IL-7 Contributes to the Progression of Human T-cell Acute Lymphoblastic Leukemias
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Cancer
Microenvironment and Immunology Research
IL-7 Contributes to the Progression of Human T-cell
Acute Lymphoblastic Leukemias
Ana Silva1,3, Angelo B.A. Laranjeira4, Leila R. Martins1, Bruno A. Cardoso1, Jocelyne Demengeot2,
s Yunes4, Benedict Seddon3, and Joa
J. Andre ~o T. Barata1
Abstract
The importance of microenvironmental factors for driving progression in leukemia has been debated.
Previous evidence has pointed to interleukin-7 (IL-7), a fundamental cytokine to normal T-cell development
and homeostasis, as an important determinant of the viability and proliferation of T-cell acute lymphoblastic
leukemia (T-ALL) cells in vitro. In this study, we report that IL-7 is also a critical determinant of T-ALL
progression. T-ALL cell lines and primary T-ALL samples initiated leukemia more slowly when engrafted to
immunocompromised Rag2/IL2rg/ mice lacking IL-7. This effect was not related to reduced engraftment or
homing of transplanted cells to the bone marrow. Instead, IL-7 deficiency diminished expansion of leukemia
cells in the bone marrow and delayed leukemia-associated death of transplanted mice. Moreover, infiltration of
different organs by T-ALL cells, which characterizes patients with advanced disease, was more heterogeneous
and generally less efficient in IL-7–deficient mice. Leukemia progression was associated with increased Bcl-2
expression and cell viability, reduced p27Kip1 expression, and decreased cell-cycle progression. Clinical
measurements of IL-7 plasma levels and IL-7 receptor (IL-7R) expression in T-ALL patients versus healthy
controls confirmed that IL-7 stimulates human leukemia cells. Our results establish that IL-7 contributes to
the progression of human T-cell leukemia, and they offer preclinical validation of the concept that targeting
IL-7/IL-7R signaling in the tumor microenvironment could elicit therapeutic effects in T-ALL. Cancer Res; 71(14);
1–10. 2011 AACR.
Introduction epithelium and bone marrow stromal cells promote survival
of T-cell acute lymphoblastic leukemia (T-ALL) cells via
Interleukin-7 (IL-7) is essential for normal T-cell devel- production of IL-7 (8, 9). Importantly, IL-7 induces in vitro
opment and homeostasis. Paradoxically, several lines of proliferation of the majority (>70%) of primary T-ALL sam-
evidence indicate that the IL-7/IL-7 receptor (IL-7R) axis ples (10, 11). IL-7 promotes cell-cycle progression and
may also play a significant role in promoting leukemogen- viability of T-ALL cells via activation of the phosphoinosi-
esis. IL-7 can act as an oncogene in vivo, as IL-7 transgenic tide-3-kinase (PI3K)/protein kinase B (PKB)/Akt signaling
mice develop B- and T-cell lymphomas (1, 2), and murine pathway (12), resulting in downregulation of the cdk inhi-
thymocytes spontaneously overexpressing IL-7R have a bitor p27Kip1 and upregulation of Bcl-2 (13). Remarkably,
selective advantage that contributes to proliferation and IL-7 may contribute to resistance to treatment with rapa-
leukemogenesis (3). IL-7 is produced by bone marrow mycin in ALL in general (14) and to imatinib in Bcr/
and thymic stroma (4–7), suggesting that IL-7 is present AblþArf ALL (15, 16). However, these observations are
in the microenvironments where leukemia cells develop, counterweighted by the understanding that most malignant
thus having the potential to directly modulate leukemia cells proliferate independently of external cues. Further-
growth. In vitro studies have shown that thymic more, although IL-7 may have a significant impact on some
primary T-ALL cells cultured in vitro under conditions of
growth factor paucity, it is possible that IL-7 has only a
Authors' Affiliations: 1Instituto de Medicina Molecular, Faculdade de
^ncia, Oeiras,
redundant effect on leukemia expansion in vivo, given the
Medicina da Univer, Lisbon; 2Instituto Gulbenkian de Cie
Portugal; 3Division of Immune Cell Biology, MRC National Institute for diversity of other survival and proliferative signals present
4
Medical Research, London, United Kingdom; and Laborato rio de Biologia in the tumor milieu.
Molecular, Centro Infantil Boldrini, Campinas, SP, Brazil
In the present study, we sought to clarify the importance of
Note: Supplementary data for this article are available at Cancer Research IL-7 for the progression of human T-ALL in vivo, using a
Online (http://cancerres.aacrjournals.org/).
combination of xenotransplant mouse models of human
Corresponding Author: Joa~o T. Barata, Cancer Biology Unit, Instituto de
Medicina Molecular, Lisbon University Medical School, Av. Prof. Egas
leukemia and analysis of primary T-ALL specimens collected
Moniz, 1649-028 Lisbon, Portugal. Phone: 351217999524; Fax: directly from patients. Our results indicate that IL-7 is con-
351217999524; E-mail: joao_barata@fm.ul.pt sumed by and stimulates human T-ALL cells in vivo. IL-7
doi: 10.1158/0008-5472.CAN-10-3606 induces p27Kip1 downregulation and Bcl-2 upregulation in
2011 American Association for Cancer Research. T-ALL cells, promotes their viability and proliferation, and
www.aacrjournals.org OF1
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Silva et al.
contributes to leukemia progression. Our data support the indicated time points. Organs were collected for postmortem
notion that microenvironmental cues, produced in a nonau- analysis. Mice were classified as leukemic when T-ALL cells
tocrine fashion, can partake in leukemia development. represented more than 20% of the bone marrow cellularity,
according to the clinical definition of human leukemia (23). We
Material and Methods further classified mice as preleukemic (0.1% malig-
nant cells in the bone marrow) and nonleukemic (>0.1%).
Cells
Primary T-ALL cells were collected from peripheral blood or In vivo bioluminescence imaging
bone marrow of patients at diagnosis as described previously Mice were intraperitoneally (i.p.) injected with 150 mg
(17). Normal thymocytes were isolated from thymic tissue luciferin/g, anaesthetized, and scanned with an IVIS Lumina
obtained from children undergoing cardiac surgery. Informed bioimaging device (Caliper Life Sciences). Mice were imaged
consent and Institutional Review Board approval were obtained after 7 minutes and total flux (photons per second) was
for all primary leukemia and thymic sample collections. The calculated using Living Image software (Caliper Life Sciences).
IL-7–dependent T-ALL cell line TAIL7 was established by our
group, and it is regularly tested by flow cytometry and PCR (18). Blood and organ analysis
TAIL7 cells share significant similarities with primary leukemic For flow cytometric analysis, collected blood was depleted
samples (18). The growth factor–independent T-ALL cell line of erythrocytes by using red blood cell lysis buffer (BD
HPB-ALL expresses the IL-7R and responds to IL-7. P12-ICHI- Pharmingen). Bone marrow was extracted by flushing off
KAWA is a growth factor–independent IL-7R–negative T-ALL bones. Spleen, liver, and kidneys were mechanically disinte-
cell line. Both cell lines were from a cell bank and characterized grated into single-cell suspensions, which were then washed in
as specified (19). HPB-ALL cells were stably transduced with PBS, stained for 45 minutes at 4 C with monoclonal antibody
lentiviral vectors, kindly provided by Dr. Luigi Naldini (San for human CD2 or CD5 (Becton Dickson), and analyzed using a
Raffaele Scientific Institute, Milan, Italy; 20), driving the expres- FACSCalibur flow cytometer (Becton Dickinson) and FlowJo
sion of luciferase and GFP (HPB-ALL.Luc.GFP). Primary T-ALL software (Tree Star). For histology analysis, small represen-
samples and cell lines were immunophenotyped using stan- tative portions of the collected organs (spleen, liver, and
dard techniques (21) and their maturation stage was classified kidneys) and at least 1 complete femur were preserved in
according to the European Group for the Immunological Clas- formalin at 4 C for 24 to 48 hours with hematoxylin/eosin
sification of Leukemias (EGIL) criteria (ref. 22; Table 1). staining.
Xenotransplantation CFSE labeling for analysis of engraftment efficiency
Protocols and studies involving animals were conducted TAIL7 cells (10 106 cells/mL) were incubated for 10
in accordance with the UK Home Office regulation and local minutes at 37 C with 2 mmol/L of carboxyfluorescein succi-
guidelines. Rag2/, Il2rg/, and Il7/ strains were obtained nimidyl ester (CFSE; Sigma-Aldrich). Cells were washed 3
from The Jackson Laboratory and intercrossed to generate times in culture medium, resuspended as 107 cells/250 mL
compound knockouts required for this study. Il7/Il2rg/ in Iscove's modified Dulbecco's medium/bovine serum
Rag2/ and Il2rg/Rag2/ strains were both on C57Bl6/j albumin, and injected i.v. into mice.
background. Strains were bred in specific pathogen-free facil-
ities at the National Institute for Medical Research (London, Intracellular staining
UK), Instituto Gulbenkian de Ci^encia (Oeiras, Portugal), and Bone marrow cells were stained with anti-human CD2-PE- or
Instituto de Medicina Molecular (Lisbon, Portugal). HPB-ALL. CD5-fluorescein isothiocyanate (FITC)-conjugated antibodies,
Luc.GFP, TAIL7, P12, and primary T-ALL cells (5 106 to 2 fixed in 1 Fix/Perm solution (eBioscience) for 30 minutes at
107 cells) were injected i.v. in a final volume of 250 mL per 4 C, washed in PBS, and resuspended in 1 Permeabilization
injection. Mice were sacrificed when moribund or at the Buffer (eBioscience) for 10 minutes at 4 C. Samples were
Table 1. Immunophenotype and maturation stage of primary T-ALL samples and cell lines
CD1a CD2 CD3 cCD3 CD5 CD7 CD4 CD8 IL-7Ra Maturation stage
Pt#1 þ þ þ þ þ þ þ þ III (cortical)
Pt#2 þ þ þ þ þ þ IV (mature)
TAIL7 þ þ þ þ þ þ þ II (pre-T)
HPB-ALL þ þ þ þ þ þ þ þ þ III (cortical)
P12 þ þ þ þ þ þ III (cortical)
NOTE: Maturation stages were defined according to the EGIL criteria (see Materials and Methods). Plus sign indicates >30% positive
cells; minus sign indicatesPublished OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
IL-7 Accelerates Human T-ALL In Vivo
stained for 30 minutes at 4 C with anti-Bcl-2-FITC antibody in vivo studies using primary human leukemia cells may help
(Dako), anti-Ki-67-PE antibody (Becton-Dickinson), or with elucidate processes unique to human disease. Both recombi-
irrelevant isotype-matched antibody. For viability analysis, nant IL-7 and endogenously produced murine IL-7 are bioac-
bone marrow cells were surface stained for human CD5-FITC, tive upon human lymphocytes, including T-ALL cells (24, 25).
washed and resuspended in 1 binding buffer, and stained with Therefore, we sought to test the in vivo effects of microenvir-
Annexin V–phycoerythrin (PE). Samples were washed and onmental IL-7 on human T-ALL development by xenogeneic
analyzed by flow cytometry. Results were expressed as the transplantation of leukemia cells into Rag2/Il2rg/ hosts
percentage of positive cells in comparison with the negative differing exclusively in the capacity to produce IL-7. In the
control and as specific mean intensity of fluorescence (MIF). absence of Rag2 recombinase and the IL-2 common g-chain,
these hosts lack T, B, and natural killer (NK) cells and there-
Immunoblotting fore tolerate T-ALL transplantation. HPB-ALL T-ALL cells
Cell lysates were resolved by 10% SDS-PAGE, transferred stably expressing luciferase and eGFP (HPB-ALL.Luc.GFP)
onto nitrocellulose membranes, and immunoblotted with were transplanted i.v. into Rag2/Il2rg/ [IL-7 wild-type
antibodies against ZAP-70 (Upstate) or p27Kip1 (Santa Cruz (WT)] and Rag2/Il2rg/Il7/ [IL-7 knockout (KO)] mice.
Biotechnology). Immunodetection was carried out by incuba- Five weeks (35 days) posttransplantation, mice were injected
tion with horseradish peroxidase–conjugated anti-mouse or with luciferin to assess tumor burden and localization by
anti-rabbit IgG (Promega) and developed by enhanced che- whole-body bioluminescence imaging. IL-7 WT animals dis-
miluminescence (Amersham-Pharmacia). played strikingly stronger and more disseminated luciferase
activity (Fig. 1A, left). Quantification of bioluminescence
Quantitative reverse transcriptase PCR intensity confirmed that the tumor burden was significantly
Total RNA (1 mg) was reverse transcribed using the ImProm higher in IL-7 WT mice (Fig. 1A, right). Analysis of sacrificed
II Reverse Transcriptase enzyme (Promega) and random hex- animals showed that the lungs, spleen, liver, kidneys, and bone
amers. The quantitative assessment of IL7RA transcripts was marrow were infiltrated with HPB-ALL.Luc.GFP cells (data
made by SYBR green-based quantitative reverse transcriptase not shown). These data suggested that absence of IL-7 delayed
PCR (qRT-PCR) on an ABI PRISM 7500 (Applied Biosystems) disease progression and dissemination in vivo.
with the following primers: ABL, TGGAGATAACACTCTAAG- To confirm these observations, we used TAIL7 cells, which
CATAACTAAAGGT and GATGTAGTTGCTTGGGACCCA; and are IL-7-dependent in vitro and constitute a model for primary
IL7R, GGATTAAGCCTATCGTAT-GGC (exon 7) and GCTTGA- T-ALL (18). First, we transplanted CFSE-labeled TAIL7 cells into
ATGTCATCCACCCT (exon 8). Standard curves were obtained IL-7 WT and IL-7 KO mice and evaluated engraftment 6 hours
by serial dilutions of PCR products, and IL7RA transcript values after i.v. injection. Analysis of the bone marrow revealed the
were normalized with respect to the number of ABL transcripts. presence of very few leukemia cells, and no difference between
IL-7 WT (36.5 26.7) and IL-7 KO (31.6 16.7) mice (Supple-
IL-7 plasma quantifications mentary Fig. S1). Spleen and several nonlymphoid organs
Human IL-7 levels were quantified in plasma, using the high (lung, liver, and kidney) were also analyzed and all presented
sensitivity IL-7 Quantikine HS Elisa kit (R&D) according to the very few cells with no differences between the 2 mouse strains
manufacturer's instructions. Samples were assayed in duplicate. (data not shown). Our data indicate that IL-7 did not affect the
engraftment and initial homing capacity of T-ALL cells.
Ex vivo assessment of IL-7Ra expression in primary We next evaluated the effect of IL-7 on disease progression.
T-ALL cells The percentage of TAIL7 cells circulating in the peripheral
Primary T-ALL cells were cultured in 24-well plates as 2 blood increased with time and was consistently higher in IL-7
106cells/mL at 37 C with 5% CO2 in RPMI-1640 medium WT mice at all time points analyzed (Fig. 1B). These differ-
supplemented with 10% FBS (Invitrogen Corporation), with ences may have resulted from compromised leukemia expan-
or without 10 ng/mL hIL-7 (Peprotech EC). Immediately sion in the bone marrow of IL-7–deficient mice. To test this
ex vivo (0 hour) and at 24 hours of culture, cells were stained possibility, mice were sacrificed 84 days posttransplantation,
with IL-7Ra-PE (R&D) analyzed by flow cytometry. before disease symptoms were apparent. All IL-7 WT animals
(n ¼ 9) presented clear bone marrow involvement. Most mice
Statistical analysis (6 of 9; 67%) displayed overt leukemia according to the clinical
Log-rank test, Student t test, Mann–Whitney test, and definition (23), presenting more than 20% leukemia cells in the
2-way ANOVA were used to compare data, as appropriate bone marrow. All the remaining IL-7 WT mice showed
(P < 0.05 was considered significant). detectable disease in the bone marrow and were thus con-
sidered preleukemic (3 of 9; 33%), as defined in Materials and
Results Methods (Fig. 1C). In contrast, only 4 of 10 (40%) IL-7 KO mice
were leukemic and 2 of 10 preleukemic (20%; Fig. 1C). The
IL-7 accelerates leukemia expansion and leukemia- remaining 4 IL-7 KO mice did not show detectable signs of
related death in mice xenotransplanted with human bone marrow involvement at this time point or displayed
T-ALL cell lines extremely low leukemia cellularity (Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Silva et al.
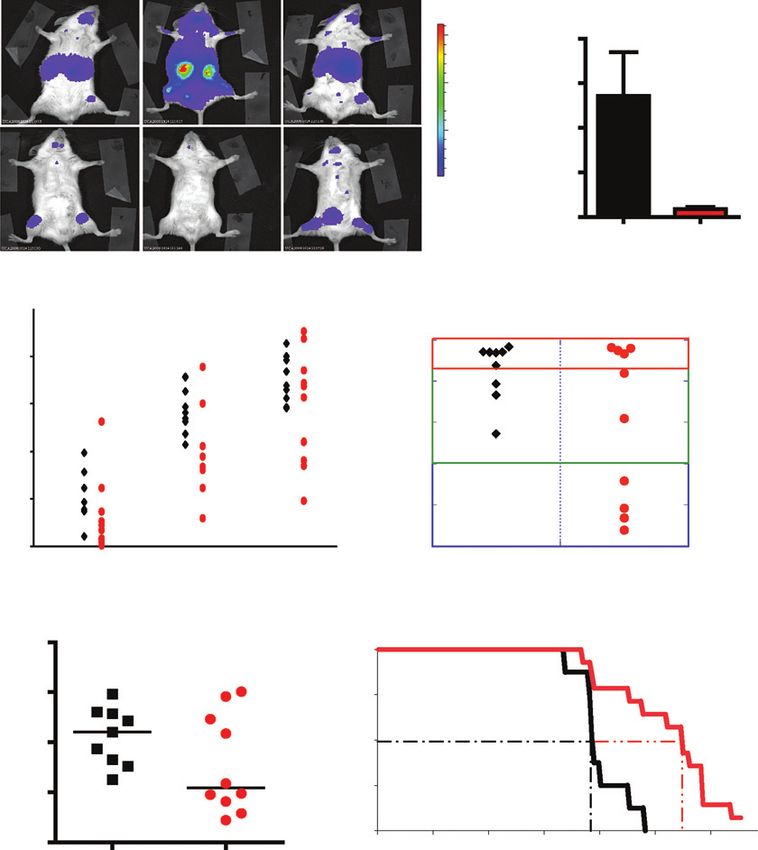
A
I II III
P = 0.0245
60,000 8.0
IL-7 WT
total flux (photons/s) × 108
Bioluminescence
40,000 6.0
20,000 4.0
IV V VI
IL-7 KO
2.0
Color scale
Min = 400 0.0
Max = 64,545
IL-7 WT IL-7 KO
B C
Blood
P = 0.0047 Bone marrow
100
10 Leukemic
Percent huCD2+ cells
Percent huCD2+ cells
10
1
1 Pre-leukemic
0.1
0.1
0.01 0.01 Non-leukemic
0.001 0.001
IL-7 WT IL-7 KO
56 70 84
Days posttransplantation
D E
Spleen
10 100
P = 0.0782
Percent huCD2+ cells
1 P = 0.0023
Percent survival
0.1 50
0.01
0
0.001 0 25 50 75 100 125 150
IL-7 WT IL-7 KO Days (postinjection)
Figure 1. IL-7 accelerates leukemia expansion and leukemia-related death in mice xenotransplanted with human T-ALL cell lines. A, Rag2/Il2rg/ mice,
either Il7þ/þ (IL-7 WT; n ¼ 3) or Il7/ (IL-7 KO; n ¼ 3), were injected i.v. with 107 HPB-ALL.Luc.GFP cells and analyzed 5 weeks posttransplantation. Animals
were injected i.p. with 150 mg luciferin/g 7 minutes before imaging and ventrally scanned for total-body luminescence (total flux), with a medium binning
3-minute exposure, using IVIS Lumina and Living Image software. Images on the left are overlays of the luciferase signal with photographs of each animal.
Corresponding bioluminescence values are represented as mean SEM in the graph on the right (P ¼ 0.0245; Student t test). B–E, IL-7 WT (black; n ¼ 9)
or IL-7 KO (red; n ¼ 10) mice were injected i.v. with 107 TAIL7 cells. B, human leukemia cells were monitored in the blood through time by flow cytometric analysis
with an anti-human CD2 antibody. Differences between IL-7 WT and IL-7 KO groups were statistically significant (P ¼ 0.0047; 2-way ANOVA). C, animals
were sacrificed 84 days posttransplantation, and bone marrow infiltration of TAIL7 cells was evaluated as described in the Materials and Methods. Animals were
classified as leukemic if presenting more than 20% T-ALL cells in the bone marrow, preleukemic if 20% or less, and nonleukemic if less than 0.1%. D,
infiltration of TAIL7 cells in the spleen was analyzed at the same time point (P ¼ 0.0782; Mann–Whitney test). Lines indicate median values of each group.
E, survival of TAIL7-transplanted IL-7 WT (n ¼ 8) and IL-7 KO mice (n ¼ 14) was compared. Kaplan–Meier survival curves are indicated. Disease progression was
significantly accelerated in IL-7 WT mice (P ¼ 0.0023; log-rank test). Experiments in A and E were carried out once. Results in B–D are representative
of 3 independent experiments.
OF4 Cancer Res; 71(14) July 15, 2011 Cancer Research
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
IL-7 Accelerates Human T-ALL In Vivo
A Blood
B Bone marrow
P = 0.0259 50 P = 0.0002
100
Percent huCD5+ cells
40
Percent huCD2+ cells
10
30
1 Figure 2. IL-7 accelerates
20 leukemia expansion in mice
0.1 xenotransplanted with human
IL-7 WT 10 primary T-ALL cells. IL-7 WT
0.01
IL-7 KO (n ¼ 3) and IL-7 KO (n ¼ 4) mice
0 were injected i.v. with 5 106
0.001
56 84 112 IL-7 WT IL-7 KO primary T-ALL cells. A, human
Days posttransplantation T-ALL cells were monitored in the
blood through time by flow
C Spleen Liver cytometric analysis, using an anti-
P = 0.0014 P = 0.0040 human CD2 antibody. Differences
80 40 between IL-7 WT and IL-7 KO
groups were statistically
60 significant (P ¼ 0.0259; 2-way
30
ANOVA). Representative results of
1 of 2 independent experiments
40 20 carried out (each using a different
patient sample) are shown.
Animals were sacrificed 112 days
20 10 posttransplantation and (B) bone
marrow (P ¼ 0.0002; Student t
Percent huCD5+ cells
test), (C) spleen (P ¼ 0.0014), liver
0 0 (P ¼ 0.0040), kidney (P ¼ 0.0873),
IL-7 WT IL-7 KO IL-7 WT IL-7 KO and lung (P ¼ 0.0044) were
disrupted into a cell suspension
Kidney Lung
P = 0.0873 P = 0.0044 and analyzed for primary T-ALL
30 10 cell infiltration by flow cytometry
with an anti-human CD5 antibody.
8 Results indicate mean SEM for
each organ and represent a pool
20
6 from 2 independent experiments
using 2 different patient samples
for a total of 4 IL-7 WT and 6 IL-7
4 KO mice.
10
2
0 0
IL-7 WT IL-7 KO IL-7 WT IL-7 KO
with malignant blasts (Fig. 1D). The nonlymphoid organs mentary Fig. S2). Importantly, we found that the median
analyzed (liver, kidney, and lungs) also presented leukemia survival of TAIL7-transplanted IL-7 WT mice (97 days) was
infiltration (Supplementary Fig. S2). However, IL-7 KO mice significantly shorter than that of IL-7 KO mice (138 days; P ¼
consistently displayed a more heterogeneous distribution and 0.0023; log-rank test; Fig. 1E). Postmortem analysis showed
lower median values of malignant infiltration in all lymphoid that all animals died of leukemia with multiple organ infiltra-
and nonlymphoid organs analyzed (Fig. 1C and D; Supple- tion (Supplementary Fig. S3). Hence, although our data
Figure 3. IL-7 promotes p27Kip1 downregulation, Bcl-2 upregulation, proliferation, and viability of xenotransplanted human primary T-ALL cells. IL-7 WT and
IL-7 KO mice were injected i.v. with 5 106 primary T-ALL cells from 2 distinct patients in independent experiments. Animals were sacrificed 112 days
posttransplantation and primary T-ALL cells in the bone marrow were discriminated by flow cytometry with anti-human CD5 antibodies. A, Bcl-2 protein levels
were assessed by flow cytometry after intracellular staining of primary T-ALL cells (patient 1) with FITC-conjugated anti–Bcl-2 antibody. Left, results are
representative of both patients analyzed. Specific MIF, as described in Materials and Methods, is indicated in the histogram for each condition. Right, Bcl-2
levels in T-ALL cells collected from IL-7 WT and IL-7 KO mice (P ¼ 0.0006; Student t test). Results indicate mean SEM and represent a pool from
the 2 different patient samples for a total of 4 IL-7 WT and 6 IL-7 KO mice. B, primary T-ALL cells (patient 1) recovered from IL-7 WT (n ¼ 3) and IL-7 KO (n ¼ 2)
mice were stained with Annexin V–PE and viability was determined by flow cytometry. Left, a representative histogram overlay is shown with the percentage
of Annexin V–negative cells indicated. Right, percentage of Annexin V–negative T-ALL cells collected from IL-7 WT and IL-7 KO mice (P ¼ 0.0514).
Results represent a pool from the 2 different patient samples for a total of 4 IL-7 WT and 2 IL-7 KO mice. C, total bone marrow cells from IL-7 WT (n ¼ 5) and IL-7
KO (n ¼ 6) mice were lysed, resolved by 10% SDS-PAGE, and immunoblotted with anti-p27Kip1 antibody. Left, membrane was stripped and reprobed
www.aacrjournals.org Cancer Res; 71(14) July 15, 2011 OF5
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Silva et al.
A P = 0.0006
100 50
IL-7 KO 24.1
Bcl-2 expression (MIF)
IL-7 WT 41.1
80 40
in huCD5+ cells
Cell number
60 30
40 20
20 10
0 0
100 101 102 103 104
IL-7 WT IL-7 KO
Bcl-2
B 100 P = 0.0514
Percent Annexin V–negative
100
IL-7 KO 30.0
IL-7 WT 70.6 80
80
huCD5+ cells
Cell number
60
60
40 40
20 20
0 0
100 101 102 103 104
IL-7 WT IL-7 KO
Annexin V
C 1.2 P = 0.0260
p27Kip1 expression (a.u.)
IL-7 WT IL-7 KO TAIL7 1.0
1 2 3 4 5 6 7 8 9 10 11 12 13
0.8
p27Kip1
0.6
Actin
0.4
0.2
0.0
IL-7 WT IL-7 KO
D
P = 0.0121
100
100
IL-7 KO 496
Percent K-i67–positive
IL-7 WT 934 80
80
huCD5+ cells
Cell number
60
60
40 40
20 20
0 0
100 101 102 103 104 IL-7 WT IL-7 KO
Ki-67
with actin to confirm equal loading. Patient 1, lanes 1, 2, 6, and 7; patient 2, lanes 3 to 5 and 8 to 11; total lysates of TAIL7 cells were used as a positive control.
Right, corresponding densitometric analysis of p27Kip1 levels, normalized to actin loading control and expressed in arbitrary units (a.u.), in IL-7 KO and
IL-7 WT mice (P ¼ 0.0260). D, Ki-67 protein levels were assessed by flow cytometry after intracellular staining of primary T-ALL cells with PE-conjugated
anti–Ki-67 antibody. Left, a representative histogram overlay is shown. Right, percentage of Ki-67–positive T-ALL cells collected from IL-7 WT and IL-7 KO
mice (P ¼ 0.0121). Results represent a pool from the 2 different patient samples for a total of 4 IL-7 WT and 6 IL-7 KO mice.
OF6 Cancer Res; 71(14) July 15, 2011 Cancer Research
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
IL-7 Accelerates Human T-ALL In Vivo
indicate that lack of IL-7 is not sufficient to prevent leukemia human T-ALL cells respond to and benefit significantly from
progression and leukemia-related death, IL-7 clearly contrib- the presence of IL-7 in vivo. We collected evidence directly
uted significantly to leukemia acceleration in vivo. from patient specimens at diagnosis supporting this notion.
Both IL-7 KO and IL-7 WT mice lack g-chain, which is We found that T-ALL cells had significantly lower IL7RA
essential for efficient IL-7–mediated signaling by forming mRNA levels than normal T-cell precursors (Fig. 4A) and that
the IL-7R together with IL-7Ra (26, 27). Thus, the differences IL-7 levels were decreased in the plasma of T-ALL patients as
observed in leukemia expansion should largely reflect the compared with healthy age-matched controls (Fig. 4B), sug-
direct impact of IL-7 on T-ALL cells, which display both IL-7Ra gesting that T-ALL cells consume IL-7 and thereby down-
and g-chain (11, 28). For confirmation, we transplanted regulate IL-7Ra. In support of this notion, we found that 4 of 6
growth factor–independent, IL-7Ra–negative P12 T-ALL cells patient samples cultured ex vivo in medium without IL-7
and analyzed IL-7 KO and WT mice at 4 weeks. Both strains showed upregulation of IL-7Ra surface expression, which
showed similar signs of leukemia (Supplementary Fig. S4). was prevented by the addition of IL-7 (Fig. 4C). Taking into
account that more than 70% of T-ALL patient samples
IL-7 accelerates leukemia expansion in mice respond to IL-7 ex vivo and express functional IL-7Rs (11,
xenotransplanted with human primary T-ALL cells 28), our current observations suggest that human T-ALL cells
To ask whether the influence of IL-7 was a more general from a majority of patients might be sensitive to IL-7 signaling
feature of T-ALL and not restricted only to leukemia cell lines, in vivo in the actual disease setting.
we xenotransplanted primary T-ALL cells collected from 2
patients at diagnosis, which differed in their developmental Discussion
maturation block (Table 1). According to the EGIL criteria
(22), the samples were arrested at the cortical (patient 1) and There is mounting evidence that microenvironmental fac-
mature (patient 2) thymocyte stage, whereas TAIL7 cells are tors can contribute to tumor progression. IL-7 is produced in
pre-T and HPB-ALL are cortical but differ from patient 1 in the the thymus and bone marrow, the microenvironments where
expression of CD3 (Table 1). Similar to the cell lines, we T-ALL develops and expands. Interestingly, IL-7 has been
observed a longitudinal increase in the percentage of circulat- shown to have oncogenic potential in vivo. Transgenic mice
ing T-ALL cells, with systematically higher levels in the pre- expressing IL-7 in lymphocytes develop B- and T-cell lympho-
sence of IL-7 (Fig. 2A). In addition, IL-7 WT mice culled mas (1) as a consequence of an autocrine loop involving IL-7
112 days posttransplantation presented significantly higher stimulation via IL-7R (2). However, there is no evidence that
leukemia involvement in the bone marrow than IL-7 KO human T-ALL cells express IL-7, largely excluding the exis-
animals (Fig. 2B). Furthermore, absence of IL-7 clearly tence in this malignancy of an IL-7–dependent autocrine loop
impaired infiltration of organs such as spleen, liver, kidney, that has been suggested to play a role in other hematologic
and lungs (Fig. 2C). cancers (32, 33). Thus, in the present study, we explored the
alternative hypothesis that IL-7 produced by the microenvir-
IL-7 promotes p27Kip1 downregulation, Bcl-2 onment may promote human T-ALL expansion in vivo.
upregulation, proliferation, and viability of Our data indicate that IL-7 does not affect the engraftment
xenotransplanted human primary T-ALL cells and initial homing capacity of T-ALL cells to the bone marrow
We previously showed that IL-7 promotes survival and cell- or other organs. However, it remains to be evaluated whether
cycle progression of T-ALL cells in vitro via Bcl-2 upregulation the late differences between IL-7–deficient and IL-replete
and p27Kip1 downregulation (12, 13). We sought to understand mice regarding leukemia infiltration into organs such as
whether the same mechanisms and functional effects were spleen and liver reflect modulation of leukemia cell trafficking
responsible for the accelerated leukemia progression in the from the bone marrow to these organs and/or of malignant
presence of IL-7 in vivo. We collected T-ALL cells from the bone cell proliferation within each infiltrated organ.
marrow of mice sacrificed at 112 days and analyzed them for We found that lack of IL-7 is not sufficient to prevent
viability and Bcl-2 expression. Primary leukemia cells collected leukemia progression and leukemia-related death, such that
from IL-7 WT animals displayed significantly higher Bcl-2 levels all animals died of leukemia irrespective of IL-7. This may
(Fig. 3A) and increased percentage of viable cells (Fig. 3B), in reflect the acquisition of growth factor independence or the
agreement with the observation that IL-7 transgenic mice pre- effect of other microenvironmental stimuli (e.g., g C-signaling
sent upregulation of Bcl-2 protein (29). Conversely, p27Kip1 levels cytokines (11), Notch ligands (34), or chemokines such as
were downregulated in T-ALL cells recovered from IL-7 WT mice CCL25 and CXCL13 (35, 36) that may provide T-ALL cells
(Fig. 3C), which presented higher Ki-67 expression, indicative of with alternative growth signals in the absence of IL-7. This
increased in vivo proliferation (Fig. 3D). notwithstanding, IL-7 clearly contributes significantly to leu-
kemia acceleration in vivo. Curiously, despite the evidence that
Leukemia cells in T-ALL patients appear to respond to mouse and human IL-7 have similar in vitro bioactivity toward
and consume IL-7 human lymphocytes (24, 25), including T-ALL cells (25), it was
IL-7 circulating levels are regulated by consumption (30) recently shown that in vivo IL-7 present in the mouse is less
and IL-7 signaling leads to the transcriptional downregulation active on human cells (37). This suggests that the effect of IL-7
of IL-7Ra (31). Although cancer cells may grow independently on human leukemia progression in the RAG KO xenotrans-
of external stimuli, our data argue that a significant fraction of plant models we used may constitute an underestimation of
www.aacrjournals.org Cancer Res; 71(14) July 15, 2011 OF7
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Silva et al.
questions of whether IL-7 promotes survival of a putative
A leukemia stem cell population and participates in T-ALL
1.5 initiation remain to be scrutinized.
P = 0.0002
At the molecular level, we found that IL-7–related disease
progression was associated with decreased p27Kip1 expression
(normalized to ABL)
IL-7R expression
1.0 and upregulated Bcl-2. Given our previous demonstration that
regulation of p27Kip1 and Bcl-2 is mandatory for IL-7–mediated
antiapoptotic and proliferative effects on T-ALL cells in vitro
(13), it is reasonable to infer that the same mechanisms
0.5 underlie T-ALL expansion in vivo in response to IL-7. In support
of this possibility, it is noteworthy that T-ALL patient samples
frequently display p27Kip1 downregulation (39) and Bcl-2
0.0 upregulation (40). Moreover, most primary T-ALL samples
T-ALL Thymocytes show PI3K pathway activation (17), which may be the con-
sequence not only of cell autonomous mechanisms (17, 41, 42)
B 2.0
but also of microenvironmental stimuli such as IL-7 (12). The
P = 0.016 importance of IL-7/IL-7R signaling is further illustrated by the
IL-7 plasma levels (pg/mL)
observations that in vitro IL-7 responsiveness (28) and IL7RA
1.5 gene expression (43) appear to have prognostic value in T-ALL.
Notably, our studies indicate that the in vivo impact of IL-7 on
1.0 T-ALL cells is not restricted to a particular maturation stage or
CD4/CD8 immunophenotype (Table 1). These observations are
0.5 reminiscent of our in vitro findings, in that T-ALL patient
samples proliferated in response to IL-7 irrespective of the
stage at which they were developmentally blocked (11).
0.0
Altogether, our data provide the first clear evidence that
human T-ALL cells utilize the IL-7/IL-7R axis for expansion in
T-ALL CTRL vivo, providing experimental support to the hypothesis that
C responsiveness to a g C-signaling cytokine contributes to
100 Ex vivo (o h)
T-ALL development in vivo (5, 44). Because the majority of
+ IL-7 (24 h) T-ALL cases are known to respond to IL-7 in vitro (11) and
80 – IL-7 (24 h) thus possibly benefit from IL-7 stimulation in vivo, therapeutic
approaches that integrate targeting IL-7, IL-7R, or key ele-
Percent Max
60 ments in IL-7/IL-7R–mediated signaling into current proto-
cols may prove beneficial for the treatment of T-ALL. In
40 addition, our observations confirm the need for caution when
administering IL-7 in the context of cancer immunotherapy
20 (45) and suggest that the promise that IL-7 holds as an
immunorestorative (46, 47) and in enhancing immunologic
responses against cancer (48, 49) should be further evaluated
0
strictly in patients suffering from cancers that are well char-
100 101 102 103 104
acterized as refractory to the cytokine.
IL-7Rα expression
Disclosure of Potential Conflicts of Interest
Figure 4. Ex vivo evidence for leukemia cell responsiveness to IL-7 in No potential conflicts of interest were disclosed.
T-ALL patients. A, RNA was collected from primary T-ALL samples (n ¼ 12)
and normal control thymocytes (n ¼ 9), and qRT-PCR was carried out
to determine IL-7Ra mRNA expression levels (P ¼ 0.0002, unpaired 2- Acknowledgments
tailed Student t test). B, IL-7 plasma levels in samples collected from T-ALL
patients (n ¼ 7) or age-matched healthy controls (n ¼ 8) were determined We thank Dr. Paulo Lúcio for kindly providing the immunophenotypic
using an IL-7 ELISA (P ¼ 0.016). C, IL-7Ra expression in T-ALL cells was analysis of primary patient samples, Dr. Miguel Abecasis for generously provid-
ing the thymic specimens, and Daniel Ribeiro, Nadia Correia, Jaíra Ferreira de
evaluated by flow cytometry ex vivo and after 24 hours of in vitro culture
Vasconcellos, and Priscila P. Zenatti for their invaluable help. We especially
with or without IL-7 (10 ng/mL). Histogram overlay is representative of 4 of acknowledge the contribution of patients and their families, and Dr. Mario
6 (67%) patients analyzed. Chagas and all the physicians and nurses at the Pediatric Service of Instituto
Portugu^es de Oncologia de Lisboa, in providing primary samples.
the impact of IL-7 on human T-ALL patients. Moreover,
Grant Support
because cancer cells are generally viewed as nonreliant on
growth factor signals (38), the fact that IL-7 accelerated This work was supported by grants from Associaç~ao Portuguesa Contra a
disease progression is especially remarkable. The related Leucemia, FCT (POCI/SAU-OBS/58913 and PTDC/SAU-OBD/104816), and
OF8 Cancer Res; 71(14) July 15, 2011 Cancer Research
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
IL-7 Accelerates Human T-ALL In Vivo
Children with Leukaemia Charity, UK, to J.T. Barata; FAPESP (08/10034-1) to J.A. advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate
Yunes, Medical Research Council, UK, program code U117573801 to B. Seddon. this fact.
A. Silva, L.R. Martins, and B.A. Cardoso had PhD fellowships from FCT-SFRH.
A.B.A. Laranjeira had a PhD fellowship from FAPESP.
The costs of publication of this article were defrayed in part by the Received October 5, 2010; revised March 16, 2011; accepted May 12, 2011;
payment of page charges. This article must therefore be hereby marked published OnlineFirst May 18, 2011.
References
1. Rich BE, Campos-Torres J, Tepper RI, Moreadith RW, Leder P. 19. DSMZ Human and Animal Cell Lines [cited 2011 June 27]. Available
Cutaneous lymphoproliferation and lymphomas in interleukin 7 trans- from: http://www.dsmz.de/human_and_animal_cell_lines/
genic mice. J Exp Med 1993;177:305–16. 20. Amendola M, Venneri MA, Biffi A, Vigna E, Naldini L. Coordinate dual-
2. Abraham N, Ma MC, Snow JW, Miners MJ, Herndier BG, Goldsmith gene transgenesis by lentiviral vectors carrying synthetic bidirectional
MA. Haploinsufficiency identifies STAT5 as a modifier of IL-7-induced promoters. Nat Biotechnol 2005;23:108–16.
lymphomas. Oncogene 2005;24:5252–7. 21. Henriques CM, Rino J, Nibbs RJ, Graham GJ, Barata JT. IL-7 induces
3. Laouar Y, Crispe IN, Flavell RA. Overexpression of IL-7R alpha rapid clathrin-mediated internalization and JAK3-dependent degra-
provides a competitive advantage during early T-cell development. dation of IL-7Ralpha in T cells. Blood 2010;115:3269–77.
Blood 2004;103:1985–94. 22. Bene MC, Castoldi G, Knapp W, Ludwig WD, Matutes E, Orfao A, et al.
4. Jiang Q, Li WQ, Aiello FB, Mazzucchelli R, Asefa B, Khaled AR. Cell Proposals for the immunological classification of acute leukemias.
biology of IL-7, a key lymphotrophin. Cytokine Growth Factor Rev European Group for the Immunological Characterization of Leukemias
2005;16:513–33. (EGIL). Leukemia 1995;9:1783–6.
5. Barata JT, Cardoso AA, Boussiotis VA. Interleukin-7 in T-cell acute 23. Barabe F, Kennedy JA, Hope KJ, Dick JE. Modeling the initiation and
lymphoblastic leukemia: an extrinsic factor supporting leukemogen- progression of human acute leukemia in mice. Science 2007;316:
esis? Leuk Lymphoma 2005;46:483–95. 600–4.
6. Mazzucchelli RI, Warming S, Lawrence SM, Ishii M, Abshari M, 24. Johnson SE, Shah N, Panoskaltsis-Mortari A, LeBien TW. Murine and
Washington AV, et al. Visualization and identification of IL-7 producing human IL-7 activate STAT5 and induce proliferation of normal human
cells in reporter mice. PLoS One 2009;4:e7637. pro-B cells. J Immunol 2005;175:7325–31.
7. Alves NL, Richard-Le Goff O, Huntington ND, Sousa AP, Ribeiro VS, 25. Barata JT, Silva A, Abecasis M, Carlesso N, Cumano A, Cardoso AA.
Bordack A, et al. Characterization of the thymic IL-7 niche in vivo. Proc Molecular and functional evidence for activity of murine IL-7 on human
Natl Acad Sci U S A 2009;106:1512–7. lymphocytes. Exp Hematol 2006;34:1133–42.
8. Scupoli MT, Vinante F, Krampera M, Vincenzi C, Nadali G, Zampieri F, 26. Kondo M, Takeshita T, Higuchi M, Nakamura M, Sudo T, Nishikawa S,
et al. Thymic epithelial cells promote survival of human T-cell acute et al. Functional participation of the IL-2 receptor gamma chain in IL-7
lymphoblastic leukemia blasts: the role of interleukin-7. Haematolo- receptor complexes. Science 1994;263:1453–4.
gica 2003;88:1229–37. 27. Ziegler SE, Morella KK, Anderson D, Kumaki N, Leonard WJ, Cosman
9. Scupoli MT, Perbellini O, Krampera M, Vinante F, Cioffi F, Pizzolo G. D, et al. Reconstitution of a functional interleukin (IL)-7 receptor
Interleukin 7 requirement for survival of T-cell acute lymphoblastic demonstrates that the IL-2 receptor gamma chain is required for
leukemia and human thymocytes on bone marrow stroma. Haema- IL-7 signal transduction. Eur J Immunol 1995;25:399–404.
tologica 2007;92:264–6. 28. Karawajew L, Ruppert V, Wuchter C, Ko € sser A, Schrappe M, Do€ rken
10. Touw I, Pouwels K, van Agthoven T, van Gurp R, Budel L, Hooger- B, et al. Inhibition of in vitro spontaneous apoptosis by IL-7 correlates
brugge H, et al. Interleukin-7 is a growth factor of precursor B and T with bcl-2 up-regulation, cortical/mature immunophenotype, and
acute lymphoblastic leukemia. Blood 1990;75:2097–101. better early cytoreduction of childhood T-cell acute lymphoblastic
11. Barata JT, Keenan TD, Silva A, Nadler LM, Boussiotis VA, Cardoso AA. leukemia. Blood 2000;96:297–306.
Common gamma chain-signaling cytokines promote proliferation of T- 29. El Kassar N, Lucas PJ, Klug DB, Zamisch M, Merchant M, Bare CV,
cell acute lymphoblastic leukemia. Haematologica 2004;89:1459–67. et al. A dose effect of IL-7 on thymocyte development. Blood
12. Barata JT, Silva A, Brandao JG, Nadler LM, Cardoso AA, Boussiotis 2004;104:1419–27.
VA. Activation of PI3K is indispensable for interleukin 7-mediated 30. Guimond M, Veenstra RG, Grindler DJ, Zhang H, Cui Y, Murphy RD,
viability, proliferation, glucose use, and growth of T cell acute lym- et al. Interleukin 7 signaling in dendritic cells regulates the homeostatic
phoblastic leukemia cells. J Exp Med 2004;200:659–69. proliferation and niche size of CD4þ T cells. Nat Immunol 2009;10:
13. Barata JT, Cardoso AA, Nadler LM, Boussiotis VA. Interleukin-7 149–57.
promotes survival and cell cycle progression of T-cell acute lympho- 31. Park JH, Yu Q, Erman B, Appelbaum JS, Montoya-Durango D, Grimes
blastic leukemia cells by down-regulating the cyclin-dependent HL, et al. Suppression of IL7Ralpha transcription by IL-7 and other
kinase inhibitor p27(kip1). Blood 2001;98:1524–31. prosurvival cytokines: a novel mechanism for maximizing IL-7-depen-
14. Brown VI, Fang J, Alcorn K, Barr R, Kim JM, Wasserman R, et al. dent T cell survival. Immunity 2004;21:289–302.
Rapamycin is active against B-precursor leukemia in vitro and in vivo, 32. Cattaruzza L, Gloghini A, Olivo K, Di Francia R, Lorenzon D, De Filippi
an effect that is modulated by IL-7-mediated signaling. Proc Natl Acad R, et al. Functional coexpression of Interleukin (IL)-7 and its receptor
Sci U S A 2003;100:15113–8. (IL-7R) on Hodgkin and Reed-Sternberg cells: Involvement of IL-7 in
15. Williams RT, Roussel MF, Sherr CJ. Arf gene loss enhances onco- tumor cell growth and microenvironmental interactions of Hodgkin's
genicity and limits imatinib response in mouse models of Bcr-Abl- lymphoma. Int J Cancer 2009;125:1092–101.
induced acute lymphoblastic leukemia. Proc Natl Acad Sci U S A 33. Nakayama J, Yamamoto M, Hayashi K, Satoh H, Bundo K, Kubo M,
2006;103:6688–93. et al. BLNK suppresses pre-B-cell leukemogenesis through inhibition
16. Williams RT, den Besten W, Sherr CJ. Cytokine-dependent imatinib of JAK3. Blood 2009;113:1483–92.
resistance in mouse BCR-ABLþ, Arf-null lymphoblastic leukemia. 34. Armstrong F, Brunet de la Grange P, Gerby B, Rouyez MC, Calvo J,
Genes Dev 2007;21:2283–7. Fontenay M, et al. NOTCH is a key regulator of human T-cell acute
17. Silva A, Yunes JA, Cardoso BA, Martins LR, Jotta PY, Abecasis M, leukemia initiating cell activity. Blood 2009;113:1730–40.
et al. PTEN posttranslational inactivation and hyperactivation of the 35. Qiuping Z, Qun L, Chunsong H, Xiaolian Z, Baojun H, Mingzhen Y,
PI3K/Akt pathway sustain primary T cell leukemia viability. J Clin et al. Selectively increased expression and functions of chemokine
Invest 2008;118:3762–74. receptor CCR9 on CD4þ T cells from patients with T-cell lineage acute
18. Barata JT, Boussiotis VA, Yunes JA, Ferrando AA, Moreau LA, Veiga lymphocytic leukemia. Cancer Res 2003;63:6469–77.
JP, et al. IL-7-dependent human leukemia T-cell line as a valuable tool 36. Qiuping Z, Jie X, Youxin J, Qun W, Wei J, Chun L, et al. Selectively
for drug discovery in T-ALL. Blood 2004;103:1891–900. frequent expression of CXCR5 enhances resistance to apoptosis in
www.aacrjournals.org Cancer Res; 71(14) July 15, 2011 OF9
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
Silva et al.
CD8(þ)CD34(þ) T cells from patients with T-cell-lineage acute lym- T-cell acute lymphoblastic leukemia: a Children's Oncology Group
phocytic leukemia. Oncogene 2005;24:573–84. study. Mol Cancer 2010;9:105.
37. van Lent AU, Dontje W, Nagasawa M, Siamari R, Bakker AQ, Pouw 44. McCormack MP, Rabbitts TH. Activation of the T-cell oncogene
SM, et al. IL-7 enhances thymic human T cell development in "human LMO2 after gene therapy for X-linked severe combined immunode-
immune system" Rag2/IL-2Rgammac/ mice without affecting ficiency. N Engl J Med 2004;350:913–22.
peripheral T cell homeostasis. J Immunol 2009;183:7645–55. 45. Capitini CM, Chisti AA, Mackall CL. Modulating T-cell homeostasis
38. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100: with IL-7: preclinical and clinical studies. J Intern Med 2009;266:141–
57–70. 53.
39. Wolfraim LA, Fernandez TM, Mamura M, Fuller WL, Kumar R, Cole DE, 46. Sportes C, Hakim FT, Memon SA, Zhang H, Chua KS, Brown MR, et al.
et al. Loss of Smad3 in acute T-cell lymphoblastic leukemia. N Engl J Administration of rhIL-7 in humans increases in vivo TCR repertoire
Med 2004;351:552–9. diversity by preferential expansion of naive T cell subsets. J Exp Med
40. Coustan-Smith E, Kitanaka A, Pui CH, McNinch L, Evans WE, Rai- 2008;205:1701–14.
mondi SC, et al. Clinical relevance of BCL-2 overexpression in child- 47. Sportes C, Babb RR, Krumlauf MC, Hakim FT, Steinberg SM, Chow
hood acute lymphoblastic leukemia. Blood 1996;87:1140–6. CK, et al. Phase I study of recombinant human interleukin-7 admin-
41. Palomero T, Sulis ML, Cortina M, Real PJ, Barnes K, Ciofani M, et al. istration in subjects with refractory malignancy. Clin Cancer Res
Mutational loss of PTEN induces resistance to NOTCH1 inhibition in 2010;16:727–35.
T-cell leukemia. Nat Med 2007;13:1203–10. 48. Li B, VanRoey MJ, Jooss K. Recombinant IL-7 enhances the potency
42. Gutierrez A, Sanda T, Grebliunaite R, Carracedo A, Salmena L, Ahn Y, of GM-CSF-secreting tumor cell immunotherapy. Clin Immunol
et al. High frequency of PTEN, PI3K, and AKT abnormalities in T-cell 2007;123:155–65.
acute lymphoblastic leukemia. Blood 2009;114:647–50. 49. Pellegrini M, Calzascia T, Elford AR, Shahinian A, Lin AE, Dissanayake D,
43. Cleaver AL, Beesley AH, Firth MJ, turges NC, O’Leary RA, Hunger SP, et al. Adjuvant IL-7 antagonizes multiple cellular and molecular inhibitory
et al. Gene-based outcome prediction in multiple cohorts of pediatric networks to enhance immunotherapies. Nat Med 2009;15:528–36.
OF10 Cancer Res; 71(14) July 15, 2011 Cancer Research
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.Published OnlineFirst May 18, 2011; DOI: 10.1158/0008-5472.CAN-10-3606
IL-7 Contributes to the Progression of Human T-cell Acute
Lymphoblastic Leukemias
Ana Silva, Angelo B.A. Laranjeira, Leila R. Martins, et al.
Cancer Res Published OnlineFirst May 18, 2011.
Updated version Access the most recent version of this article at:
doi:10.1158/0008-5472.CAN-10-3606
Supplementary Access the most recent supplemental material at:
Material http://cancerres.aacrjournals.org/content/suppl/2011/05/18/0008-5472.CAN-10-3606.DC1
E-mail alerts Sign up to receive free email-alerts related to this article or journal.
Reprints and To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Subscriptions Department at pubs@aacr.org.
Permissions To request permission to re-use all or part of this article, use this link
http://cancerres.aacrjournals.org/content/early/2011/07/01/0008-5472.CAN-10-3606.
Click on "Request Permissions" which will take you to the Copyright Clearance Center's
(CCC)
Rightslink site.
Downloaded from cancerres.aacrjournals.org on January 20, 2021. © 2011 American Association for Cancer
Research.You can also read

















































