Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia - De ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Open Vets. 2021; 2: 40–54
Communication
Fatik Baran Mandal, Ph.D.
Hurdle in taxonomy: A case of malaria parasites
and other Haemosporidia
https://10.1515/ovs-2020-0110 are: morphological, cytological, biochemical, ecological,
Received Mar 29, 2021; accepted Jul 13, 2021 and ethological. Morpho taxonomy is called the
traditional taxonomy. Each approach of taxonomy has
Abstract: A crucial progress in taxonomy matches
own advantages and drawbacks. However, there is a
with the growth of various branches of biological
conflict between the traditional taxonomy and molecular
sciences. This validates the taxonomic positions of
level systematics. The progress of molecular biology and
many organisms. Haemoproteus, Leucocytozoon and
introducing of new tools and techniques help in finalizing
Plasmodium, the members of the Haemosporidia, are
the taxonomic position of the organism along with its
the well recognized parasitic genera. Revisiting the
evolutionary history. Taxonomy refers to the classification
progress in animal taxonomy appears to be important to
of groups of organisms. Systematics arranges various
evaluate our studies. Haemosporidia being microscopic,
taxa like phyla, subphyla, classes, orders, suborders etc.
their taxonomy specially the morphotaxonomy has
following a hierarchy of evolutionary interrelated groups.
sometimes created confusion. Therefore, analysis of
Taxonomy puts names for organisms and groups of
progression of the taxonomy of the avian Haemosporidia
related organisms. Systematics deals with the taxonomy
demands special attention. Modern phylogenetic analyses
and evolution of organisms to establish an evolutionary
revealed a wealth of information, which is undoubtedly
relationship between taxa meaningfully. Thus, both the
useful for protozoology and other related sciences.
taxonomy and systematic depend on each other.
Techniques of molecular taxonomy are applied to draw
Zoologists took the role of principal taxonomists
the phylogeny covering members of Haemosporidia. The
for protozoa in the early period. Definitely, they had
study of the life cycles of both hosts and parasites are
less opportunity for studying the taxonomic position of
absorbing. Besides, analysis of their evolution through
protozoans with the traditional tools and techniques. Also,
molecular phylogenetics appears to be vital in studying
authoritarianism by the zoologists perhaps played a major
haemosporidians and to gain insight for basic and
role in fixing the systematic position of the lower fungi,
applied sciences. This article examines the potential of
protozoa, and algae at the then time [1]. Now Protozoa are
molecular phylogenetics in refining systematics of avian
not the sole domain of protozoologists. Members of the
Haemosporidia and to explain a holistic view of the group.
Protozoa have important implications in the medical and
veterinary parasitology as well as in microbiology. Gradual
Keywords: Phylogenetics, Hemoproteus, Plasmodium,
development of the classification scheme and progress of
cytochrome b
biochemistry, genetics and molecular biology improved
our knowledge of protozoa and to fill up the gaps in our
knowledge thereof. Many taxonomists still work with
1 Introduction traditional techniques of protozoa. The classification of
protozoa was complicated due to lumping of protozoa,
with algae and some other eukaryotic microorganisms
Systematics, taxonomy, classification, synonyms, and
together as Protista and by data collected from modern
homonyms are becoming difficult for students, teachers,
biological techniques. For a more detailed account in this
and researchers. This applies to the field of zoology,
context, the reader may consult the classical works [2-8].
microbiology, parasitology, medical parasitology,
Similarly, improvements in microscopic methods and
entomology and medical entomology. Difficult situations
molecular biology revealed the morphological, cellular
sometimes arise in dealing with taxa like Protozoa,
and subcellular characteristics of protozoa in detail.
Rotifera and Insecta. Various approaches to taxonomy
Open Access. © 2021 Fatik Baran Mandal, Ph.D., published by De Gruyter. This work is licensed under the Creative Commons Attribution
4.0 Public License. Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 41
The efforts of scientists throughout the globe through identification of specimens. The process of identification
centuries have developed the present-day classification relies on information about the characters of the concerned
of the protozoa. Interesting history of those efforts would organism. Specific taxonomic approach (morphological/
also attract readers. ecological/ behavioral/cytological) requires specific
Here, I review the taxonomy of protozoa, especially information to classify organisms into groups. Taxon is
the avian Haemosporidia to present the complexity of considered as a group of biological organisms (taxa, plural).
the assemblages of diverse groups of animals within Taxa in a classification have rank, with the basic rank (basal
the group. I shall also deal with the brief descriptions, rank) as species. The higher ranks include genus, subgenus,
general characteristics, life cycles, and systematics of the tribe, family, order, suborder, class, subclass, phylum, and
three genera of avian Haemosporidia. Other groups of kingdom. Scientific names are the primary but critical keys to
animals involved in systematics of avian Haemosporidia unlock biosystematics information about living organisms.
will also be taken into account, as and when required, Scientific names are unique. Typically, only one valid
for the sake of clarity. Then I shall discuss the present scientific name indicates a specific organism throughout the
trend of research on avian Haemosporidia as a case world. Disagreements in classification are due to differences
study to illustrate how the hurdles in morphotaxonomy in paradigm and/or information. Theoretically, there should
are gradually decreasing along with the development of be only one correct classification. However, in practice there
biological tools and techniques. were multiple classifications in the past; some of which
are in use today. For the correct classification, we must
2 Methods develop a nomenclatural system that supports two unique
keys (valid name and original name). The valid name is the
correct name and the best name to provide the full value of
While consulting literature on animal classification,
scientific name for a taxon. The original name refers to the
opportunity presented itself to examine the progress in
valid name in the classification in which it was proposed,
taxonomy and to realize how sometimes, synonyms and
which would be useful only to retrieve information across
homonyms create confusion to get the valid information
multiple classifications.
of a genus, or a taxon. Besides, I had to consult the
Classifications also include terms like synonymy
literature on taxonomy of avian haemoparasites or blood
which indicate two or more names for the same species.
inhabiting parasites especially of the genus Haemproteus
Homonymy shows various species having the same
(Haemosporidia: Haemosporina) during 1986-1991 for my
name. However, the rules of nomenclature can solve
PhD work. At that time morpho-taxonomy was mostly
such conflicts. For example, if homonymy becomes an
followed. In the present study, I have analyzed the trends
obstacle, the rule of uniqueness can address it.The rules of
of progress of taxonomy and systematics of the avian
typification can solve the problem of synonymy following
Haemosporidia from my experience of consultation of
the convention. The scientific name to be used is settled by
classic literature, as well as the survey of recent literature
giving priority to the oldest name. The codes of zoological
in the field following standard procedure.
nomenclature can solve the problems of homonymy
Ethical approval: The conducted research is not related
and synonymy, and can guide us for proper formation
to either human or animals use.
and documentation of scientific names. Linnaeus and
Fabricius, following Aristotelian essentialism/typology,
3 Findings developed the first classification. Organisms are grouped
together as they share the criteria of the group, which
The findings are given in subheadings and in sequence as is the type. Later, to develop systematics and to get a
below for convenience. holistic view, evolution was added to the taxonomy.
The classifications then lead to phylogeny, which is
the genealogical hypothesis of relationship. Later,
3.1 Systematics and taxonomy various ways of deciphering phylogeny and translating
phylogenetic information into a hierarchical classification
The science of classification of organisms following the valid (phylogenetics vs cladistic methods) have developed. A
rules is called taxonomy. We get a picture of biodiversity, perusal of literature reveals that much has been written
and the information required for entire biology from the about the relative advantages of phenetics, evolutionary
taxonomy. Taxonomy mostly depends on the accurate systematics, and cladistics [2-8].42 Fatik Baran Mandal, Ph.D.
The International Code of Zoological Nomenclature [11]. Identification of Protozoa has employed microscopic
(ICZN) in its 4th edition consists mainly of a series of methods by tradition. Now molecular methods identify
18 chapters comprising 90 articles. The ICZN contains many cryptic protozoan species. The cryptic species
recommendations and examples to address the conflicts indicates two or more species that were grouped as a single
or misunderstandings in taxonomy. Five different codes nominal species due to their apparent morphological
of nomenclature are now in use, one each for bacteria, similarities. In recent times, molecular analyzes can
viruses, plants, cultivated plants, and animals. In the distinguish different species within the cryptic species.
early 1990s it was attempted to develop a single code of This clears the controversy in the taxonomic position
nomenclature for the living world which was followed and elucidates the systematics of the protozoa. The use
by a publication of draft BioCode. Use of such BioCode of electron microscopes presented a golden epoch of
is significant in taxonomy and in broader aspects in the research in protozoology in the mid-20th century. Some
study of systematics. Naming establishes a taxon as the parasitic protozoa are ectoparasites, while some are
basic unit of biodiversity research. Giving a proper name endoparasites. There is blood inhabiting, skin and tissue
to a species first can only provide the scope to consider inhabiting and visceral- protozoa. The protozoa had about
the infraspecific entities which are found in various rules four times the number of taxonomic units above the family
of nomenclature. The higher-level classification and the level than the classification scheme developed by Butschli
Tree of Life depend on species. The Linnaean binomial (Honigberg Report of 1964) [12]. There are variations in
nomenclature, often considered as outdated now, still nature, survival strategies, pathogenicity, and immune
allows to maximize the number of named species. This responses among various groups of parasites. Parasites
nomenclature places the species into the Tree of Life inhabit vertebrates like Pisces, Amphibia, Reptilia, Aves,
along with their genus, which is difficult with alternative and Mammalia. Researchers have classified the parasites
non-Linnaean systems. However, with molecular level following their own realization, which sometimes fail
advances and with implementation of the Nagoya to follow the principles of taxonomy and thus vary
Protocol, new challenges are emerging. Taxonomy substantially [13-16]. Besides, the nature of parasites
provides the framework to name, classify, and understand belongs to various taxa like Protozoa, Platyhelminthes,
our natural environment. Besides, every branch of applied Nematoda, and Nematomorpha and even of Insecta varies
science deal with organisms which require accurate extensively.
naming and identification of the organism. Several policy The classification of eukaryotic protists has been in
matters became obstacles in the progress of taxonomy. flux for over two centuries [2] following the 19th century
For example, in the past few decades, the focus was on authors like Owen (kingdom Protozoa, 1858), Hogg
revenue-generating science. The governments allocated (kingdom Primigenum, 1860), and Haeckel (kingdom
more research budgets for applied research work, but much Protista, 1866). In the 1970s, a trend developed to divide
less to basic science research. This affects the taxonomic the group protozoa into several subkingdoms. Since
advancement, with permanent employment positions then, new discoveries of ultrastructure of protists have
being eliminated, or replaced by other disciplines with driven to propose explicit phylogenies for protists. In a
emphasis on applied research [9, 10]. conservative approach, Cavalier-Smith [2] opted to retain
a single kingdom Protozoa including all eukaryotes except
the primitive Archezoa and the four eukaryotic kingdoms
3.2 Systematics and taxonomy of Protozoa (Chromista, Plantae, Fungi, and Animalia). In his elegant
review of 42 pages along with over 150 references, Cavalier
Protozoa were accepted as a subset of the kingdom –Smith [2] stated to raise the taxonomic rank of kingdom
Animalia in the middle of the 20th century. Protozoans Protozoa which was created by the Owen (originally
are single-celled organisms in which a single cell protozoa as a class; Goldfuss, 1818). Cavalier –Smith [2]
performs every function of an individual. So, to designate separated protists from animals and plants and divided the
protozoans as acellular is more meaningful. The term protozoa into 18 phyla namely Apicomplexa, Ciliophora,
protozoon is equivalent to the German word Urthiere that Dinozoa, Entamoebia, Euglenozoa, Haplosporidia,
means primitive animals. Leeuwenhoek observed protozoa Percolozoa, Heliozoa, Mycetozoa, Myxosporidia, Opalozoa,
with microscopes made from simple lenses. Members of Paramyxia, Percolozoa, Radiozoa, Reticulosa, Rhizopoda,
the Protozoa are either free living, or parasitic, ranging Choanozoa, and Mesozoa. Cavalier –Smith also defined
from one micrometer to one, or two millimeters in length protozoa with diagnostic characters as follows: Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 43
Predominantly unicellular, plasmodial or colonial Fundraising to procure tools and establish a good
phagotrophic eukaryotes, wall-less in the trophic taxonomic laboratory is still problematic in numerous
state. Primitively possessing mitochondria and countries, and also in individual research institutes
peroxisomes (unlike Archezoa); when mitochondria and or universities. However, necessary skills enhance the
peroxisomes are both secondarily absent (Parabasalia, marketability of the new generation of taxonomists in
Entamoebia, Lyromonadea, and anaerobic ciliates the job market [18]. Many publications [19-22] show the
only), hydrogenosomes and/or Golgi dictyosomes are hurdle in taxonomy which is being discussed here in the
present instead. Ciliary hairs are never rigid and tubular section 3.4 and 3.5.
(unlike most chromists); haptonema absent (excludes Many novel criteria emerged from the use of electron
nonphotosynthetic [94] haptophytes). Chloroplasts, when microscopy. Those criteria helped in developing kingdom
present (some euglenoids and dinoflagellates only), systems of classification, avoiding the dichotomy
contain neither starch nor phycobilisomes (unlike in of animal-or-vegetable. In 1860 John Hogg[23] first
Plantae), have stacked thylakoids, and usually have three, proposed the kingdom Protoctista to separate them from
rather than two, envelope membranes. Chloroplasts are animals and plants. Ernst Haeckel [24]. in the year 1866
located in the cytosol, never within a smooth periplastid also proposed the third kingdom Protista. Haeckel, a
membrane inside either the lumen of the rough biological giant of the second half of the 19th century,
endoplasmic reticulum or a fourth smooth membrane created the Protista, to contain the microscopic and
(unlike Chromista); ejectisomes never of the double-scroll unicellular organisms. He believed that those organisms
cryptist type (this excludes the cryptist Goniomonas); the placed in the Protista should be from the then existing
few multicellular species have minimal cell differentiation kingdoms containing the macroscopic and multicellular
and altogether lack collagenous connective tissue plants and animals. This evolutionarily based systematic
sandwiched between two dissimilar epithelia (unlike concept was again refined in 1878. Haeckel’s concept was
Animalia). controversial from its inception to now. However, this
Source: Cavalier-Smith [2]. is valuable to focus on the phylogenetic component of
taxonomy and on the microscopic eukaryotic organisms
All 41 species and 6 genera belonging to phylum
called the protists. Attempts were taken in the 20th
Apicomplexa were named between 1826 and 1850. In the
century to improve the systematics of all protists. One of
year 1987, those numbers stood respectively at 4516 and
the major challenges to the workers is to determine ways
339 including 444 species of Haemosporida in 9 genera
of including information from phylogenetic cladograms
[17]. All members of Apicomplexa are parasitic, having
into ranked hierarchical schemes of classification [22,25].
apical complexes to penetrate the host cell which signifies
Corliss’s work [1,4-6] help us immensely in unravelling
the name Apicomplexa. They belong to either the class
the early history of protozoa. Haeckel’s [24] idea of origin
Perkinsea, or class Sporozoea, and exhibit alternation
of protist, animal, and plant independently from various
of generation with cycles of sporogony (sexual) and
precellular ancestors led Woese and Fox [26] to propose
schizogony (asexual cycle) besides gametogony. The
a ‘progenote’ ancestor. Cavalier-Smith [3] considered
gametocytes and sporozoites are the invasive or infecting
Apicomplexa to rank as a subphylum and Sporozoa as an
stages in the life cycle of the members of Apicomplexa.
infraphylum.
Haemosporidians belong to the suborder Haemospororina
Revision of the classification of protozoa called the
in the phylum Apicomplexa of the subkingdom protozoa.
‘Levine Report’ of 1980 [17] acknowledged seven phyla in
The avian Haemosporina comprises three genera,
the Subkingdom Protozoa with 229 taxa above the family
namely Haemoproteus (Haemoproteidae), Leucocytozoon
level. The commonly used guide to the Protozoa appeared
(Leucocytozoidae) and Plasmodium (Plasmodiidae).
with its classification scheme based on the Levine
Plasmodium is a typical malaria parasite. Perhaps most
Report [17]. This scheme classified protozoa into seven
taxonomic papers published before and to a major extent
phyla, namely Sarcomastigophora, Labyrinthomorpha,
now contain morphological descriptions of a taxon.
Apicomplexa, Microspora, Ascetospora, Myxozoa and
New species are often designated without information
Ciliophora. However, Zhang [27] has placed Myxozoa as
on behavior, natural history, key for identification,
a separate phylum of the Kingdom Animalia, although
and phylogenetic hypotheses. Taxonomy changes
Myxozoa was a group of protozoa before. Molecular
rapidly due to molecular biology, sometimes combining
phylogenetic analyses support the close relationship
multiple, both old and novel tools. Currently, individual
between Myxozoa and Cnidaria [22,28] showing its
taxonomists apply modern tools for taxonomic research.
benefits.44 Fatik Baran Mandal, Ph.D.
3.3 Systematics and taxonomy of The vector, Pseudolynchia canariensis Macquart
Haemosporidia (Diptera: Hippoboscidae) was also described as
Microlynchia pusilla, Ornithomyia avicularia, and Lynchia
Haemoproteus, Plasmodium, and Leucocytozoon along hirsuta (Hippoboscidae) [46] (a case of synonymy).
with other genera, form the group Haemosporida. Similarly, misidentification of Haemoproteus columbae
Members of Haemosporidia parasitize amphibians Kruse as H. maccallumi from the avian host Zenaidura
through mammals. There are pitfalls in the identification macroura and as H. palumbis from the host (Columba
of hosts and their haemosporidian parasites. Members of palumbus) present cases of synonymy. In a similar way,
the family Haemoporoteidae Doflein, 1916 are the most Haemoproteus nettionis was described inadvertently as
common parasites of birds. Danilevski [29] first noted H. anatis and H. anseris [47]. Thus, the concept of host
pigmented parasites in the RBC of pigeons. Later in specificity stands no longer valid now. In case of vector
1890, Kruse [30] erected the genus Haemoproteus for the specificity in human malaria, Anopheles is the sole vector.
pigmented parasite, which was similar to Plasmodium. However, mosquitoes belonging to genera like Culex,
Adie [31], O’Roke [32], Coatney and Roudabush [33], Haiba Anopheles, Mansonia, Aedes, Culiseta, and Aedeomyia
[34], White and Bennett [35] studied the taxonomy of transmit species of avian Plasmodium [48]. Again, the rule
avian haemoproteids. The International Reference Centre of vector specificity does not explain such transmission
for avian haematozoa deals with various aspects of avian in birds. Hematozoa was thought to contain Haemosporea
haemoparasitology. In fact, taxonomic errors were due and Piroplasmea before. Levine et al. [17] considered
to ‘one host-one parasite’ belief. Besides, morphological haematozoa to contain Haemoproteus, Trypanosoma,
variations of the same parasite in various hosts also Leucocytozoon, Plasmodium, Babesia, and Lankesterella.
created errors. Molecular studies show the evolutionary The term haematozoa is now less in use following
origin and prevalence of Plasmodium species in birds of classification of Protozoa [17]. The members of the group
various orders across different countries [36-39]. The level Haematozoa lacks sporocysts, conoids, and nine-triplet
of host order in case of Haemoproteus and Leucocytozoon centrioles which are typical of eukaryotes.
is limited. Host based parasite taxonomy was also a source Controversy exists regarding the number of families
of taxonomic confusion, which required re- classification in Haemosporina. Some claim that it should contain the
of avian fauna. family Plasmodiidae only, while others claim three families.
The use of molecular markers for avian In the latter case, the three families are Haemoproteidae,
haemosporidians began in 2000 [40]. Valkiunas [41] Garniidae (genus Fallisia), and Plasmodiidae. According
considers the Haemoproteidae as the most primitive group, to another opinion, the family Leucocytozoidae is distantly
and Leucocytozoidae as the most recent. The excellent related to the above-mentioned groups. It is proposed
review on Valkiuna’s study by Catherine Cosgrove [42] to include Plasmodium, Haemoproteus, Hepatocystis
is mention-worthy here. Taxonomy of the Leucocytozoon and Plasmodium in a monophyletic clade as the malaria
is being revised using molecular techniques to achieve parasites. Leucocytozoon is also considered as a malaria
accuracy. This is supposed to change the parasite- vector- parasite in another approach [49]. This proposal has
bird associations. However, Garnham and Duggan [43] created debate among the protozoologists and researchers
included at least 17 genera and subgenera inhabiting of other related fields.
various reptile, bird and mammal species in the order
Haemosporidia. These genera and subgenera are Akiba,
Fallisia, Garnia, Vinckeia, Nycteria, Polychromophilus, 3.4 Haemosprodia
Rayella, Giovannolaia, Huffia, Haemamoeba, Novyella,
Carinamoeba, Haemocystidium, Ophidiella, Sauramoeba, Members of avian Haemosporidia represent an excellent
Saurocystozoon and Hepatocystis. However, Levine et al. model system for studying vector borne protozoan
[17] in their revised classification of the Protozoa have diseases like malaria. The morphology and life cycles
not used the term Haemosporidia. A total of 145 avian of Plasmodium, Haemoproteus, and Leucocytozoon
haemoproteid species along with 35 spp. as nomen nudum are complex. The family Garniidae comprises the
and one as numen dubium were listed in 1988 while this lesser-known genus Fallisia. Mixed infections with
number was 196 in 1982 [44]. Multi host parasites may Plasmodium and Haemoproteus are common. Similarities
explain this [45] instead of strict host specificity. This and differences in the life histories of Plasmodium,
again is a case of taxonomic hurdle. Hepatocystis, Haemoproteus, and Leucocytozoon can be Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 45
generally summarized. Laveran [50] for the first-time have one and some have two types of gametocytes [43].
observed Plasmodium in 1881 in humans. Danilevski [51] Leucocytozoon marchouxi is pathogenic for doves and
realized its parasitic nature. Since then, many studies pink pigeon [57] while L. danilewskyi for some owl species.
focused on the biology Haemosporidia. Laboratory studies Schizonts of Leucocytozoon occur in muscle, cause damage
of avian malaria centered on the Plasmodium gallinaceum. and necrosis [58]. Extent of tissue damage causes mortality.
This parasite stands as one of the first models for human Leucocytozoon causes a disease called leucocytozoonosis.
malaria. Avian haemosporidians are important for study The disease is also called gnat fever, turkey malaria, and
for their association with birds’ health. Introduction of duck malaria which can be fatal in poultry. Number of
bird malaria into Hawaii in the 19th century affected the described species of Leucocytozoon is over 100 [59,60].
endemic birds. Outbreaks of the haemosporidian infection However, vectors are known only for a few species of
cause serious economic losses in poultry. Five subgenera Leucocytozoon. Simulium anatinum and S. rugglesi transmits
of genus Plasmodium are Giovannolaia, Bennettinia, L. simondi. L. smithis and L. simondi are economically
Novyella, Haemamoeba, and Huffia. Plasmodium causes important. Simulium meridionale and S. slossonae transmit
the disease malaria. Female Anopheles transmits L. smithi that inhabits turkeys. Molecular work has added
Plasmodium. MacCallum [52] discovered the asexual more diversity of Leucocytozoon in its vector, the black
stages of Plasmodium in RBC of birds. Ronald Ross [53] fly [61,62]. Birds serve as a host for asexual development
described the sporogony of Plasmodium relictum in for Leucoctozoon (schizogony and gametogony; reviewed
culicine mosquitoes. Shortt and Garnham [54] discovered by Adler et al. [63]. Birds with chronic infections
that Plasmodium completes schizogony in the liver show weak immune response and low reproduction.
before starting the blood cycle in the year1948. In 1982, Leucocytozoonosis had serious effects on the poultry in
Krotoski and others [55] described the dormant stages North America [63]). The U.S. Agricultural Research Service
of the sporozoites called hypnozoites in the liver [56]. estimated an average loss of nearly $750,000 per year in the
Plasmodium causes avian malaria with sublethal effects US during 1942 to 1951 due to leucocytozoonosis in turkeys.
on host fitness. The most detrimental long-term impact of In the 1970s, the last major outbreaks of leucocytozoonosis
avian malaria is the reduction of the host population due in domestic turkeys happened. Turkeys now are raised
to effects on the host’s reproductive system. primarily in poultry houses, reducing the incidence of
Members of the family Hippoboscidae and disease because the vectors generally do not venture inside
Ceratopogonidae (Diptera: Ceratopogonidae) are shelters [64]. Leucocytozoon enlarges the liver and spleen
the vectors of the subgenera Parahaemoproteus and and causes anemia [63].
Haemoproteus of genus Haemoproteus, respectively. Host specificity of a parasite ranges from specialists
Parahaemoproteus does not infect doves and pigeons infecting one, or a few related host species to generalists
(Columbiformes). Number of species is the highest in the which infects many host species. Host specificity has
genus Haemoproteus among avian Haemosporidia. Some no relationship to the abundance, or evolutionarily
species of Haemoporoteus pose threats to their avian distinctiveness of the host(s). Haemoproteus is the most
hosts. Haemoproteus produces subclinical infections with common genus encountered in bird species, followed
enlargement of kidneys, liver, spleen, and gizzards and also by Plasmodium and Leucocytozoon. Most species of
invades the lungs. Anorexia, anemia, and depression have Haemoproteus and Leucocytozoon are relatively host
been reported in some birds infected with Haemoproteus. specific and found in closely related species within the
The subgenus Haemoproteus infects non-passerine bird same host family. Species of avian Plasmodium have much
species from the Columbiformes, Pelecaniformes, and broader host specificity and may occur in several avian
Charadriiformes. Parahaemoproteus infects birds across families [43,65,66].
all the avian phylogeny [38]. Leucocytozoon and Akiba are
the two subgenera of the family Leucocytozoidae. Only one
species, Akiba, is known for avian Leucocytozoidae. Life 3.5 Life cycles of members of avian
cycle of Leucocytozoon is similar to that of Haemoproteus. Haemosporidia
Gametocytes of various species of Leucocytozoon develop
either in RBC, or lymphocytes. Gametocytes lack Members of Haemosporidia completes their sexual phase
haemozoin granules like Plasmodium and Haemproteus of life cycle in the vector and they use their vertebrate host
but distort the host cell. Morphologic forms of gametocytes for completion of their asexual phase of life cycle. Both
are either round or elongated (fusiform). Some species the phases of life cycles are described below:46 Fatik Baran Mandal, Ph.D.
3.5.1 Sporogony gametes unite to produce zygote in the vector. Zygote
develops through stages such as ookinete and oocyst
The sexual cycle takes place in dipteran vectors and results in the infective stage, which is called the
in all three genera. Hematophagous dipterans sporozoite. In Hepatocystis, the ookinete penetrates the
(Insecta: Diptera) of the families Ceratopogonidae, stomach wall of Culicoides and moves into its hemocoel
Hippoboscidae, Culicidae, and Simuliidae serve as the to develop into oocyst.
vectors. Both mosquitoes and biting midges of the order Two types of oocyst are described in Haemoproteus.
Diptera are the vectors for Plasmodium and Hepatocystis, H. columbae in the hippoboscid flies develop into large
respectively. Hippoboscid flies and biting midges serve oocysts with many sporozoites. Oocysts of H. columbae that
as the vectors of Haemoproteus spp. Simuliids and mature in Culicoides are slightly larger than the zygotes
biting midges transmit Leucocytozoon. Life histories from which they arise. Two types of sporozoites develop
of Plasmodium, Haemoproteus and Leucocytozoon bear differently from two types of oocysts. In Leucocytozoon,
distinctiveness to place them into three families namely the oocysts are relatively small; some of which develop
Leucocytozoidae, Haemoproteidae, and Plasmodiidae intracellularly, some intercellularly, and some below the
respectively. The sporogony (sexual cycle) comprises basement membrane. Sporozoites in Leucocytozoon have
stages like gametes, zygote, ookinete, oocyst, as well one round and one pointed end similar to sporozoites
as the end product, the sporozoites and they occur of Haemoproteus in hippoboscid vectors. Sporozoites of
in vector fly. Life histories of the said three genera Haemoproteus in Culicoides possess pointed ends similar
show a limited specificity for the dipteran vectors. to that of Plasmodium and Hepatocystis.
Mosquitoes (e.g. Anopheles, Culex bitaeniorynchus,
Culex vishnui, Aedes butleri, Mansonia uniformis,
Aedes pseudomediofasciatus) transmit avian malaria. 3.5.2 Schizogony
Hippoboscid flies transmit Haemoproteus columbae
of pigeons. Culicoides also transmit some species of Schizogony is the asexual lifecycle in which sporozoites,
Haemoproteus. the end product of sporogony, transform into schizonts
Several ornithophilic simuliids transmit and upon maturity schizont bursts to liberate merozoites.
Leucocytozoon. A female black fly ingests the gametocytes Two and possibly three types of schizonts are found in
of Leucocytozoon during taking its bloodmeal from the Leucocytozoon. Hepatic schizonts occur in cells of the
infected bird. The parasite then undergoes both asexual liver. Large megalo-schizonts of L. simondi occur in
and sexual development. It completes its life cycle over the liver, kidney, pancreas, brain, heart, gizzard wall,
3–4 days on the fly. During the next blood meal, the fly intestinal wall, and in other locations. Most of the
transmits the infected form of the parasite, the sporozoites merozoites released from the schizont invade blood
to the host. In Leucocytozoon sporozoites are injected into cells to become gametocytes; some also produce another
the host blood by the vectors, the black fly, during their generation of schizont. Thus, a cycle of schizogony
blood meal. Members of the genus Leucocytozoon infect continues to happen for ultimate production of the end
a wider range of host cells. However, when they infect the product (merozoite) of schizogony to continue the life
RBC, Leucocytozoon digests hemoglobin without forming cycle of the parasite. In Plasmodium, schizogony occurs
the hemozoin granules. In the case of Leucocytozoon and in the blood, liver and other tissues. Schizogony starts
Haemoproteus, exflagellation, the process of male gamete after the entry of sporozoites in the host. In case of other
formation, occurs rapidly in comparison to Plasmodium two genera, it occurs only in the tissues. The schizogony
gallinaceum. (asexual cycle) with the end product of merozoites,
Culicoides adersi transmits Hepatocystis kochi in as well as the gametogony, occurs in avian hosts, or
mammals. Hepatocystis kochi constitutes an important vertebrate hosts. Gametogony results in the production
position in phylogenetic hypotheses for the relationship of male and female gametocytes. The exo-erythrocytic
for several genera of Haemosporidia inhabiting cycle takes place in endothelial cells of capillaries,
mammals, lizards and birds [49]. Thus, Plasmodium lymphocytes, spleen, and in liver parenchymal cells. In
and Hepatocystis develop in mosquitoes and simulids, Haemoproteus, a prepatent period lasts for two to three
respectively. The parasite in the form of sporozoite weeks. In Leucocytozoon simondi, it lasts for less than
enters the vector fly during sucking their host blood. six days. The prepatent period is the time gap between
The male and female gametocytes on entering the vector the entry of the infective stage in the host and clinical
fly produce male and female gametes, respectively. The manifestation of the disease. Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 47
3.6 Molecular phylogenetics in resistance in Plasmodium to control malaria infection.
Haemosporidia Molecular phylogeny enables us to control such diseases.
Molecular approach uses the genetic complement,
Microscopy was the method to identify avian DNA –hybridization, DNA bar-coding to name a few.
haemosporidia with morphological data before 2000. Haemosporidia is a highly diverse group with a wide
This was and is the traditional taxonomy in case of host range. Thus, phylogenetic and ecological studies
microscopic protozoa. Several morphological differences propelled the use of molecular tools [70]. mtDNA is the
help to identify various parasite genera in a blood smear/ smallest in the haemosporidian among all eukaryotes.
tissue smear. Vector competency, life history traits, This DNA has cytochrome c oxidase subunit1 (cox1),
and morphological data together formed the earliest cytochrome b(cytb) and cytochrome c oxidase subunit
phylogenies of Haemosporidia before the 1990s. Renewed 3(cox3). mtDNA genes identify species including cryptic
interest in avian haemosporidian emerged from the species to draw phylogenetic relationships. It also explores
development of reliable molecular methods. These tools the biogeographical and historical patterns [62,70,71].
ease the screening of haemosporidian infection. They are Realizing the importance of mtDNA, Pacheco et al. [70]
also important for reexamining the global epidemiology of reviewed the rate of mtDNA evolution. They used a data
infection with haemosporidian. Molecular data has helped set comprising 114mt DNA genomes from Haemoproteus,
to alter and clarify this original phylogeny. Polymerase Plasmodium, Hepatocystis, and Leucocytozoon. Pacheco
chain reaction (PCR) became a prominent molecular and others in the year 2017 estimated the phylogeny
tool to screen the haemosporidians. For Plasmodium and divergence times for the distribution of major
and Haemoproteus, and Leucocytozoon, the PCR screens haemosporidians of birds. The study shows that mtDNA
targeting the cytb gene stand reliable often. Mitochondrial genome is a suitable biomarker for phylogenetic analysis.
cytochrome b (cytb) sequences of H. sacharovi have In the mtDNA phylogeny, Plasmodium of lizards and birds
been identified. Phylogenetic relationships of subgenus appeared as monophyletic. However, the group contains
Haemoproteus show that H. sacharovi and H. turtur branch five clades, four of which are well-supported based on
from the clade along with the Parahaemoproteus. The their biomarker-based similarities. In a similar case,
study provided barcodes for H. sacharovi, and predicted a only six species of Plasmodium were examined for their
revised classification of avian haemoproteid species. One relatedness from lizard hosts. As the number of species
can get a clear picture of the systematics of the concerned examined were few, it was inconclusive that the said
taxa from the study as well. species of Plasmodium could form a monophyletic group.
The current subgeneric classification of avian It is interesting to note that 101 species and subspecies of
haemoproteids is generally effective. However, the Plasmodium inhabit various species of lizards. Pacheco
position of some species requires revision [67]. This et al. [70] also showed that whole mtDNA was vital
again shows that systematics is still enigmatic, at least for phylogenetic analysis. Recently, Nilsson et al. [71]
in some cases. The identification of avian malaria reported multiple cryptic species of Haemoproteus majoris
parasites was revolutionized by many researchers [37,68]. from its avian host. In more recent times, true diversity of
Bensch et al. [37] designed a primer set to amplify a haemosporidians is believed to be over 260 species [72].
478-nucleotide part of the cytb gene from Haemoproteus Variation in the bar-coding region of the cytb gene has
and Plasmodium of birds. Designing more primers since resulted in over 3600 lineages. Besides, phylogenetic
then amplified many nucleotide fragments. Some primers analysis of avian haemosoridian was detailed by many
amplify Leucocytozoon fragments homologous to those of workers [for example 73-79]. The cytb gene is useful to
Haemoproteus and Plasmodium. This makes it possible to identify parasite lineage and to know the evolutionary
compare the same fragment in all three genera [69]. history of avian haemosporidian. Multigene phylogenetic
Traditional taxonomy described 500-odd species of analysis is useful and most reliable in this context [73,74].
avian haemosporidia, of which less than half were valid Phylogenetic evidence emerged from the molecular
[41]. Molecular approach is promising to construct the data has questioned traditional taxonomic schemes. As a
evolutionary history of an organism through molecular result, major changes in the taxonomic practices are also
phylogeny. Parasites existed before the emergence arising [80]. The acquisition of reliable evidence is the key
of mammals or birds. Wild animals are the source of for a stable scheme in taxonomy. Phylogenetic evidence
many human diseases called zoonotic diseases like is crucial to get a picture of phylogeny using modern
human malaria. We must trace out the origin, spread, taxonomy [80]. Multigene phylogenies are appearing
insecticide resistance in vector mosquitoes and drug for many genera, including Plasmodium [81,82]. The48 Fatik Baran Mandal, Ph.D.
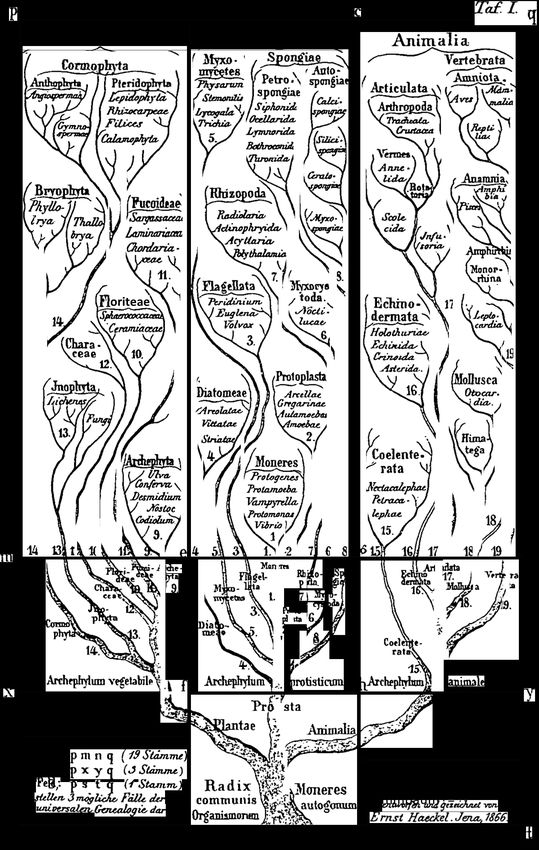
Fig. 1 Reproduction of HAECxcr’s phylo- genetic tree of life (from HxEcKEL 1866. Plate I).
Source: Corliss [5]. Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 49
availability of genome sequence for selected apicomplexan similar cryptic species, although appears to be a single
parasites has revealed the genomic landscape of virulence species, may consists two or more species, which can be
gene families [83]. Detailed information on virulence determined by the molecular method.
genes is useful to control infectious diseases. Interested Phylogenetic parameters like molecular clock and
readers may avail further information from the recent extinction rates for Haemosporidia are still lacking. This is
publication [84]. Phylogenetic studies analyzed the due to the lack of fossil DNA. The rate of mtDNA divergence
sequences from Hepatocystis (mammal), Haemoproteus between species is about 2% per million years. Studies
(bird and lizard), and Plasmodium (mammal, bird, and suggest a slower clock for Haemosporidia than their
lizard). Perkins and Schall [85] analyzed the cytb gene and vertebrate hosts. It is unclear how much slower this clock
formed the tree that rooted with Theileria annulata. Two would be. Generation time is shorter for Haemosporidia
included Leucocytozoon sequences were basal in the tree than vertebrates. Therefore, a slower haemosporidian
as a close sister out-group to the in-group. Martinsen et al. clock is surprising. A divergence rate of 1.3% per million
[49] used Leucocytozoon both as the out-group and to root years for Haemosporidia is lower than that of their avian
the tree. Outlaw and Ricklefs [82] analyzed the same data hosts. This rate is higher than co-speciation estimates
using an alternate rooting method. (Ricklefs and Outlaw [89]. If 1.3% divergence rate is
Comprehensive phylogenies show that correct, certain haemosporidians could have diversified
Parahaemoproteus and Haemoproteus are distinct. to infect many hosts in the last 20 million years [89].
They were intermediate between Plasmodium and Hellgren et al. [90] analyzed fragments of the mtcytb
Leucocytozoon. Parahaemoproteus is more related to gene from six morphospecies of avian Haemoproteus.
Plasmodium in birds. Plasmodium is not monophyletic due The average divergence between the morphospecies
to Hepatocystis. In all three comprehensive phylogenies, was 5.5%. This suggests that separate morphospecies
Plasmodium formed two clades based on vertebrate hosts resulted in sequences with a genetic difference >5%. Two
(one in mammal and the other in lizard/bird). Evidence has morphospecies differed by only 0.7%. Krizanauskiene
been mounting for a revision of Haemospordia taxonomy. et al. [91] found that a divergence of 2% was enough for
Controversy arose to redesignate haemosporidians of species to show external distinctness. Closely related P.
birds and reptiles. Most malaria trees contain an out- falciparum and P. reichenowi diverge by 2.3% across their
group from the genus Leucocytozoon, which is related mtDNA. Insight into haemosporidian phylogeny can trace
to, but not the malaria genera. The reason is that the the origin of Plasmodium in primates. Characterizing
gametocytes of Leucocytozoon lack hemozoin. Meronts patterns of host–parasite associations in wild populations
of the Leucocytozoon are not present in the blood of the would produce models. These models can predict the
vertebrate host. For this, along with a high divergence rate, probability of infection with haemosporidian to hosts.
Leucocytozoon was believed to be ancestral to Hepatocystis,
Haemoproteus, and Plasmodium. Leucocytozoon is the
ancestral genus and therefore its role as an out-group is 3.7 Phylogenetic Relationships among
controversial. Leucocytozoon may be the most derived members of Haemosporidia
genus and is a sister group to Haemoproteus. Plasmodium
is ancestral to both (species infecting mammals being Diversity of Haemosporidian among raptorial birds was
most ancestral). This is the opposite tree configuration estimated by DNA sequencing. mtcytb revealed raptor-
from what was before found. Outlaw and Ricklefs [82] specific parasite clades in Parahaemoproteus, although
viewed merogony in blood and hemozoin production as not in Plasmodium. A divergent clade of raptor parasites
primitive. These traits evolved once in Haemosporidia was found as a sister clade to Leucocytozoon [92]. The
and were then lost in some genera. Plasmodium and evolutionary relationships among malaria parasites
Haemoproteus both keep the hemozoin trait. Only the is still an annoying phylogenetic problem. This is due
Plasmodium keeps the merogony trait. Leucocytozoon, as to bar in sampling of taxon, character and nucleotide
a most derived genus, has lost both traits. Plasmodium base composition biases characteristics of this clade.
keeps the merogony trait, which was the basis for creating The phylogenetic analysis of Plasmodium from a diverse
this genus, as merogony in blood was perhaps a recent vertebrate host were studied. Results showed that
acquisition to Haemosporidia. Interestingly, levels of evolutionary history of Plasmodium through a complex
species richness and morphologically cryptic species have series of transitions. Plasmodium is polyphyletic with their
been under-estimated for tropical biodiversity [86-88] own life-history traits. The traits evolved in a dynamic
due to rare use of DNA-based methods. Morphologically manner across the phylogeny. DNA sequencing estimated50 Fatik Baran Mandal, Ph.D.
the diversity of haemosporidian among raptorial birds. of species is likely to be higher than can be recognized
Further studies in future covering various avian families by microscopy. Thus, both the parasites and their
would contribute on the accurate diversity of bird hosts require study in terms of evolution. The diversity
inhabiting Haemosporidia. of members of Apicomplexa with recent advances in
To reconstruct the phylogenetic relationships among molecular phylogeny shows the drawbacks of the morpho-
the Plasmodium researcher used SSUrRNA gene sequences taxonomy, and stresses the absence of lineages and taxa
from 13 Plasmodium species inhabiting mammals, birds, in recent phylogenies [97].
and lizards. The phylogenetic tree thus constructed The importance of life history and epidemiological
comprises two major clades. The first with bird and reptile characteristics of the genera Haemoproteus and Plasmodium
parasites while the second with the rest of the species. are crucial to know their evolutionary relationships
Bird parasites, P. gallinaceum and P. lophurae, do not with their vertebrate hosts and vectors [98]. Martinsen
cluster with human, chimpanzee, or rodent parasites, but et al. [49] proposed to include the monophyletic clade
cluster with lizard parasites [93]. comprising Hepatocystis, Plasmodium, Haemoproteus,
Outlaw and Ricklefs [94] proposed a phylogeny where Parahaemoproteus under the malaria parasites. Braga
Plasmodium and Hepatocystis of mammals were sister to et al. [99] thought only Plasmodium as the avian malaria
all haemosporidians from reptiles and birds as proposed parasite. Identification of microscopic animals is carried
in another phylogeny. Avian Plasmodium found at the base out following some morphological keys in morphological
of avian Haemosporidia, followed by Parahaemoproteus. approach. As there are various developing stages of
Haemoproteus was sister to the genus Leucocytozoon. the parasite and morphology of a particular stage may
This example shows that to construct phylogeny in change depending on various conditions, it sometimes
separate groups or taxa is difficult without considering leads to incorrect identification. This does not happen
broad evolutionary processes. These processes are host with a molecular approach. Thus, this approach helps to
switching, ecology of host, transmission opportunity eliminate controversy in the taxonomic position of the
of the parasite from one host to the other. The first organism.
study comparing molecular phylogenies with morpho-
taxonomy got support from genetic and phylogenetic
species concepts as well. For constructing a phylogenetic
4 Discussions and conclusion
tree different criterion like molecular clock, nonreversible
Three species concept defines a species based on
model of DNA substitution and outgroup are in use.
morphological, genetic, or phylogenetic characters.
Huelsenbeck et al. [95] found outgroup and molecular
The morphological species concept distinguishes
clock as the best criteria for detecting the root of the tree.
parasite species through similarities or dissimilarities
The nonreversible model works in presence of highly
in morphology using microscopy. The genetic species
non reversible substitution process. Leucocytozoon
concept is a molecular method that identifies parasites
was found at the base of haemosporidians along with
based on similarities or divergence of DNA, RNA, or amino
avian Plasmodium and Haemoproteus as sister taxa
acid sequences. The phylogenetic species concept requires
(monophyletic) in their phylogeny [49].
that defined species are monophyletic. Martinsen et al. [49]
Use of the ssrRNA gene in phylogenetic analysis reveals
identified fourteen of fifteen Haemoproteus, Plasmodium,
that avian Plasmodium subgenera do not correspond
and Leucocytozoon parasite species using morphology.
to their morpho-taxonomy. A two-gene mitochondrial
Genetic and phylogenetic analyses supported such
phylogeny (cytb and col) of avian Plasmodium got support
identification. The exception being H. belopolskyi, which
for three subgenera namely Huffia, Haemamoeba and
belong to two separate phylogenetic clades. Three species
Bennettinia, out of the five morphology-based subgenera
concept validates five subgenera of Plasmodium that infect
[96]. Perhaps the reptiles hosted several ‘ancient’ genera
birds at molecular level. Analysis of the coI and mtcytb
of Haemosporidia. Haemosporina of modern reptiles have
genes validated the monophyly for Haemamoeba, Huffia,
evolved from such an organism. As birds evolved from
and Bennettinia. While most Novyella sequences formed a
the saurischian dinosaurs, parasitologists do not rule
clade, the two Novyella samples were outside the clade,
out a parallel evolutionary process with the extant avian
forming the most basal branches of the tree. The subgenus
Fallisia, Haemoproteus, Leucocytozoon, and Plasmodium.
Giovannolaia did not form a monophyletic group.
Mammalian haemosporidians perhaps evolved from an
The present article deals with the conflict between
earlier reptilian stock than birds. High genetic diversity
morphotaxonomy and molecular taxonomy to draw the
in members of Haemosporidia shows that the number Hurdle in taxonomy: A case of malaria parasites and other Haemosporidia 51
attention of the scientists to end the conflict for furtherance Conflict of Interest
of the protozoology and related subjects with an example Author states no conflict of interest.
of the avian Haemosporidia. From the evolutionary
Data Availability Statement
relationship, mammalian Plasmodium species is the sister
Data sharing is not applicable to this article as no datasets
clade to reptilian and avian malaria parasites. It has been
were generated or analyzed during the current study.
concluded that “Hepatocystis appears first sharing a
common ancestor closer to primate malaria parasites than
the avian parasites. Haemoproteus columbae appears References
to be closely related to two avian malaria parasites.
Study of systematics of Plasmodium requires the study [1] Corliss JO. Progress in protistology during the first decade
following reemergence of the field as a respectable
of Hepatocystis, Haemoproteus and Leucocytozoon and
interdisciplinary area in modern biological research. Prog
lizard malaria parasites’ [56]. Speciation events due to host
Protistol. 1986;1:11–63.
switching are very common among protozoan parasites. [2] Cavalier-Smith T. Kingdom protozoa and its 18 phyla. Microbiol
Ecological study is very important to get a holistic view of Rev. 1993 Dec;57(4):953–94.
systematics. Morphotaxonomy may not be always suitable [3] Cavalier-Smith T. A revised six-kingdom system of life. Biol Rev
for ascertaining species identity, or to draw its evolutionary Camb Philos Soc. 1998 Aug;73(3):203–66.
[4] Corliss JO. An interim utilitarian (“ user-friendly”) hierarchical
relationship with other species. Molecular phylogenetics
classification and characterization of the protists. Acta
consider the information collected through application of Protozool. 1994 Jan;33:1–51.
other approaches to taxonomy to give a clear picture of the [5] Corliss JO. Classification of protozoa and protists: the current
taxa under consideration [100-2]. I also find some studies status. In: Coombs GH, Vickerman K, Sleigh MA, Warren
[103-8] as interesting to get a further insight into the topic A, editors. Evolutionary relationships among protozoa.
Dordrecht: Springer Netherlands; 1998. pp. 409–47.
under discussion. Study on consensus mitochondrial
[6] Corliss JO. Protozoan taxonomy and systematics. eLS
nuclear, and apicoplast gene sequences showed a clear
[Internet]. 2001 May [cited 2021 Jul 14]:[about 7p.]. https://doi.
relationship among haemosporidian genera both with org/10.1038/npg.els.0001925.
their hosts and their vectors. Groupings is obvious among [7] Sleigh MA. Protozoa and other protists. Cambridge:
the Leucocytozoon, Haemoproteus, Parahaemoproteus Cambridge University Press; 1991.
and Plasmodium inhabiting birds, reptiles, rodents and [8] Hausmann K, Hülsmann N. Protozoology. Stuttgart: Georg
Thieme Verlag; 1996.
primates, and Hepatocystis from bats. These parasites
[9] Lücking R. Three challenges to contemporaneous taxonomy
use black-flies, louse-flies, midges, and mosquitoes as from a licheno-mycological perspective. Megataxa. 2020
vectors to complete their life cycle. Studies on host and Jan;1(1):78–103.
parasite phylogenetic trees also show varying degrees [10] Thompson FC. Names: the keys to biodiversity. In:
of co-evolution [109]. Evolutionary relationships among Reaka-Kudla ML, Wilson DE, Wilson EO, editors. Biodiversity
II. Understanding and Protecting Our Biological Resources.
species must be gathered to provide further insight in
Washington (DC): Joseph Henry Press; 1997. pp. 199–211.
systematics and biodiversity. The last but not least, I
[11] Mandal FB. Human parasitology. New Delhi: PHI Learning
would like to conclude that in the case of systematics of Private Limited; 2015.
parasitic protozoa, both the evolution and ecology of host [12] Honigberg BM, Balamuth W, Bovee EC, Corliss JO, Gojdics M,
and parasite requires careful consideration to provide a Hall RP, et al. A revised classification of the phylum Protozoa. J
holistic view of the organisms or group of organisms. Protozool. 1964 Feb;11(1):7–20.
[13] Bovee EC, Lee JJ, Hunter SH. (Lee JJ, Hutner SH, Bovee EC,
Besides, by using the information so far revealed using
editors). Class Granuloreticulosea De Saedeleer, 1934. An
modern tools and techniques in this context would help Illustrated Guide to the Protozoa. Lawrence (KN): Society of
to simplify the taxonomy and systematics. Protozoologists; 1985. pp. 246–52.
Acknowledgement: The author wishes to thank Dr. [14] Barnes RD. Invertebrates zoology. 6th ed. Philadelphia: W. B.
Asit K. Biswas. Associate Professor in English, Bankura Saunders Company; 1982.
[15] Parker TJ, Haswell WA. Textbook of zoology. London: Macmillan
Christian College, Bankura for his help to revise the
International Higher Education; 1967.
manuscript. He also wishes to extend his sincere thanks
[16] Pechenik JA. Biology of the Invertebrates. New York:
to the anonymous reviewers for their valuable suggestions McGraw-Hill Science; 2010.
in improving the quality and content of the manuscript. [17] Levine ND, Corliss JO, Cox FE, Deroux G, Grain J, Honigberg
BM, et al. A newly revised classification of the protozoa. J
Funding information Protozool. 1980 Feb;27(1):37–58.
Author states no funding involved.52 Fatik Baran Mandal, Ph.D.
[18] Agnarsson I, Kuntner M. Taxonomy in a changing world: mitochondrial DNA amplified from birds. Proc Biol Sci. 2000
seeking solutions for a science in crisis. Syst Biol. 2007 Aug;267(1452):1583–9.
Jun;56(3):531–9. [38] Beadell JS, Gering E, Austin J, Dumbacher JP, Peirce MA, Pratt
[19] de Queiroz K. A unified concept of species and its TK, et al. Prevalence and differential host-specificity of two
consequences for the future of taxonomy. Proc Calif Acad Sci. avian blood parasite genera in the Australo-Papuan region.
2005;56 Suppl 1:196–215. Mol Ecol. 2004 Dec;13(12):3829–44.
[20] de Pinna MC. Species concepts and phylogenetics. Rev Fish [39] Bensch S, Hellgren O, Pérez-Tris J. MalAvi: a public database
Biol Fish. 1999 Dec;9(4):353–73. of malaria parasites and related haemosporidians in avian
[21] Imam TS. The complexities in the classification of protozoa: hosts based on mitochondrial cytochrome b lineages. Mol Ecol
a challenge to parasitologists. Bayero J Pure Appl Sci. Resour. 2009 Sep;9(5):1353–8.
2009;2(2):159–64. [40] Waldenström J, Bensch S, Kiboi S, Hasselquist D, Ottosson
[22] Mandal FB. Biology of Non-Chordates. New Delhi: PHI Learning U. Cross-species infection of blood parasites between
Private Limited; 2018. p. 488. resident and migratory songbirds in Africa. Mol Ecol. 2002
[23] Hogg J. On the distinctions of a plant and an animal, and on a Aug;11(8):1545–54.
fourth kingdom of nature. Edinb New Philos J. 1860;12:216–25. [41] Valkiunas G. Avian malaria parasites and other haemosporidia.
[24] Haeckel E. Generelle Morphologie der Organismen [General 1st ed. Boca Raton, London, New York (NY), Washington (DC):
morphology of organisms]Berlin, New York: De Gruyter; CRC press; 2004. https://doi.org/10.1201/9780203643792.
2011. https://doi.org/10.1515/9783110848281. [42] Cosgrove C. Avian Malaria Parasites and Other
[25] Corliss JO. Haeckel’s kingdom Protista and current Haemosporidia.—gediminas Valkiunas. 2004. CRC Press, Boca
concepts in systematic protistology. In: Welträtsel und Raton, Florida, USA. 932 pp. ISBN 0-415-30097-5. US$170
Lebenswunder. Ernst Haeckel - Werk, Wirkung und Folgen. (hardcover). Syst Biol. 2005;54(5):860–3.
Linz: OÖ Landes-Kultur GmbH;1998. p. 85-104 (Kataloge des [43] Garnham PC, Duggan AJ. Catalogue of the Garnham collection
Oberösterreichischen Landesmuseums Neue Folge; jahrg. 131). of malaria parasites and other haemosporidia. Cambridge:
[26] Woese CR, Fox GE. The concept of cellular evolution. J Mol Evol. Cambridge University Press; 1988.
1977 Sep;10(1):1–6. [44] Mandal FB. Present knowledge about Haemoproteus columbae
[27] Zhang ZQ. Animal biodiversity: An outline of higher-level (Apicomplexa: Haemoproteidae) with a note on evolutionary
classification and survey of taxonomic richness. Auckland: relationship among avian malaria parasites. Wesl J Res.
Magnolia press; 2011. 2008;1:85–90.
[28] Kent ML, Andree KB, Bartholomew JL, El-Matbouli M, Desser [45] Pedersen AB, Altizer S, Poss M, Cunningham AA, Nunn CL.
SS, Devlin RH, et al. Recent advances in our knowledge of the Patterns of host specificity and transmission among parasites
Myxozoa. J Eukaryot Microbiol. 2001 Jul-Aug;48(4):395–413. of wild primates. Int J Parasitol. 2005 May;35(6):647–57.
[29] Danilevski VI. La parasitologie comparée du sang: Nouvelles [46] Mandal FB, Chandra G. The vectors of avian haemoproteids: a
recherches sur les parasites du sang des oiseaux [Comperative review. Trans. Zool. Soc East India. 2006;10(1&2):35–40.
parasitology of blood: New research on parasites of avian [47] Fallis AM, Wood DM. Biting midges (Diptera: Ceratopogonidae)
blood]South Yarra: Leopold Publishing; 2017. as intermediate hosts for Haemoproteus of ducks. Can J Zool.
[30] Kruse W. XVIII. Ueber Blutparasiten. (Aus dem 1957 Jun;35(3):425–35.
bakteriologischen Laboratorium der zoologischen Station [48] Valkiūnas G, Anwar AM, Atkinson CT, Greiner EC, Paperna
zu Neapel.) [About blood parasites (From the Bacterological I, Peirce MA. What distinguishes malaria parasites from
Laboratory of the Zoological Station in Naples)]Volume 121. other pigmented haemosporidians? Trends Parasitol. 2005
Berlin, Boston: De Gruyter; 2021. pp. 359–72. Aug;21(8):357–8.
[31] Adie H. Nouvelles recherches sur la sporogonie de [49] Martinsen ES, Perkins SL, Schall JJ. A three-genome phylogeny
Haemoproteus columbae [New research on the sporogony of malaria parasites (Plasmodium and closely related
of Haemoproteus columbae]. Arch Inst Pasteur Alger. genera): evolution of life-history traits and host switches. Mol
1925;3:9–15. Phylogenet Evol. 2008 Apr;47(1):261–73.
[32] O’Roke EC. The morphology of Haemoproteus lophortyx sp. [50] Laveran A. Un nouveau parasite trouvé dans le sang des
nov. Science. 1929 Nov;70(1818):432. malades atteints de fièvre palustre: origine parasitaire des
[33] Coatney GR, Roudabush RL. Some blood parasites from accidents de l’impaludisme [A new parasite found in the blood
Nebraska birds. Am Midl Nat. 1937 Nov;18(6):1005–30. of patients with Malaria fever: parasitic origin of malaria
[34] Haiba MH. Plasmodia of common Egyptian birds. J Comp accidents]Paris: Libraire J.-B. Balliere et Fils; 1881.
Pathol Ther. 1948 Apr;58(2):81–93. [51] Danilevski VY. About blood parasites (Haematozoa). Rus Med.
[35] White EM, Bennett GF. Avian Haemoproteidae. 12. The 1884;46:948.
haemoproteids of the grouse family Tetraonidae. Can J Zool. [52] Maccallum WG. On the flagellated form of the malarial
1979 Jul;57(7):1465–72. parasite. Lancet. 1897 Nov;150(3872):1240–1.
[36] Escalante AA, Ayala FJ. Evolutionary origin of Plasmodium and [53] Ross R. Report on the Cultivation of Proteosoma, Labbé, in
other Apicomplexa based on rRNA genes. Proc Natl Acad Sci Grey Mosquitos. Ind Med Gaz. 1898 Dec;33(12):448–51.
USA. 1995 Jun;92(13):5793–7. [54] Shortt HE, Garnham PC. Demonstration of a persisting
[37] Bensch S, Stjernman M, Hasselquist D, Ostman O, Hansson exo-erythrocytic cycle in Plasmodium cynomolgi and
B, Westerdahl H, et al. Host specificity in avian blood its bearing on the production of relapses. BMJ. 1948
parasites: a study of Plasmodium and Haemoproteus Jun;1(4564):1225–8.You can also read

















































