lived up to 18 years of age (Kolb et al., 2015a). et al.’s (1973) study is one of the earliest examples
A p o s s i b l e e x t e n d e d l o n g e v i t y o f d wa r f e d of deer rib cross-sectional histology (see their figs 1
Candiacervus from Bate Cave was also inferred using and 2 on p. 327) where a combination of secondarily
a healed oblique fracture in a metatarsal, indicating remodelled and plexiform lamellar bone was reported
the animal had survived injury (Palombo & Zedda, for growing deer. Skedros et al. (2003) also examined
2016). This, however, does not negate high mortality rib histology in O. hemionus in skeletally mature male
at the juvenile stage, which was noted by van der Geer specimens with no evidence for advanced age (i.e. no
et al. (2014) based on mortality and survival patterns. extensive dental wear) to also report a combination of
They found juveniles to have been dying at a rate secondarily remodelled and plexiform bone, along with
similar to that seen in extant mainland ruminants, high remodelling rates and evidence of micro-damage.
but at the same time also long-living for those that The distribution of secondary osteons in these samples
reached the adult phase. Given the relaxed selection was not extensive as the deer had probably not lived
in a predation-released environment, the high juvenile long enough to experience several generations of
mortality was interpreted as a mechanism driving remodelling (Fig. 1).
adaptation to insularity (van der Geer et al., 2014). When working with fossil bone microstructure,
No combined quantitative and qualitative insights ideally a reference collection of extant aged and
into Candiacervus rib microstructural dynamics have sexed deer rib sections would be available to make
yet been reported. Not only is histological analysis of a rigorous interpretation of the histology observed.
vertebrate fossils a powerful tool for reconstructing Kolb et al. (2015a: 7) suggested that extinct and
various aspects of palaeobiology in different taxa extant cervids exhibit a similar arrangement of bone
(Padian & Lamm, 2013; de Buffrénil et al., 2021), it can tissue types in their limb bones. As such, working
be used to compare intra-skeletal bone growth trends with the above-mentioned published histology data
when working with different skeletal elements (Currey, and descriptions, the present study hypothesized that
1979; Prondvai et al., 2018; Stewart et al., 2021). The dwarfed Candiacervus ribs should show extensive
majority of palaeohistological research continues to remodelling given the species’ assumed longevity
sample limb bones (Stein & Sander, 2009; Waskow on Pleistocene Crete. If these deer were still in their
& Mateus, 2017) when investigating biomechanical, earlier stages of ontogeny, we would expected to
skeletochronological and developmental questions. observe parts of primary bone such as those noted for
However, aspects of skeletal physiology are arguably extant taxa.
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58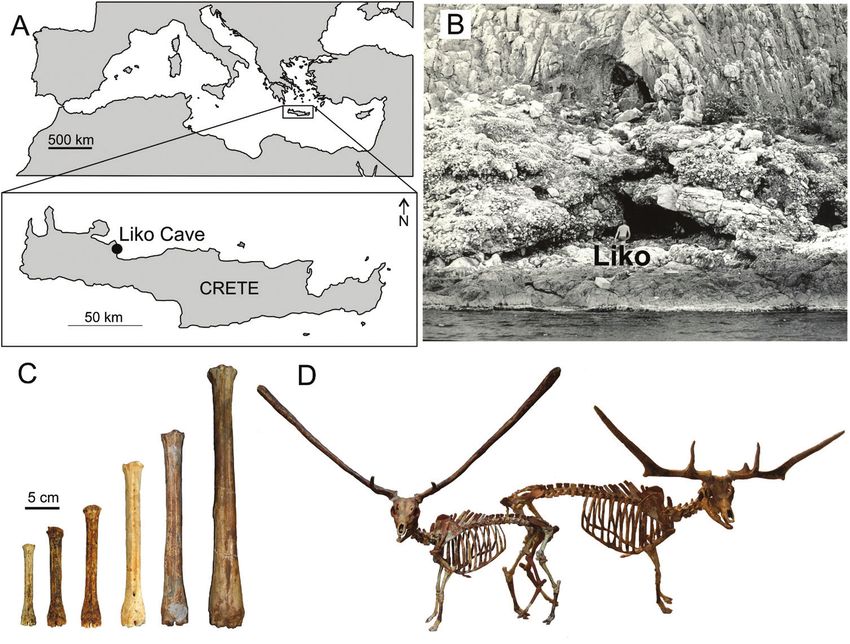
44 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
Figure 1. Rib histology in skeletally mature, male Rocky Mountain mule deer (Odocoileus hemionus), with no advanced
dental wear. A, cross-section histology sketch with x marks indicating secondary osteons and white space denoting primary
bone where no remodelling had yet taken place. B, magnified image of rib histology from the same sample as in A, showing
secondary osteons (SO) with Haversian canals (HC) and micro-damage (white arrows – the micro-crack shown is ~130 µm
long). L: lateral (meaning cutaneous), M: medial (meaning pleural), Cr: cranial (meaning superior), Cd: caudal (meaning
inferior). Both segments A and B are from figures 5 (p. 842) and 4 (p. 841) in Skedros et al. (2003). Permissions to reproduce
were issued by John Wiley and Sons, licence numbers 5211100893380 and 5212751122379.
MATERIALS AND METHODS (inferior, in a quadrupedal stance) base of the rib cage.
The proximal rib segment has a characteristic costal
Seven rib bone fragments from size 2 Candiacervus
facet and a head, both of which precede rib curvature
fossils recovered from Liko Cave were studied (Fig. 2). It
(Ramsay & Lyman, 2014). The sternal end is flatter
cannot be ascertained exactly how many Candiacervus
and less curved. Side can be determined through the
individuals these ribs constitute. We here refer the
positioning of the head, curvature of the shaft and
material to size 2, as it is not possible to assign ribs to
a caudal location of the costal groove (Ramsay &
one of the three size 2 species, which are based on skull,
Lyman, 2014). Our identification includes estimated
antler and dental features, in agreement with the earlier
siding (left/right/unsided), shaft location (vertebral,
palaeohistology studies on this deer. The specimens
mid-shaft, sternal) and number range denoted with a
were selected at random from a larger assemblage
question mark (?) to indicate that it is not a definite
curated at the Museum of Palaeontology and Geology
identification (from one to 13 as deer have 13 pairs
of the University of Athens (Greece). More than 6000
of ribs). The external morphology of each fragment
disarticulated fossils ascribed to Candiacervus come
was also assessed against a composite mount of dwarf
from Liko Cave (Lyras et al., 2019). Several coastal
Candiacervus (van der Geer et al., 2006; Fig. 1D), and
caves in Crete have yielded Candiacervus material, but
by cautiously referring to methodologies of rib position
at Liko Cave (Fig. 2), more than 95% of fossils belong
identification developed for extant deer species (e.g.
to size 2 (Lyras et al., 2019). The remainder belongs
Odocoileus virginianus; Ramsay & Lyman, 2014). We
to sizes 3 (C. cretensis), 4 (C. rethymnensis) and 5
acknowledge that the sectioning location of each rib
(C. dorothensis) (de Vos, 1984, 2000).
(see below), and the uncertainty of estimated rib side
Because the rib fragments used in our study come
and number, might link to bone remodelling stages
from a disarticulated and commingled assemblage, it is
observed across the sample due to the trajectory of
difficult to identify their anatomical position (number)
rib bone formation (e.g. proceeding from proximal to
(Fig. 3). However, certain anatomical landmarks of
distal; Waskow & Sander, 2014), which is a limitation
each rib allow for an estimation of whether the rib
of this study. A further limitation is that we make no
derives from the cranial or caudal aspect of the rib
direct comparison with rib samples from documented
cage (Ramsay & Lyman, 2014). For example, rib shafts
extant deer species, but rely on published literature
become flatter as they curve towards the ventral
and existing descriptions of histology.
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58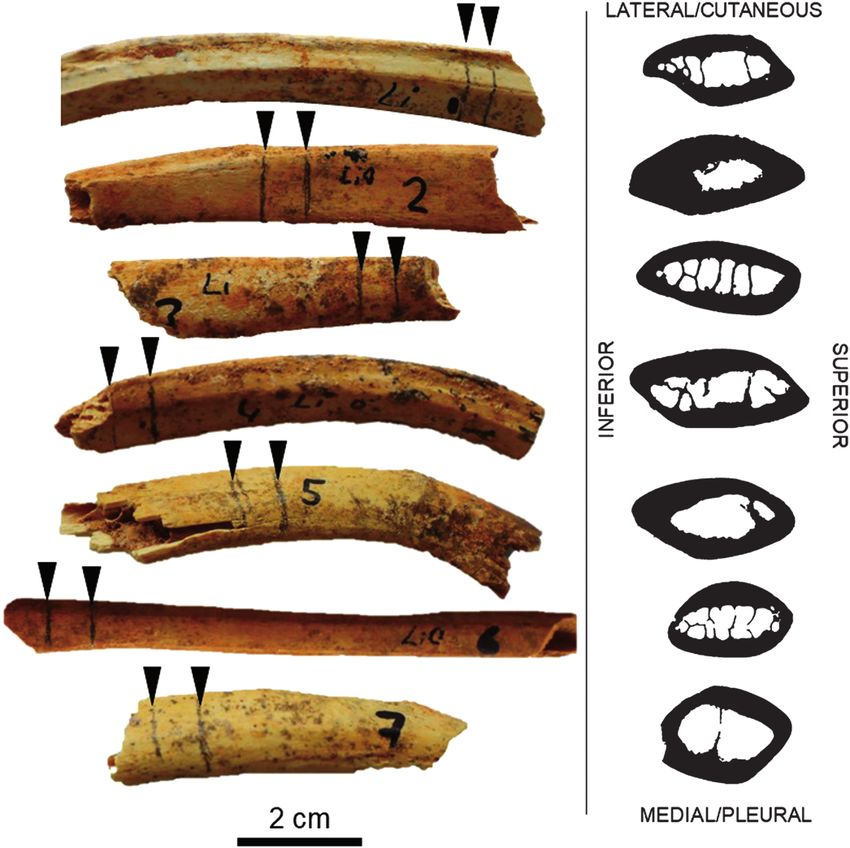
RIB REMODELLING IN INSULAR DWARF DEER 45
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
Figure 2. A, the Mediterranean region with the location of the island of Crete (Greece). B, Liko Cave (photograph John de
Vos) with adult person for scale. C, six size classes of the endemic Pleistocene deer Candiacervus of Crete, here represented
by metatarsals. Material curated at Museum of Palaeontology of the University of Athens, Greece (AMPG; smallest four
sizes) and Museum of Palaeontology of ‘La Sapienza’ University of Rome, Italy (largest two sizes). D, two dwarf species of
Candiacervus, representing size 1 (left, C. ropalophorus, composite mount at AMPG) and size 2 (right, C. devosi, digital
mount). Artwork by George Lyras.
Cross-sectional geometry and histology slides were next ground on a series of grinding pads
Approximately 1-cm-thick sections were removed of various coarseness (400–1200), and polished using
from each fragment using a Dremel tool with a rotary a paste made from Buehler polishing powder. Each
blade. The sections were removed as close as possible slide was bathed in an ultrasonic cleaner, dehydrated
from broken edges of the rib fragments to not damage in a series of ethanol solutions at concentrations of
the remainder of the rib further. The preparation of 90%, 95% and absolute, and cleared using xylene. The
thin sections followed standard methods (Miszkiewicz completed thin sections were covered with glass slips
et al., 2019, 2020; Walker et al., 2021). Each extracted using DPX glue. Imaging was undertaken using an
section was embedded in Buehler epoxy resin. The Olympus BX53 microscope and a DP74 camera using
embedded blocks were cut in half transversely through transmitted, polarized and lambda compensated light.
the shaft using a Kemet MICRACUT 151 low-speed A stitched image of a full cross-section was recorded
precision saw equipped with a diamond blade. This first at a magnification of 10×. Further regions of
exposed the transverse surface of each sample, which interest were captured using 20× and 40× objectives.
was then glued onto a microscope glass slide using The stitched histology image was converted to black
Stuck epoxy glue. Once dry, the glued ‘thick’ section and white using Photoshop. It was next imported into
was further trimmed on the low-speed saw. The Fiji/ImageJ2 (Schindelin et al., 2012) for thresholding,
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–5846 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
Figure 3. The samples of Candiacervus size 2 ribs arranged in order of original labelling (1 to 7). The sectioning location
for histology is marked with a pencil at locations of the rib fragments where minimal further damage to the specimens
would otherwise take place (indicated with black triangular markers). The right panel shows corresponding images of cross-
sections (these are not to scale).
converting bone into black pixels and the background descriptions of the bone tissue matrix followed by
image into white pixels. The image was cropped close histomorphometric data are provided. Haversian tissue
to the borders of each cross-section. Using the Fiji/ was categorized into a severity scale, which consisted
ImageJ2 ‘freehand’ tool and a touch screen stylus, the of ‘low’, ‘moderate’ and ‘high’ remodelling as is referred
inner border of each slice, demarking medullary space, to in previous publications (e.g. Martinez-Maza et al.,
was carefully traced. The selected space was cleared 2014; McHugh, 2014; Eleutério et al., 2015). ‘Low’ bone
using the ‘fill’ tool in Fiji/ImageJ2 so that the medullary remodelling is defined as extremely sporadic with only
cavity was composed of white pixels only, clearing any isolated secondary osteons found throughout the bone
trabecular spicules visible. The image was converted to tissue. Remodelling that is limited to the intra-cortical
16-bit. Cross-sectional geometry was applied to collect portion of bone and where only a single generation of
total area (TA), medullary area (MA), and cortical area remodelling can be detected can be considered ‘moderate’.
(CA), from which CA/TA (%CA = CA/TA × 100) was Remodelling that is widespread and shows several
calculated (Skedros et al., 2003). CA/TA should allow generations of secondary osteons superimposed upon one
us to gauge cortical-to-medullary bone space proportion another indicates highly remodelled Haversian bone.
distribution across all the rib samples (Little, 1984). The above morphological descriptions were further
A Bone-J (Domander et al., 2021) ‘slice geometry’ plugin complemented through bone histomorphometry.
was used to obtain measurements of the total cross- Following methods reported by Lyras et al. (2016,
section, from which CA and MA were extracted. 2019) and Stewart et al. (2021), and bone histology
Histological examination followed terminology standards (Dempster et al., 2013), regions of interest
and definitions by Francillon-Vieillot et al. (1990) from each rib image were examined quantitatively
and Padian & Lamm (2013). Histomorphological using the Fiji/ImageJ open access software (Schindelin
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58RIB REMODELLING IN INSULAR DWARF DEER 47
et al., 2012). Osteocyte lacunae counts/densities classified as having been subject to high remodelling
were not included because of a patchy preservation (Table 1), two samples experiencing moderate–high
of the lacunae. In total, data for four variables are remodelling, and one sample showing evidence of
reported: vascular canal density (Ca.Dn) defined as somewhat moderate remodelling. In this section each
the number of all canals divided by the area of region rib specimen is referred to as numbered in Figure
of interest; vascular canal diameter (Ca.Dm) and 3 (see Table 1 for full IDs). Rib 1 produced the only
osteon diameter (On.Dm), defined as the maximum sample that showed isolated, though neighbouring
diameter of canals and osteons (accounting for any in places, secondary osteons which were widespread
irregularly shaped units); and osteon density (On.Dn), but punctuating (i.e. not erasing) sheets of largely
defined as the number of both secondary (intact and avascular parallel-fibred bone seen from the mid-
partially remodelled osteons) and all primary osteons cortical to sub-periosteal regions of the thin section.
per region of interest (Lyras et al., 2016, 2019; Stewart As such, Haversian remodelling was mostly limited to
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
et al., 2021). The counts were made using the ‘multi- the inner cortical bone space. The secondary osteons
point’ tool, whereas the diameter was measured using were loosely spread out (Fig. 4B) and growth marks
the ‘straight line’ tool of Fiji/ImageJ. The diameter were seen although they became obliterated through
measures of vascular canals were only taken from the presence of secondary osteons, and sporadic
intact vessels (excluding enlarged resorptive cavities). double zonal (indicating an arrested phase of osteon
As such, there were minor discrepancies between the formation) osteons (Fig. 5B). Simple longitudinal
total number of vascular canals used in the calculation vessels and primary osteons were observed closer to
of densities, and the number of vessels from which the endosteal and sub-periosteal regions of cortical
diameter was measured. Two specimens from the entire bone. It is possible this rib represents the youngest
sample were re-examined to double check for technical deer in the entire sample.
errors in image analysis. The bone histomorphometry The bone histology of rib fragments 2 and 7 was
derives from two 2-mm 2 rectangular regions of similar to that reported for Rib 1, whereby the majority
interest located along the midpoint of the pleural and of secondarily remodelled tissue was constrained to the
cutaneous parts of each rib slice. These data will be intra-cortical space, and a band of mostly avascular
used in a descriptive manner to quantify size changes parallel-fibred bone encapsulated the outer cortex
in histology and cross-sectional geometry. (e.g. Fig. 5C). In Ribs 2 and 7, however, the cutaneous
portion of the bone showed much stronger remodelling
than the pleural side. There, several generations of
remodelled secondary osteons were seen across bone
RESULTS
tissue stretching from the endosteal to periosteal
There was some morphological variation across the borders of bone (e.g. Fig. 5H). As such, these two ribs
sample on the basis of exterior rib size (Table 1), were classified as moderately–highly remodelled. Both
which was expected due to the sectioning performed ribs appeared to show an external fundamental system
on different locations of each rib. However, once %CA (EFS; Calderón et al., 2019) made up of largely parallel-
was considered, it became clear that all rib fragments fibred bone marked with growth lines (Fig. 4C, H). Rib
were composed of similar amounts of cortical bone with 7 also shows several lines of arrested growth (LAGs;
two ‘outliers’ of maximum 84% and minimum 56% Fig. 4H), which were not possible to trace around the
(Table 1). The shape of each cross-section also varied section given the remodelling.
somewhat throughout the sample, but it consistently The remaining four rib fragments (Ribs 3–6)
ranged from an oval to more circular outline (Fig. 3). examined here were all categorized as showing high
All cross-sections had ‘empty’ medullary spaces, with levels of secondary remodelling, with their histology
four out of the seven rib fragments showing multiple being made up of widespread true Haversian tissue
trabecular spicules within the medullary cavity (Fig. 3). (Figs 4D–G, 5D–G). Rib 3 was almost entirely strongly
Nothing unusual was noted about the basic morphology remodelled with several generations of secondary
of each rib, with all samples also showing suitable osteons with the exception of a relatively thin (~200 µm
histological preservation for further examination. The in width) band of parallel-fibred bone (PFB), part of
key diagenesis obscuring histology was concentrated on the EFS located on the cutaneous side of the rib (Fig.
the endosteal border and in localized patches obliterating 6A). The transition between Haversian bone and this
osteocyte lacunae in some locations. band of PFB was clear and the border was irregularly
disrupted by secondary osteons (Fig. 6A). In Rib 4, an
unremodelled component was that of endosteal lamellar
Bone histomorphology bone (Fig. 4E). This slide also showed evidence for a
All rib fragments showed remodelled Haversian variety of osteon variants. At least one ‘giant’ osteon,
bone (Figs 4–6). Of the seven samples, four were measuring 660 µm in diameter with a relatively giant
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–5848
Table 1. Measurements for Candiacervus (size 2) rib cross-sectional geometry (TA – total area in mm2, MA – medullary area in mm2, CA – cortical area in mm2)
and histology (BMU – bone multicellular unit products*, Ca.Dn – vascular canal density per mm2, Ca.Dm – vascular canal diameter in μm, On.Dm – osteon
diameter in μm, OPD – osteon population density computed from remodelled and intact osteons per mm2)
Specimen ID Bone Cross-sectional geometry Histology
TA MA CA CA/TA Remodelling BMU products* Ca.Dn Ca.Dm On.Dm OPD
Candiacervus II Liko Cave Right mid-shaft cranial thoracic 46.86 16.57 30.29 65% Moderate Type I, double 26.34 25.14 163.65 21.46
temp n. 1 t/m 7 (R1) rib 10th–12th vertebral end zonal osteons
Candiacervus II Liko Cave Unsided mid-shaft rib sternal 46.05 7.56 38.49 84% Moderate– Type I 29.08 23.61 140.45 28.47
temp n. 1 t/m7 (R2) end 5th–9th high
J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
Candiacervus II Liko Cave Unsided mid-shaft rib 5th–9th 28.35 12.51 15.84 56% High Type I 29.55 25.80 137.42 41.86
temp n. 1 t/m7 (R3) sternal
Candiacervus II Liko Cave Left mid-shaft rib 10th sternal 47.63 16.68 30.95 65% High Type I, Type II, 30.84 28.52 156.03 31.41
temp n. 1 t/m7 (R4) drifting
Candiacervus II Liko Cave Right mid-shaft rib 5th–9th vertebral 55.6 17.13 38.47 69% High Type I, resorp- 21.95 45.96 204.278 23.69
temp n. 1 t/m7 (R5) tion cavities
Candiacervus II Liko Cave Right mid-shaft 13th rib vertebral 29.36 11.25 18.11 62% High Type I, resorp- 27.55 28.12 171.86 29.75
temp n. 1 t/m7 (R6) tion cavities
Candiacervus II Liko Cave Unsided mid-shaft rib 5th–9th sternal 74.09 21.23 52.86 71% Moderate– Type I 20.15 31.89 182.01 24.35
temp n. 1t/m7 (R7) high
*We use ‘products’ to denote microscopic evidence of BMU activity in the form of different types of osteons or their formation stages (e.g. resorption, complete osteon). Type I is the common secondary
osteon; Type II is the embedded, double-zone osteon with two cement lines, and drifting osteon ‘curls’ through the cortex (see Crescimanno & Stout, 2012).
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022RIB REMODELLING IN INSULAR DWARF DEER 49
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
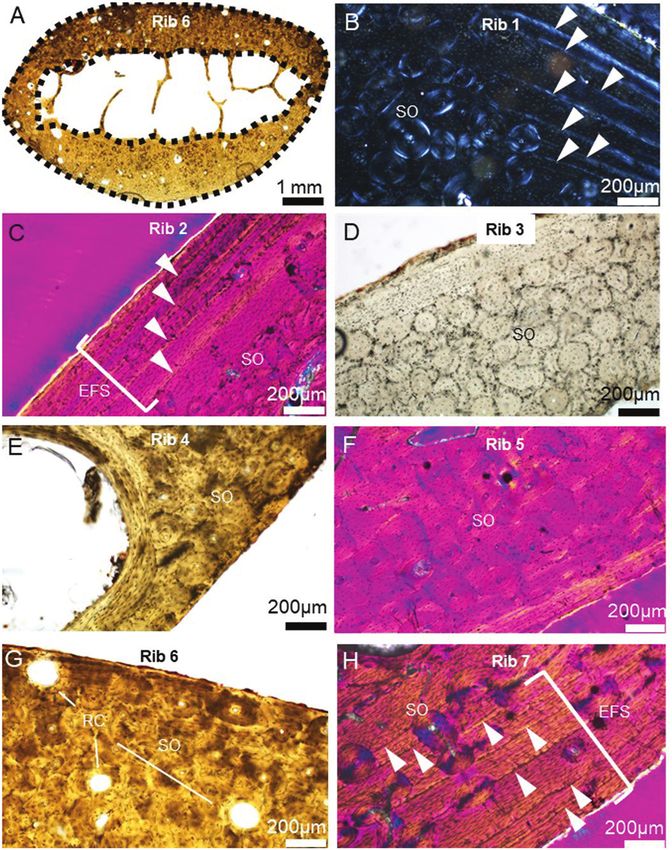
Figure 4. Key bone histology characteristics in dwarf Candiacervus (size 2) observed in this study (see Table 1 for specimen
IDs). A, a cross-section through Rib 6 with black dashed lines marking the outer (periosteal) and inner (endosteal) borders
used in computing total and medullary area (orientation: top is medial, bottom is lateral, left is superior, right is inferior).
White triangular markers in B (cutaneous cortex in superior rib region), C (pleural cortical midpoint) and H (pleural cortex
in superior rib region) point to growth marks and lines of arrested growth. RC in G (pleural cortex in superior rib region):
resorption cavities; SO: secondary osteon(s) in all images including D (cutaneous cortex in inferior rib region), E (pleural
cortex in superior rib region) and F (pleural cortical midpoint). EFS: external fundamental system.
canal of 310 µm diameter (probably an incomplete been experiencing active BMU activity at the time of
remodelling event of a bone multi-cellular unit, BMU; death (Ryser et al., 2009).
Ryser et al., 2009), was noted, along with instances
of type II (one osteon embedded within another) and
drifting (osteons which ‘curl’ horizontally) osteons Bone histomorphometry
(Skedros et al., 2007; Cooke et al., 2021). In addition to The densities of vascular canals did not change
strong remodelling (Figs 4F, G, 5F, G, 6B, C), Ribs 5 and much throughout the sampled regions of interest
6 had several instances of active resorption cavities across the seven ribs, but there were instances
(Fig. 4A, G), with Rib 5 displaying resorptive bays of relatively increased (when compared to the
coalescing into larger pores. These two ribs would have values recorded for the other ribs) osteon densities
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–5850 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
Figure 5. Haversian bone characteristics in dwarf Candiacervus (size 2) observed in this study (see Table 1 for specimen
IDs). White triangular markers in A (cutaneous cortex in inferior rib region), B (pleural cortical midpoint), C (pleural
cortex in superior rib region) and E (pleural cortical midpoint) point to cement lines of fully formed secondary osteons. The
orientations for the other images are: D (cutaneous cortical midpoint), F (pleural cortex in superior rib region), G (pleural
cortex in inferior rib region) and H (pleural cortical midpoint). In B, a double-zone osteon is indicated with the white arrow
marker. HB: Haversian bone.
in Ribs 3 and 4 (Table 1). When considered in DISCUSSION
relation to %CA, these two high osteon densities
Crete, the largest Greek island, has been isolated
were associated with some of the thinner cortical
from the mainland for roughly 5 Myr (Legakis &
slices from across the entire sample of ribs. As
Kypriotakis, 1994). Pleistocene Crete supported rich
such, smaller ribs still produced large amounts
endemic mammal faunas with in total 17 species of
of Haversian bone. Furthermore, the diameter of
endemic mammals, consisting of dwarf elephants
osteons (On.Dm) appeared to also relate positively
and mammoths, dwarf hippos, several species of
to the diameter of vascular canals (Ca.Dm). Across
deer (varying in size from dwarf deer to giant deer),
the whole sample, the values of Ca.Dm increased
giant mice, an otter and the Cretan shrew (Lyras
along with the values of On.Dm (Table 1).
et al., 2021). Only one species, the Cretan shrew,
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58RIB REMODELLING IN INSULAR DWARF DEER 51
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
Figure 6. Images illustrating the large extent of bone remodelling in ribs from dwarf Candiacervus (size 2; see Table 1 for
specimen IDs). A, a stitched image of a portion of cutaneous cortical bone showing an irregularly interrupted border (red
dashed line) between dense Haversian bone in mid- and deep cortex, and parallel-fibred bone (PFB) in the sub-periosteal
region (see black lines subdividing the two tissue types). B–C (pleural cortical midpoint in Rib 5) and D (cutaneous cortical
midpoint in Rib 3) show generational deposition of secondary osteons whereby complete osteons erase the evidence of older,
fragmented osteons (FO).
has survived to the present day (Reumer, 1986). The several years of time to accrue multiple generations
evolution of dwarf and giant island species has made of osteons (Reinwald & Burr, 2008). If Candiacervus
Crete an island of key interest to biogeographers were not relatively long-lived, we would expect only
(van der Geer et al., 2017). The extraordinary case localized remodelling events, no true Haversian
of dwarf Candiacervus with its very small size bone tissue, with more widespread plexiform bone
and disproportionally long antlers in two species present, and well-preserved growth marks, as has
continues to provide fascinating insights into its been shown for comparably sized and skeletally
palaeobiology. The presented rib histology results mature mammals that are relatively short-lived
show that Candiacervus species belonging to size class (Singh et al., 1974; Currey, 2003). For example, while
2 would have lived long enough for its bone tissue experiencing some level of remodelling, skeletally
to experience remodelling to a degree similar to that mature raccoons (Procyon lotor) and red foxes (Vulpes
seen in larger bodied and longer lived mammals, such vulpes) retain growth marks with limited erosion
as pigs, sheep and bears (Currey, 2003; Reinwald & by Haversian remodelling (De Boef, 2009). Equally,
Burr, 2008; Donahue et al., 2021). Each rib fragment advanced age can be deduced for Candiacervus
analysed here showed evidence for secondary bone because in large, skeletally mature rib samples from
remodelling. This can be interpreted as bone having the Rocky Mountain mule deer (Odocoileus hemionus
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–5852 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
hemionus) secondarily remodelled bone did not cover et al. (2015a) for limb bones, and possibly reflected in
the entirety of a cross-section (Skedros et al., 2003). the two rib specimens in our study that still show EFS.
The observations based on ribs agree with previous The observation of EFS in some of the ribs without
literature where extended longevity was reported for extensive remodelling is worth noting as an indicator
dwarf Candiacervus based on histology of long limb of the ribs completing growth. However, associated
bones and tooth cementum (Kolb et al., 2015a) and tooth teeth and other long bone epiphyses would need to be
wear patterns (van der Geer et al., 2014). Rib samples available to determine if the deer individual(s) had
further prove to be fruitful in palaeohistological reached skeletal maturity (Calderón et al., 2019). They
studies of insular dwarfism (see discussion below). reported that the age at which skeletal and sexual
maturity of red deer (Cervus elaphus) is reached varies
with sex. In males, both sexual and skeletal maturity is
Bone remodelling in dwarf Candiacervus and reached at around 4–5 years (Calderón et al., 2019). In
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
limitations females, sexual maturity can be reached at 2.5 years,
All of the rib fragments here were well remodelled, but skeletal maturation is still in progress at that age.
with only one rib showing the lowest extent of Calderón et al. (2019) noted EFS in the femora, but not
remodelling in intra-sample comparisons. This agrees tibiae, of adult Cervus elaphus estimated to a range of
with previously reported presence of remodelled bone ages (3–13.5 years). If the rib fragments showing EFS
tissue in Candiacervus size 2 using multiple bones, in our study were from males, they could be taken to
including the femur, tibia, metatarsus, humerus, indicate adulthood, though sex cannot be reconstructed
radius, ulna, metacarpus and phalanges (Kolb et al., using the fossil material in our study.
2015a; referred to as Candiacervus sp. II). Kolb et al. Taken together, Haversian bone and avascular
(2015a) noted the scattered presence of secondary parallel-fibred bone in the sub-periosteal parts of
osteons in the bones of juvenile Candiacervus, which rib cross-sections supports metabolic bone activity
indicated that bone remodelling processes began maintaining bone tissue well into adulthood of
early in ontogeny. Adult Candiacervus specimens in these dwarfed specimens. This further matches the
Kolb et al. (2015a) also showed remnants of plexiform conclusions regarding ageing based on histology
fibrolamellar bone transitioning into Haversian bone of limb bones by Lyras et al. (2016). However,
in the deep cortex. Lyras et al. (2016) similarly observed discussing longevity in fossil mammals when using
small amounts of plexiform bone in adult femora, histological techniques would ideally also require a
antebrachii, metacarpi and metatarsi of Candiacervus skeletochronological analysis of LAGs (Woodward
size 2, noting substantial remodelling and presence et al., 2013). In the present sample, the presence of
of secondary osteons. Plexiform fibrolamellar bone growth marks and LAGs could be detected in only three
(fast forming) was not seen in any of the present rib specimens, but their analysis was not possible due to
samples, but parallel-fibred bone (slower forming) and the extensive coverage of Haversian bone. Perhaps one
Haversian bone was seen, indicating there was strong limitation to working with ribs, as much of this study
bone remodelling of ribs (Francillon-Vieillot et al., is vouching for (see ‘Utility of ribs in palaeohistological
1990). In the Candiacervus limb bones studied by studies of insular dwarfism’ below), does link to
Kolb et al. (2015a), the scattered secondary osteons (in their advanced remodelling which, ultimately, erases
the juvenile) and more advanced remodelling (in the the evidence of other histological characteristics
adult forms) were all primarily restricted the middle (Woodward et al., 2013). This was previously clearly
cortex in the posterior region. Growth in this region demonstrated by Horner et al. (2000) who noted
was prominent around the linea aspera landmark, hadrosaurid rib sections from Maiasaura peeblesorum
obscuring evidence of growth marks. An ulna section to show strong and extensive remodelling, complicating
from an adult Candiacervus specimen had the an intraskeletal LAG analysis. Having said this,
medullary cavity obliterated by endosteal lamellar Horner et al. (2000) reported the M. peeblesorum ribs
bone subsequently replaced by dense Haversian bone. had accrued three to four superimposed generations
The remaining regions of the ulnar section, however, of secondary osteons. The number of secondary osteon
were still accompanied by patches of plexiform bone. generations has been also proposed as a standardized
The location of remodelled bone in these instances proxy for ontogenetic stages in sauropods (Klein &
indicates behavioural stimulation to the limb bones, Sander, 2008; Stein et al., 2010; Mitchell et al., 2017).
which would not be expected to the same extent in the With further research corroborating our findings using
rib. The secondary osteons in the majority of the rib ribs from extant deer species, a similar proxy could
samples examined in our study spread from the inner be developed for Candiacervus in the future. Some of
cortex to the periosteal border. This implies that some the most remodelled ribs in the present study showed
time had passed since the initiation of more intra- similar levels of osteonal generations (Fig. 5D–H; Table
cortical remodelling, similar to that described by Kolb 1), providing a body size perspective that remodelling
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58RIB REMODELLING IN INSULAR DWARF DEER 53
experienced by dwarf Candiacervus was comparable (Cooke et al., 2021), double-zonal osteons have been
to that experienced by very large animals such as the linked to arrested BMU activity (Stout et al., 2019).
extinct hadrosaurid studied by Horner et al. (2000). Their formation in dwarf Candiacervus points to a
Indeed, it would have been useful to determine the dynamic and varied bone remodelling processes also
age of our Candiacervus specimens through LAGs, seen in long-lived mammals (Crescimanno & Stout,
particularly as new in vivo labelling experiments into 2012; Stout et al., 2019). The histomorphometric data
bone growth and body size are increasingly being illustrate that some of the smaller ribs also happen to
reported for ruminants, validating interpretations express the highest densities of osteons, and vascular
made from LAG counts in fossils (Hayashi et al., 2020; canal size increases with increasing osteon size. This
Calderón et al., 2021). Kolb et al. (2015a) were able to implies that remodelling progressed in Candiacervus
observe unremodelled LAGs in the femoral shafts of size 2 regardless of the small bone size and limited
a juvenile and adult Candiacervus size 2 specimens, mechanical load as is the case with ribs. This matches
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
analysis of which was further supplemented by tooth previous studies confirming that secondary osteon
cementum age estimates of 12 and 18 years. dimensions and bone remodelling processes occur in
While skeletochronology could not be performed mammal tissue regardless of phylogenetic influences
here, the highly remodelled rib histology facilitated (Felder et al., 2017).
the collection of histomorphometric data indicative of Collectively, insights from the present study and
BMU activity, including different stages of resorption those from published histology descriptions allow
and formation (Ryser et al., 2009), and examination of us to hypothesize extended longevity in dwarfed
secondary osteon variants (Skedros et al., 2007; Cooke Candiacervus, which will need to be confirmed
et al., 2021). For example, two of the rib fragments experimentally by evaluating rib osteon densities in
showed several instances of active bone resorption aged extant deer. Dwarf Candiacervus reveals the
inferred from isolated resorption cavities, and those great adaptability of this taxon to its new ecology. Crete
coalescing into larger vascular pores. While there are was a terrestrial predator-free island for Candiacervus
several causes (diet, age, sex/lactation/reproduction/ (van der Geer et al., 2014), and the dwarfed species
a n t l e r f o r m a t i o n , d i s e a s e, h o r m o n a l i s s u e s, would have had sufficient time for their skeletal
biomechanics) underlying active bone resorption that maturity to experience several generations of bone
cannot be specifically determined in fossil bone, in ribs remodelling.
it is often associated with osteopenia, which results
from metabolic diseases or natural degenerative
conditions caused by old age (Reinwald & Burr, 2008; Utility of ribs in palaeohistological studies of
Stewart et al., 2021), and is particularly notable insular dwarfism
in long-lived mammals such as humans (Agnew & Palaeohistological data from extinct island species
Stout, 2012). A possible explanation for the resorptive have previously confirmed versatility in mammalian
cavities in the ribs studied here is that degenerative tissue modification to reflect evolving life histories
processes aged the bone tissue. This would agree with (see Kolb et al., 2015b for a review; Sander et al., 2006;
all the above-mentioned longevity data stemming Miszkiewicz et al., 2019, 2020). Ribs are not a typical
from LAG, dental wear and bone histology research. sampling location in fossil vertebrate histology for
However, without validating this on an extant deer several reasons, including their poor preservation and
with documented osteoporosis, our interpretations susceptibility to become commingled easily, and their
need to be treated cautiously. Furthermore, we must irrelevance to questions of biomechanical origin. As such,
also note that deer are known to experience cyclical limb bones and dental material are usually favoured.
seasonal osteoporosis needed for antler growth, However, ribs can offer a suitable substitute in cases
evidence of which has been reported to occur in rib where curatorial permissions for invasive sampling
samples (Hillman, 1973; Baksi & Newbrey, 1989; cannot be obtained on limb bones (Waskow & Mateus,
Meister, 1956), though not exclusively (see Skedros 2017). Ribs occur in multiple ‘repeated’ forms owing
et al., 2003). Should active antler formation be the to relatively high anatomical number in a vertebrate
case in at least one of our specimens, we would expect skeleton. Their cross-section is also smaller than that
more widespread resorption spaces in addition to a of long bones, meaning the entirety of the cortical and
predominance of primary bone consistent with prime medullary space can be viewed on one glass slide when
aged deer (e.g. 2-year-old O. hemionus, Hillman, 1973). working with larger mammals. This makes technical
Furthermore, the existence of Type II, drifting and preparation more efficient than processing larger pieces
double-zonal osteons in the Haversian bone of the of bone. Several (palaeo)histological studies have now
rib fragments was observed. While it is increasingly argued that ribs are a suitable bone for analyses of
recognized that Type II and drifting osteons occur as physiological fluctuations in the skeleton, including
a result of variation in normal bone microstructure stochastic physiology or disease, because of their limited
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–5854 J. J. MISZKIEWICZ and A. A. E. VAN DER GEER
exposure to repeated and strenuous load (Waskow & samples, Phil Piper (Australian National University)
Sander, 2014; Canoville et al., 2016; Waskow & Mateus, for assistance with rib identification and Christian
2017; Waskow, 2019; Stewart et al., 2021). Evolutionary Kolb for comments on Candiacervus rib histology. We
biologists and palaeontologists are often interested in are grateful to Eric S. Long, John A. Allen, Blanca
longevity questions when working with island dwarfs to Moncunill-Solé, Carmen Nacarino-Meneses, Lucas
better understand adaptation to island conditions (Long Legendre and an anonymous reviewer for feedback
et al., 2019). The present study has attempted to show that improved an earlier version of this article. The
that the use of rib histology can aid in answering such samples were prepared and examined using laboratory
questions. Having said this, we emphasize that this study facilities at the Australian National University (ANU)
relies on a small sample of fossil ribs, and on published funded by the ANU College of Arts and Social Sciences,
sources and descriptions of bone histology in extant and the Australian Research Council (DE190100068).
taxa. Experimental histology samples obtained from Both authors equally contributed to the study
Downloaded from https://academic.oup.com/biolinnean/article/136/1/41/6554026 by guest on 17 June 2022
documented (sexed and aged) extant deer are needed to conception, design, editing and writing of the
validate the use of ribs in palaeohistology for the purpose manuscript. J.J.M. conducted lab work and analysed
of longevity estimations. Future experimental research data. Both authors have read and agreed to this version
should consider sampling longer living captive deer, in of the manuscript. The authors have no conflicts of
addition to those obtained from wild kill, and conduct interest to declare.
a rigorous rib-by-rib (controlling for side and number) Animal ethics clearance was not relevant to this study
analysis. because the examined fossil bone is from an extinct
deer dated to the Pleistocene. Permissions for study and
export of fossils were granted by Dr G. A. Lyras (National
and Kapodistrian University of Athens, Greece), and
CONCLUSIONS
conducted under a Materials Transfer Agreement.
When examining fossils, gross anatomical examination Our examination followed guidelines outlined in the
methods are limiting as they do not access the Member Bylaw on Ethics Statement (Article 12. Code of
underlying bone microstructural dynamics building Ethics), and the Guidelines from the Ethics Committee
and maintaining bone of animals when alive. In of the Society of Vertebrate Paleontology.
such instances, histological sectioning is a powerful
tool for testing hypotheses regarding, for example,
bone metabolic activity in dwarfed extinct taxa,
DATA AVAILABILITY
particularly as it can determine whether fast or slow
forming bone, and experiences of cyclical bone renewal Qualitative descriptions and images of bone
characterized a given sample. When asking questions microstructure are reported and shown in the paper.
about longevity in species represented by unique The thin sections and residual fossil materials will
fossils of significant value, a minimally destructive be repatriated to the University of Athens upon
approach is preferred. This study has shown that completion of this project. All other data have been
bone histology of ribs of a dwarfed Candiacervus deer deposited with open access at Figshare: https://doi.
from Pleistocene Crete showed advanced remodelling org/10.6084/m9.figshare.16608853.
patterns, typically seen in large-bodied and long-lived
mammals. This finding furthers our understanding
of how mammals adapt to constrained island
environments. Opting for ribs allowed us to control for REFERENCES
biomechanical signals stimulating bone remodelling Agnew AM, Stout SD. 2012. Brief communication: reevaluating
in other commonly sampled bones, such as long bones osteoporosis in human ribs: the role of intracortical porosity.
of the limbs. Moreover, the rib histology examination American Journal of Physical Anthropology 148: 462–466.
in this study matched conclusions drawn in previous Álvarez-Castañeda ST, Nájera-Cortazar LA. 2020. Do island
research where limb bones were used. This further populations differ in size and shape compared to mainland
strengthens the utility of ribs. Future research into counterparts? Journal of Mammalogy 101: 373–385.
the palaeohistology of insular fossils is encouraged to Angelelli F. 1980. Endocranial morphology of the dwarf deer
consider the sampling approach presented here. of Crete. Accademia Nazionale dei Lincei 249: 101–109.
Baksi SN, Newbrey JW. 1989. Bone metabolism during antler
growth in female reindeer. Calcified Tissue International 45:
314–317.
ACKNOWLEDGEMENTS
Bate DMA. 1905. Four and a half months in Crete in search
We thank George A. Lyras (National and Kapodistrian of Pleistocene mammalian remains. Geoogical Magazine 2:
University of Athens) for facilitating access to 193–202.
© 2022 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 136, 41–58You can also read

















































