High Frequency of Apodemus Mice Boosts Inverse Activity Pattern of Bank Voles, Clethrionomys glareolus, through Non-Aggressive Intraguild Competition
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
animals
Article
High Frequency of Apodemus Mice Boosts Inverse Activity
Pattern of Bank Voles, Clethrionomys glareolus, through
Non-Aggressive Intraguild Competition
Remo Probst * and Renate Probst
Ornis—Biology Engineering Office and Research Institute, Dr. G. H. Neckheimstr. 18/3,
A-9560 Feldkirchen, Austria
* Correspondence: remo.probst@gmx.at
Simple Summary: Ecologically similar animals can only co-occur if resources are used in a sufficiently
diverse manner. The necessary separation can be made according to habitat, food choice and activity
pattern. We investigated this question in an inner-alpine mixed forest for bank voles, Clethrionomys
glareolus, and mice of the genus Apodemus, the latter being larger and stronger. In the presence of
many Apodemus mice, bank voles were indeed more diurnal. We found no alternative explanation for
this temporal partitioning, despite considering possible other influencing factors, such as exposure to
lunar illumination, temperature, precipitation, the presence of predators or the availability of cover.
Our video footage showed that this temporal separation hardly requires any physical interaction
between the small forest rodents. We attribute this to the long common evolution of these animals.
Abstract: Sympatric animals with similar requirements can separate their ecological niches along
the microhabitat, food and time axes. There may be alternative reasons for an interspecific different
activity pattern, such as intraspecific social constraints, predator avoidance or physical conditions
such as temperature, precipitation and illumination. We investigated the importance of intraguild
competition in a 2-year study in an inner-alpine mixed forest, using small forest rodents as our model
species. Apodemus mice were the physically superior, and bank voles, Clethrionomys glareolus, the
inferior competitor. We predicted that bank voles would exhibit increased diurnal activity when
frequencies of the almost exclusively nocturnal Apodemus mice were high during a seed mast year. To
Citation: Probst, R.; Probst, R. High investigate this, we recorded 19,138 1 min videos. Controlling for confounding variables, bank vole
Frequency of Apodemus Mice Boosts diurnal activity was significantly related to the frequency of Apodemus mice. We assume that at high
Inverse Activity Pattern of Bank
densities of Apodemus mice, a purely nocturnal separation in the niche dimensions of time, habitat
Voles, Clethrionomys glareolus,
and microhabitat is no longer sufficient, and therefore an inverse activity pattern by the bank voles is
through Non-Aggressive Intraguild
reinforced. Our videos showed, however, that this does not require persistent aggressive meetings
Competition. Animals 2023, 13, 981.
and we explain this by the long co-evolution of the taxa under study.
https://doi.org/10.3390/
ani13060981
Keywords: Apodemus spp.; Clethrionomys glareolus; activity pattern; camera traps; intraguild
Received: 4 February 2023 competition; interspecific aggression; temporal partitioning; niche separation; mast year; Alps
Revised: 26 February 2023
Accepted: 7 March 2023
Published: 8 March 2023
1. Introduction
Animals are exposed to a plethora of environmental parameters in their habitat. For
Copyright: © 2023 by the authors. small mammals, such as bank voles (Clethrionomys glareolus, formerly Myodes glareolus [1])
Licensee MDPI, Basel, Switzerland. and Apodemus spp. mice (further referred to as Apodemus mice), these are primarily physical
This article is an open access article conditions such as temperature, precipitation and illumination [2–5], as well as biotic factors
distributed under the terms and such as food, predation, intraspecific organisation and interspecific competition [6–12].
conditions of the Creative Commons
Competition among species is a widespread phenomenon [13,14], and long-term
Attribution (CC BY) license (https://
coexistence is only possible if at least one important niche dimension differs. Dimensions
creativecommons.org/licenses/by/
of resource partitioning are mainly habitat, food and temporal niche [15,16]. For bank voles
4.0/).
Animals 2023, 13, 981. https://doi.org/10.3390/ani13060981 https://www.mdpi.com/journal/animalsAnimals 2023, 13, 981 2 of 13
and Apodemus mice, there are indications for competitive mitigation on all three niche axes,
microhabitat [9,17–19], nutrition [7] and chronoecology [4,20,21]. The stronger and larger
Apodemus mice are considered superior to bank voles [20,22].
Activity time has long been known as a niche axis [16,23], however, the relevance
of the temporal niche concept has been much debated [15,24]. In several studies, only
weak interspecific competition or the resulting time separation between Apodemus mice and
bank voles was found [25–27], although Clethrionomys glareolus, in contrast to the nocturnal
Apodemus mice, are frequently diurnal [28,29]. Main problems addressed are the difficulty of
directly observing competition and the need to take into account a wealth of confounding
factors potentially influencing activity time [13,25]. To overcome these methodological
problems, we conducted a 2-year camera-trapping study in the southern part of the Alps,
capturing the forest-dwelling Apodemus mice and bank voles with the available video function.
Camera-trapping is a powerful tool for wildlife research [30–34], including behavioural
investigations of difficult-to-observe species [35,36]. Thus, the diurnal activity of bank
voles could be tested against predictor variables, such as the activity of Apodemus mice and
predators, habitat characteristics and abiotic parameters. Further, the type of interactions
between the two small forest rodent taxa, ranging from tolerant meetings to aggressive
displacements [20], could be studied.
An important prerequisite for comparing the activity patterns of bank voles and Apode-
mus mice is the variation in frequency. Small forest rodents show the greatest population
changes in the context of masting, especially among temperate tree species. Masting is
the pulsed, synchronous production of large seed crops, which can dramatically affect
the consumer community [37–39]. Populations of Apodemus mice show a pronounced
outbreak–crash pattern in response to the seed masts [10]. The findings are less clear for
the more generalist bank vole [7,40–42], where cyclic, stable and intermediate population
dynamics have been found across Europe [40]. The first year of our study was characterised
by an extreme number of pollen not yet observed in the 30-year recording history of the
Austrian federal state of Carinthia [43]. This was especially true for locally dominating
woody plants, such as the Norway spruce, Picea abies, European beech, Fagus sylvatica, and
European hazelnut, Corylus avellana. The positive connection between airborne pollen and
quantity, as well as quality of seed crop in these so-called masting pollen producers could
be proven elsewhere [44–47]. The second year of the investigation was characterized by a
nil crop. In accordance with these particular differences in the availability of plant seeds,
we will hereafter refer to the first year as the mast year and the second as the non-mast year.
In most of central European forest ecosystems, bank voles and Apodemus mice are of
outstanding importance and may be designated as keystone species. They are an essential
food for a number of predators [8,48,49], can themselves act as seed consumers, as well as
dispersers in context-dependent plant-granivore interactions [50–52], may damage forest
trees [53,54], and can be vectors of a number of diseases [55,56].
In our study we expected a sharp increase in small rodent numbers after the mast pulse
and a subsequent collapse of populations, which would particularly affect the specialised
seed-eating Apodemus mice. The variations in frequency would be a prerequisite for
behavioural changes (Assumption 1). As a second important prerequisite for the shift of
the bank voles into diurnal activity due to intraguild competition, we expected an almost
exclusive nocturnal activity of Apodemus mice (Assumption 2). Controlling for confounding
abiotic and biotic parameters, temporal partitioning would be an indirect measure for
reducing intraguild competition. We predicted asymmetric relationships, whereby bank
voles would show an inverse activity pattern with an increased presence of Apodemus mice
(Prediction 1). As a direct measure, we predicted an altered antagonistic behaviour, with
higher numbers of Apodemus mice leading to increased interspecific aggressions against
bank voles (Prediction 2).Animals 2023, 13, 981 3 of 13
2. Materials and Methods
2.1. Species Identification
While bank voles are fairly easy to identify visually on appropriate videos, for example
by the ratio of body to tail length or the size of the ears [29], we have not attempted to
distinguish members of the genus Apodemus. In the latter, identification problems can
occur without calibration to the morphology of the local population at hand [57], and even
genetic identification has proven difficult [58–60]. As Apodemus mice were typically silent,
identification by means of the calls was also ruled out [61]. Due to the well-known habitat
requirements [28,62], we expected the occurrence of the yellow-necked mouse, Apodemus
flavicollis, and of the wood mouse, Apodemus sylvaticus, as most probable in our forested
study area, in an unknown numerical ratio. The alpine mouse, Apodemus alpicola, is absent
from the region according to current knowledge [63].
2.2. Study Area
The camera traps were set up on the mountain range of the Ossiacher Tauern (46,692◦ N;
14,067◦ E, 550 m a. s. l.) in Carinthia, Southern Austria. In the two years of the survey,
the mean annual temperature was 8.88 ◦ C and 9.39 ◦ C, respectively, and the total annual
precipitation was 1159 mm and 749 mm, respectively (meteorological station of Feldkirchen:
https://data.hub.zamg.ac.at/ (accessed on 15 October 2022)). The study area was on the
steep lower slope with a distinctly undulating relief and several small rocks. The concave
areas have moist and humus-rich soils (sandy loams over clay of the pseudogley type),
where a lime-maple noble deciduous forest (Tilio-Acerion) would be the primary vege-
tation. In the convex parts, beech forest types would predominate plant-sociologically
(Melampyro-Fagetum, Galio odorati-Fagetum [64]). European beech, Fagus sylvatica, as the
actual primary tree species has been pushed back, and today Norway spruce, Picea abies,
dominates the forest. However, European beech, limes, Tilia platyphyllos and T. cordata,
sycamore, Acer pseudoplatanus, European ash, Fraxinus excelsior, as well as silver fir, Abies
alba, are mixed in, and therefore the study site is classified as a mixed deciduous and
coniferous forest biotope type. Trees with a low trunk diameter dominate, but there are
individual “achievers”, as well as sections of pole wood and pioneer stages, the latter espe-
cially consisting of the European hazelnut, Corylus avellana. The study site is not located in
any state-protected area.
2.3. Camera Trap Study
2.3.1. Study Period
Camera traps were set up from September 2020 to September 2022.
2.3.2. Type and Model
Wild-Vision Full HD 5.0 with Black-LED flash was utilized for the study. The trigger
speed was lower than 1 s. The passive infrared sensor (PIR) was designed for high sensitivity.
2.3.3. Modifications
To reduce the over-illumination that occurs when animals are close to the camera trap,
half of the LED field was switched off. We changed to 64 MB cards in order to store the
large amount of video data.
2.3.4. Recordings
We used 1 min videos in HD resolution of 1280 × 720 pixels. Only those recordings
where there was no equipment failure, user error, or false trigger were used for further anal-
ysis.
2.3.5. Definition of an Event
All our specifications refer to the recorded 1 min videos. Several individuals of one
species are only counted if they were visible on a single video and at the same time.Animals 2023, 13, 981 4 of 13
2.3.6. Bait
Apodemus mice and bank voles were attracted with unpeeled, black-and-white sun-
flower seeds. Each deployment equated to 0.5 kg per camera trap. The weight of the
sunflower seeds presented was based on experimental results within the genus Apodemus.
At high densities, several hundred grams can be consumed and transported from a single
feeding site, especially during the intensively used first night [65]. In order to address the
questions of relative frequency and activity pattern, it had to be possible to feed a maximum
number of target species individuals for a sufficient period of time. The sunflower seeds
were provided on the ground and not covered. This enabled both a quick triggering of the
camera traps and the most reliable identification of small mammals, as well as other species.
2.3.7. Layout
According to a minimum convex polygon analysis of camera trap sites by means of
QGIS 3.18.0, the study area had a size of 4.8 ha. A random allocation was carried out [66],
with 83 sites being sampled over the course of the investigation.
2.3.8. Spatiotemporal Independence
In order to get a high degree of independence between trials, we set up camera traps
every 12.07 ± 6.32 sd days [67], and sample sites from the last trial were not allowed
to be repeated. Further, the analyses were standardised according to the first 24 h of
video recording per sampling site [68], hereinafter referred to as a camera trap-night.
To minimise the movement between two camera traps and the immigration of small
mammals within one trial, a distance of 58.94 ± 42.47 sd m was maintained between
typically 6 simultaneously active cameras. Distances were determined with QGIS 3.18.0
and corrected for the slope.
2.3.9. Camera Trap Placement
The bait was centred in the middle of the shot and sunflower seeds were spread over
a maximum area of 50 × 50 cm. This distance from the focal point to the camera (1–2 m)
and the height of the camera (typically from 0.25–0.75 m), and thus the angle, depended on
the different slopes in our study area. Camera traps were only placed in locations where
minimal vegetation change was required for a clear line of sight to the bait.
2.4. Data Treatment
For the analysis of day–night activity pattern, we achieved a high degree of indepen-
dence of the data by keeping only the single recording with the highest number of Apodemus
mice or bank voles from all videos of the same camera trap within a 30 min period [69]. The
48 periods of 30 min periods within a calendar day were assigned to either day or night,
separated by sunrise or sunset. The two twilight periods of a day were assigned to the time
of day in which more of the 30 min were located.
2.5. Predictors
For the multivariate regression analysis of the effects of extrinsic factors, we tested
6 additional independent variables besides mast year/non-mast year: (1) lunar illumina-
tion: % illumination in the camera trap-night (https://moonphases.co.uk/ (accessed on
10 December 2022)); (2) minimum temperature: average of the minimum values of the
two calendar days of one camera trap-night (https://data.hub.zamg.ac.at/ (accessed on
12 December 2022)); (3) precipitation at night: precipitation total (mm) from 19:01–7:00 CET
per camera trap-night (https://data.hub.zamg.ac.at/ (accessed on 12 December 2022));
(4) predators: number of 30 min periods per camera trap-night with at least one visible
predator on the available videos. In our dataset, red fox, Vulpes vulpes, stone marten, Martes
foina, European polecat, Mustela putorius, and tawny owl, Strix aluco, were classified as
mandatory predators of small mammals. Moreover, (5) cover: for our year-round compari-
son, we recorded the % of cover with lying deadwood, snags and rocks in a 10 m radiusAnimals 2023, 13, 981 5 of 13
of the camera trap, because numerous studies have shown a positive correlation between
the abundance of voles of the genus Clethrionomys and the amount of woody debris and
brushwood coverage (so-called thigmotaxis [70–72]); and (6) Apodemus spp.: Σ of Apodemus
mice from the pseudo-replication-limited 30 min periods per camera trap-night.
2.6. Statistical Analysis
The data were statistically analysed by the program package R 4.2.2 (https://www.
R-project.org/ (accessed on 14 December 2022)). Mann–Withney U tests were used to
examine the frequency differences between two groups, and χ2 tests to examine the rela-
tionship between nominal variables. The dichotomous day–night activity of bank voles
was investigated by multivariate logistic regression. Activity patterns were analysed with
Hermans–Rasson tests, using the R-package “CircMLE” [73,74]. Overlap coefficient (∆)
between the temporal activity patterns of Apodemus mice and bank voles was estimated
with the R-package “overlap” [75]. This coefficient ranges between 0 (no overlap) and
1 (complete overlap, [76]). Since the number of cases for the comparisons was greater than
75, the ∆4 estimator was calculated [76]. In addition, 95% confidence intervals for ∆4 using
10,000 bootstraps were determined. The overlap was rated as “medium” when ∆4 was
between 0.50 and 0.75, as “high” with ∆4 > 0.75 and as “very high” if ∆4 was larger than
0.90 [77]. Statistical significance was set at p < 0.05, and marginal significance at p < 0.10.
3. Results
3.1. Frequency of Small Forest Rodents
On 215 camera trap-nights, 19,138 analysable videos (319.0 h) were taken within
the two years of survey. 102 camera trap-nights were conducted in the first mast year,
and 113 in the non-mast year. At least one small forest rodent was detected on 15,931
1 min videos (265.5 h). Using the maximum number of either simultaneously visible
bank voles or Apodemus mice per camera trap-night as a measure of frequency, population
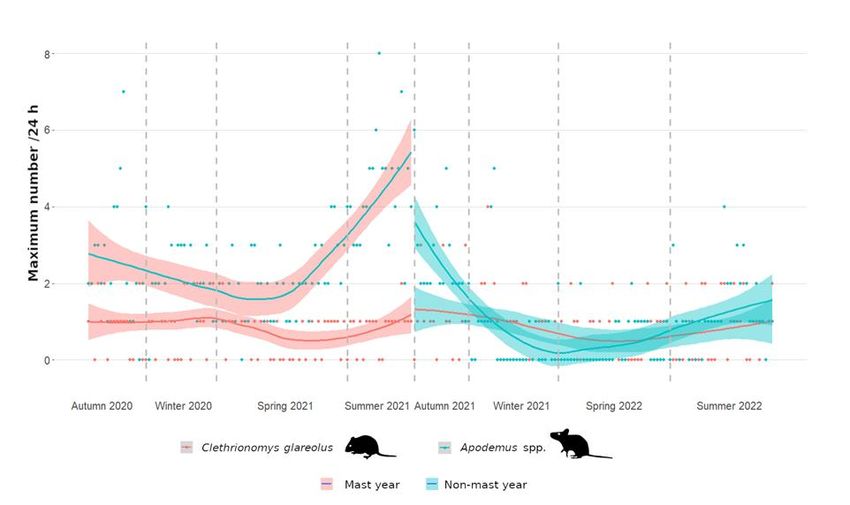
trends between the mast year and the non-mast year could be compared (Figure 1). The
frequency of bank voles did not differ between the mast year and the non-mast year
(U = 5965, p = 0.63, rg = 0.035). Bank voles showed a tendency to be more frequent after
the reproductive phase from spring into summer in both survey years, but three, or in a
single case four, individuals visible on the same video always represented an exception.
In contrast, Apodemus mice were significantly more frequent in the mast year compared
to the non-mast year (U = 9297, p < 0.001, rg = 0.612). In the mast year, Apodemus mice
overwintered in high numbers and reached their population peak the following summer.
Regularly up to five, and exceptionally up to eight, individuals could be counted on
the videos at the same time. In the non-mast year, the population collapsed totally and
recovered only slowly towards the end of the study, in the second summer. In winter and
spring of the non-mast year, Apodemus mice were not detected on many camera trap-nights
and were rarer than bank voles during this phase (Figure 1).
3.2. Activity Patterns of Small Forest Rodents
Activity patterns were analysed using maximum rodent numbers per 30 min intervals.
According to Hermans–Rasson tests, activity patterns were significantly different from
random in the bank voles (t = 25.58, p = 0.01) and Apodemus mice (t = 161.49, p = 0.01).
This was also valid when the mast year (bank voles: t = 25.79, p < 0.001, Apodemus mice:
t = 1159.60, p < 0.001) and the non-mast year (bank voles: t = 30.98, p < 0.001, Apodemus
mice: t = 464.90, p < 0.001) were tested separately. Bank voles showed a predominance of
diurnal activity in the mast year (56.5%), while nocturnal activity predominated in the non-
mast year (62.6%). The association between diurnal activity and mast year was significant
(χ2 (1) = 47.029, p < 0.001, Cramérs V = 0.188). Apodemus mice were overall much less
diurnal than bank voles (mast year: 6.3%, non-mast year: 2.7%), but again, diurnal activity
was significantly higher in the mast year (χ2 (1) = 13.14, p < 0.001, Cramérs V = 0.072).
Over both study years, the “medium” overlap of activity rhythms between bank voles andAnimals 2023, 13, 981 6 of 13
Animals 2023, 13, x FOR PEER REVIEW 6 of 13
Apodemus mice was 59% (∆4 = 0.591, 95% CIs = 0.564–0.619). The overlap was noticeably
lower in the mast year (∆4 = 0.501, 95% CIs = 0.463–0.538) than in the non-mast year
(∆4 = 0.647, 95% CIs = 0.609–0.684).
Figure 1. Population development of small forest rodents in an inner-alpine mixed forest. The
Figure 1. Population development of small forest rodents in an inner-alpine mixed forest. The fre-
frequency
quency of Apodemus
of Apodemus mice mice increased
increased significantly
significantly inmast
in the the mast
year,year, whereas
whereas thisthis
waswas
not not
the the
casecase
for for
bank voles, Clethrionomys glareolus. In the non-mast year, the population of Apodemus mice
bank voles, Clethrionomys glareolus. In the non-mast year, the population of Apodemus mice col- collapsed.
lapsed.
3.3. Temporal Niche of Bank Voles
A multivariate
3.2. Activity logistic
Patterns of Small regression
Forest Rodentswas used to investigate the dichotomous activity
pattern of Clethrionomys glareolus against seven extrinsic predictors (Table 1). The prob-
Activity patterns were analysed using maximum rodent numbers per 30 min inter-
ability of nocturnal activity was significantly increased in the non-mast year (p < 0.001,
vals. According to Hermans–Rasson tests, activity patterns were significantly different
OR = 2.329). Overall precipitation during the night significantly reduced activity (p < 0.001,
from random in the bank voles (t = 25.58, p = 0.01) and Apodemus mice (t = 161.49, p = 0.01).
OR = 0.932). A marginal significant effect was caused by lunar illumination (p = 0.051,
This was also valid when the mast year (bank voles: t = 25.79, p < 0.001, Apodemus mice: t
OR = 0.997) and the activity of Apodemus mice (p = 0.060, OR = 0.992), both of which led to
= 1159.60, p < 0.001)
a tendency towardsandincreased
the non-mast yearactivity.
daytime (bank voles: t = 30.98, p < 0.001, Apodemus mice:
t = 464.90, p < 0.001) were tested separately. Bank voles showed a predominance of diurnal
activity
Table in the mast
1. Effects year environmental
of seven (56.5%), whilepredictors
nocturnalon activity predominated
the nocturnal in the
activity of bank non-mast
voles, Clethriono-
year (62.6%). The
mys glareolus. association between diurnal activity and mast year was significant (χ2
(1) = 47.029, p < 0.001, Cramérs V = 0.188). Apodemus mice were overall much less diurnal
than bankStandard
Estimate voles (mast
Erroryear: Z-Score
6.3%, non-mast year: 2.7%),
p-Values Odds but again, diurnal
Ratio 2.50% activity was
97.50%
(Intercept) significantly higher
0.186 0.163 in the mast year (χ (1)0.255
1.139
2 = 13.14, p < 0.001,
1.205 Cramérs0.874 V = 0.072).1.660
Over
Non-mast both study years,
0.845 0.132the “medium” 6.426overlapAnimals 2023, 13, 981 7 of 13
(U = 32,337, p = 0.008, rg = −0.130), but in a significant decrease in nocturnal records of
bank voles in the non-mast year (U = 86,762, p < 0.001, rg = 0.215).
3.4. Intraguild Aggression
Apodemus mice and bank voles were only seen together on 350 videos out of 15,931
recordings, with at least one of these small forest rodents filmed (2.20%). Encounters were
about equally frequent in the non-mast year and in the mast year, although in the latter
Apodemus mice (but not bank voles) were much more frequent (Table 2, comp. Figure 1).
In the mast year, Apodemus mice were less aggressive, whereas in the non-mast year bank
voles more often avoided Apodemus mice or were actively chased away from the food by
them (χ2 (2) = 7.240, p = 0.027, Cramérs V = 0.144).
Table 2. Aggressive behaviour of Apodemus mice towards bank voles, Clethrionomys glareolus, distin-
guished between the mast year and the non-mast year.
No Aggression Avoidance Chase Total
Mast Year 74 (43.0%) 77 (44.8%) 21 (12.2%) 172 (49.1%)
Non-mast Year 52 (29.2%) 99 (55.6%) 27 (15.1%) 178 (50.9%)
4. Discussion
Rodent communities are considerably shaped by interspecific competition for limited
resources and temporal partitioning is one possible dimension of niche separation [16,78,79].
We studied whether bank voles show an inverse activity pattern compared to the superior
Apodemus mice. There are important, but contradictory, studies on this question [4,20,21,
25,80–89], and there is still no consensus on the influence of interference competition on
the chronoecology of the bank voles due to the use of different methods (e.g., capture data,
cafeteria experiments, and camera-trapping), the incomplete inclusion of confounding
variables, the lack of year-round studies and the limited knowledge on the frequency,
as well as nature of direct confrontations with Apodemus mice. We tried to approach
this issue with a large sample size of videos from camera traps in an inner-alpine mixed
forest, which were baited to increase the number of observations [20]. We assume that
our smell or handling hardly delayed the use of the bait, because (diurnal) bank voles
regularly appeared in the first half hour after setting up the camera trap (19.2 ± 7.66 sd min,
minimum 3 min, n = 10).
Assumption 1: An important requirement to test competition among species is met
if at least one species is particularly common [9]. We expected the massive increase of
small forest rodents in the seed mast year. In agreement with many other studies [10], this
first assumption was fulfilled in Apodemus mice. In contrast, bank voles remained at low
population levels throughout the study. Such relatively stable populations have been found,
for example, in the transition zone between the northern cyclic and the southern outbreak
populations in the so-called “limes norrlandicus” in Scandinavia [40], in undergrowth-poor
forest stands of the central European lowlands [42] and on low-productivity study areas
of the inner Alps [90]. For our research question, this ideally meant a quasi-experimental
variation in the frequency of the dominant competitor, with the frequency of the inferior
species remaining approximately constant. The small number of bank voles also renders
(largely unknown) intraspecific effects unlikely to outweigh interspecific competition [91].
Assumption 2: Albeit Apodemus mice were more diurnal in the mast year, they were
almost exclusively nocturnal overall. This is in agreement with numerous findings on
activity patterns within the genus Apodemus [4,21], which we could not distinguish at
a species level in our study. Bank voles showed more variability, with diurnal activity
predominating in the mast year, but nocturnal activity predominating in the non-mast
year. Accordingly, the overlap in activity rhythms between the two small rodent taxa was
much smaller in the mast year, when Apodemus mice were common. This suggests anAnimals 2023, 13, 981 8 of 13
avoidance strategy, which, however, remains a vague assumption without the inclusion of
other possible explanatory variables.
Prediction 1: We predicted an inverse activity pattern of bank voles with an increased
presence of Apodemus mice, controlling for a set of other possible explanatory variables.
The analyses revealed a close relationship between the frequency of Apodemus mice and the
diurnal activity of Clethrionomys glareolus, particularly and significantly in the mast year.
This indirect measure strongly suggests a temporal partitioning effect due to intraguild
competition, because none of the numerous additional explanatory variables describe the
same relationship: overnight precipitation reduced bank vole nocturnal activity, regardless
of the frequency of Apodemus mice. This is consistent with findings in other studies [3,82],
but contrary evidence exists [2] and causes are not fully understood (e.g., thermoregulatory
costs, interspecific concurrence, and predation risk). Lunar illumination led to an overall
marginally increased diurnal activity of the bank voles, but interestingly with different
effect directions in the mast and the non-mast years. While bank vole night activity
increased significantly with lunar illumination in the mast year, it was significantly reduced
in the non-mast year. We suggest an association to intraguild competition in this case
as well, because Apodemus mice are specifically known for their lunar phobia [3,4,19].
Therefore, bank voles might take advantage of the relatively competition-free period of
bright lunar nights, as Apodemus mice reduce their activity even at high densities due to
bright illumination. At low Apodemus mice densities, bank voles may use even the darkest
hours to avoid visual predators [92,93]. This would explain the contrasting findings on this
physical factor in other studies [3,4,19].
For the non-significant predictor cover, we assume too much uniformity between
camera trap locations. To the bank voles’ disadvantage [9,18,26,42,94–96], our study
forest was generally poor in cover below the tree layer, which is probably reflected in
the comparatively low frequency of this species, even in the mast year [62,97]. We de-
tected very few predators on our video recordings (n = 7). All were nocturnal, almost
exclusively mammalian predators (85.7%) and occurred primarily in the non-mast year
(87.5%), where a time-delayed population increase of these generalist, non-migratory,
and territorial predator species can be expected as a consequence of dispersal. Therefore,
it is even unlikely that indirect cues from predators, such urine, faeces and anal gland
secretions [8,93,98–101], led to increased daytime activity in bank voles in the mast year. In
addition, there was also no discernible correlation between the minimum temperature and
the behaviour of the bank voles. In other studies, the effects of temperature were small as
well or counteracting [2,3,82].
Finally, it is particularly important to note that the independent variable mast year/non-
mast year itself is significantly correlated with the diurnal activity of bank voles. Since
a test for multicollinearity showed that predictors of the multivariate logistic regression
model were not redundant (each variance inflation factor < 2), there must be an influence
on bank vole diurnal activity pattern beyond the mere frequency of Apodemus mice. Having
excluded the most common alternative variables, we surmise an additional influence of the
other two niche axes: habitat and food. It is well known that the dietary niche between these
small forest rodents overlaps much more in mast years, and high densities of Apodemus
mice make spatial separation more difficult for the bank voles, even at the microhabitat
level [41,102,103]. We therefore assume that under such ecological preconditions, the di-
urnal activity becomes particularly important for Clethrionomys glareolus. Due to a similar
food choice and the “omnipresent” Apodemus mice, raised fitness costs of diurnal activity
(e.g., increased predation risk [104,105]) can no longer be compensated for at night.
Prediction 2: The two taxa encountered each other very rarely in front of the camera
traps. The prediction that a higher number of Apodemus mice would lead to increased
intraguild aggression towards bank voles was not confirmed. In the non-mast year, despite
increased aggressiveness of Apodemus mice, no inverse activity pattern of bank voles anal-
ogous to the mast year was observed. Thus, the greater number of Apodemus mice had a
stronger negative impact on bank voles in the mast year than could be compensated byAnimals 2023, 13, 981 9 of 13
improved food supply. As the number of encounters did not differ between the mast year,
with numerous Apodemus mice, and the non-mast year, we assume an active avoidance
strategy by the bank voles. Given the long co-evolution of these small forest rodents, it
would be unlikely that dominance relationships need to be actively re-established per-
manently. Other authors, too, could not find evidence for persistent aggressive meetings
between Apodemus mice and bank voles [20,25,27]. In the non-mast year, the few adult
Apodemus mice (as far as assessable by body size) were particularly aggressive, probably in
the struggle for scarce food resources.
5. Conclusions
Our studies showed that a high frequency of the almost exclusively nocturnal Apode-
mus mice led to increased diurnal activity of the bank voles, and this could not be explained
by an extensive set of alternative predictors. We assume that at high Apodemus mice densi-
ties, increased separation along the time axis by bank voles must occur, as at night, there is
no longer sufficient relief of intraguild competition in all niche dimensions: time, microhab-
itat and diet. The “omnipresence” and intimidation by Apodemus mice is satisfactory for the
bank voles’ inverse activity pattern, without the need for frequent aggressive meetings. For
rigorous quantification of the exclusive influence of Apodemus mice on the activity budget
of bank voles, studies in areas where the two genera do not live syntopically, and extensive,
capture-mark-recapture or removal experiments simultaneous to the camera trap survey,
would be purposeful.
Author Contributions: Conceptualization, R.P. (Remo Probst) and R.P. (Renate Probst); methodology,
R.P. (Remo Probst); software, R.P. (Renate Probst); validation, R.P. (Renate Probst); formal analysis,
R.P. (Remo Probst); investigation, R.P. (Remo Probst) and R.P. (Renate Probst); resources, R.P. (Remo
Probst); data curation, R.P. (Renate Probst); writing—original draft preparation, R.P. (Remo Probst);
writing—review and editing, R.P. (Remo Probst). All authors have read and agreed to the published
version of the manuscript.
Funding: This research received no external funding.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: Not applicable.
Acknowledgments: We thank N. Nau for the opportunity to conduct the study on his property. We
are very grateful to M. Wunder for GIS work, F. Klein for statistical analyses, S. Glatz-Jorde for a
vegetation assessment and A. Wunder for improving our English.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. Kryštufek, B.; Tesakov, A.S.; Lebedev, V.S.; Bannikova, A.A.; Abramson, N.I.; Shenbrot, G. Back to the future: The proper name for
red-backed voles is Clethrionomys Tilesius and not Myodes Pallas. Mammalia 2020, 84, 214–217. [CrossRef]
2. Vickery, W.L.; Bider, J.R. The influence of weather on rodent activity. J. Mammal. 1981, 62, 140–145. [CrossRef]
3. Wróbel, A.; Bogdziewicz, M. It is raining mice and voles: Which weather conditions influence the activity of Apodemus flavicollis
and Myodes glareolus? Eur. J. Wildl. Res. 2015, 61, 475–478. [CrossRef]
4. Viviano, A.; Scarfò, M.; Mori, E. Temporal partitioning between forest-dwelling small rodents in a mediterranean deciduous
woodland. Animals 2022, 12, 279. [CrossRef]
5. Orrock, J.L.; Danielson, B.J. Temperature and cloud cover, but not predator urine, affect winter foraging of mice. Ethology 2009,
115, 641–648. [CrossRef]
6. Hernández, M.C.; Navarro-Castilla, Á.; Wilsterman, K.; Bentley, G.E.; Barja, I. When food access is challenging: Evidence of wood
mice ability to balance energy budget under predation risk and physiological stress reactions. Behav. Ecol. Sociobiol. 2019, 73, 145.
[CrossRef]
7. Butet, A.; Delettre, Y.R. Diet differentiation between European arvicoline and murine rodents. Acta Theriol. 2011, 56, 297–304.
[CrossRef]
8. J˛edrzejewski, W.; Rychlik, L.; J˛edrzejewska, B. Responses of bank voles to odours of seven species of predators: Experimental
data and their relevance to natural predator-vole relationships. Oikos 1993, 68, 251–257. [CrossRef]Animals 2023, 13, 981 10 of 13
9. Amori, G.; Castigliani, V.; Locasciulli, O.; Luiselli, L. Long-term density fluctuations and microhabitat use of sympatric Apodemus
flavicollis and Myodes glareolus in central Italy. Community Ecol. 2015, 16, 196–205. [CrossRef]
10. Pucek, Z.; J˛edrzejewski, W.; J˛edrzejewska, B.; Pucek, M. Rodent population dynamics in a primeval deciduous forest (Białowieża
National Park) in relation to weather, seed crop, and predation. Acta Theriol. 1993, 38, 199–232. [CrossRef]
11. Eccard, J.A.; Ylönen, H. Interspecific competition in small Rodents: From populations to individuals. Evol. Ecol. 2003, 17, 423–440.
[CrossRef]
12. Andreassen, H.P.; Sundell, J.; Ecke, F.; Halle, S.; Haapakoski, M.; Henttonen, H.; Huitu, O.; Jacob, J.; Johnsen, K.; Koskela, E.; et al.
Population cycles and outbreaks of small rodents: Ten essential questions we still need to solve. Oecologia 2021, 195, 601–622.
[CrossRef] [PubMed]
13. Grant, P.R. Interspecific competition among rodents. Annu. Rev. Ecol. Syst. 1972, 3, 79–106. [CrossRef]
14. Schoener, T.W. Field experiments on interspecific competition. Am. Nat. 1983, 122, 240–285. [CrossRef]
15. Halle, S.; Stenseth, N.C. Chronoecology: New light through old windows—A conclusion. In Acitivity Patterns in Small Mammals.
An Ecological Approach; Halle, S., Stenseth, N.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 273–284.
16. Schoener, T.W. Resource Partitioning in ecological communities: Research on how similar species divide resources helps reveal
the natural regulation of species diversity. Science 1974, 185, 27–39. [CrossRef]
17. Baláž, I.; Jakab, I.; Tulis, F.; Ambros, M. Spatial density of two sympatric species yellow-necked mouse Apodemus flavicollis and
bank vole Clethrionomys glareolus in different environment. Folia Oecologica 2016, 43, 121–128.
18. Hille, S.M.; Mortelliti, A. Microhabitat partitioning of Apodemus flavicollis and Myodes glareolus in the sub-montane Alps: A
preliminary assessment. Hystrix 2011, 21, 157–163. [CrossRef]
19. Buesching, C.D.; Newman, C.; Twell, R.; Macdonald, D.W. Reasons for arboreality in wood mice Apodemus sylvaticus and bank
voles Myodes glareolus. Mammal. Biol. 2008, 73, 318–324. [CrossRef]
20. Andrzejewski, R.; Olszewski, J. Social behaviour and interspecific relations in Apodemus flavicollis (Melchior, 1834) and Clethriono-
mys glareolus (Schreber, 1780). Acta Theriol. 1963, 10, 155–168. [CrossRef]
21. Greenwood, P.J. Timing of activity of the bank vole Clethrionomys glareolus and the wood mouse Apodemus sylvaticus in a deciduous
woodland. Oikos 1978, 31, 123–127. [CrossRef]
22. Sozio, G.; Mortelliti, A. Empirical evaluation of the strength of interspecific competition in shaping small mammal communities
in fragmented landscapes. Landsc. Ecol. 2016, 31, 775–789. [CrossRef]
23. Park, O. Nocturnalism—The development of a problem. Ecol. Monogr. 1940, 10, 485–536. [CrossRef]
24. Carothers, J.H.; Jaksic, F.M. Time as a niche difference: The role of interference competition. Oikos 1984, 42, 403–406. [CrossRef]
25. Grüm, L.; Bujalska, G. Bank voles and yellow-necked mice: What are interrelations between them? Pol. J. Ecol. 2000, 48, 141–145.
26. Casula, P.; Luiselli, L.; Amori, G. Which population density affects home ranges of co-occurring rodents? Basic Appl. Ecol. 2019,
34, 46–54. [CrossRef]
27. Bartolommei, P.; Gasperini, S.; Bonacchi, A.; Manzo, E.; Cozzolino, R. Multiple Captures as indicator of social tolerance in a guild
of terrestrial rodents. Mammal. Biol. 2018, 93, 169–172. [CrossRef]
28. Niethammer, J.; Krapp, F. Handbuch der Säugetiere Europas: Sciuridae, Castoridae, Gliridae, Muridae; Akademische Verlagsgesellschaft:
Wiesbaden, Germany, 1978.
29. Niethammer, J.; Krapp, F. Handbuch der Säugetiere Europas: Cricetidae, Arvicolidae, Zapodidae, Spalacidae, Hystricidae, Capromyidae;
Akademische Verlagsgesellschaft: Wiesbaden, Germany, 1982.
30. Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A Review of
camera Trapping for conservation behaviour research. Remote Sens. Ecol. Conserv. 2017, 3, 109–122. [CrossRef]
31. Rovero, F.; Zimmermann, F. Camera Trapping for Wildlife Research; Pelagic Publishing: Exeter, UK, 2016.
32. Lashley, M.A.; Cove, M.V.; Chitwood, M.C.; Penido, G.; Gardner, B.; DePerno, C.S.; Moorman, C.E. Estimating wildlife activity
curves: Comparison of methods and sample size. Sci. Rep. 2018, 8, 4173. [CrossRef]
33. Caravaggi, A.; Gatta, M.; Vallely, M.-C.; Hogg, K.; Freeman, M.; Fadaei, E.; Dick, J.T.A.; Montgomery, W.I.; Reid, N.; Tosh, D.G.
Seasonal and predator-prey effects on circadian activity of free-ranging mammals revealed by camera traps. PeerJ 2018, 6, e5827.
[CrossRef]
34. Nardotto, A. Living with the enemy: Activity rhythms of the red fox and some potential preys in an urban environment. Nat.
Hist. Sci. 2022, 9, 63–66. [CrossRef]
35. Wearn, O.R.; Glover-Kapfer, P. Snap happy: Camera traps are an effective sampling tool when compared with alternative methods.
Royal Soc. Open Sci. 2019, 6, 181748. [CrossRef]
36. Mills, C.A.; Godley, B.J.; Hodgson, D.J. Take only photographs, leave only footprints: Novel applications of non-invasive survey
methods for rapid detection of small, arboreal animals. PLoS ONE 2016, 11, e0146142. [CrossRef]
37. Ostfeld, R.S.; Keesing, F. Pulsed resources and community dynamics of consumers in terrestrial ecosystems. Trends Ecol. Evol.
2000, 15, 232–237. [CrossRef]
38. Yang, L.H.; Bastow, J.L.; Spence, K.O.; Wright, A.N. What can we learn from resource pulses? Ecology 2008, 89, 621–634. [CrossRef]
39. Yang, L.H.; Edwards, K.F.; Byrnes, J.E.; Bastow, J.L.; Wright, A.N.; Spence, K.O. A meta-analysis of resource pulse–consumer
interactions. Ecol. Monogr. 2010, 80, 125–151. [CrossRef]
40. Hansson, L.; J˛edrzejewska, B.; J˛edrzejewski, W. Regional differences in dynamics of bank vole populations in Europe. Pol. J. Ecol.
2000, 48, 163–177.Animals 2023, 13, 981 11 of 13
41. Selva, N.; Hobson, K.A.; Cortés-Avizanda, A.; Zalewski, A.; Donázar, J.A. Mast pulses shape trophic interactions between
fluctuating rodent populations in a primeval forest. PLoS ONE 2012, 7, e51267. [CrossRef] [PubMed]
42. Mazurkiewicz, M. Population dynamics and demography of the bank vole in different tree stands. Acta Theriol. 1991, 36, 207–227.
[CrossRef]
43. Zwander, V.H.; Aigner, S.; Koll, H. Der Pollenflug in Kärnten im Jahr 2020. Carinthia II 2021, 211, 163–177.
44. Geburek, T.; Hiess, K.; Litschauer, R.; Milasowszky, N. Temporal pollen pattern in temperate trees: Expedience or fate? Oikos
2012, 121, 1603–1612. [CrossRef]
45. Kasprzyk, I.; Ortyl, B.; Dulska-Jeż, A. Relationships among weather parameters, airborne pollen and seed crops of Fagus and
Quercus in Poland. Agric. For. Meteorol. 2014, 197, 111–122. [CrossRef]
46. Reil, D.; Imholt, C.; Eccard, J.A.; Jacob, J. Beech fructification and bank vole population dynamics—Combined analyses of
promoters of human Puumala virus infections in Germany. PLoS ONE 2015, 10, e0134124. [CrossRef] [PubMed]
47. Ascari, L.; Siniscalco, C.; Palestini, G.; Lisperguer, M.J.; Suarez Huerta, E.; De Gregorio, T.; Bregaglio, S. Relationships between
yield and pollen concentrations in Chilean hazelnut orchards. Eur. J. Agron. 2020, 115, 126036. [CrossRef]
48. Lanszki, J.; Zalewski, A.; Horváth, G. Comparison of red fox Vulpes vulpes and pine marten Martes martes food habits in a
deciduous forest in Hungary. Wildlife Biol. 2007, 13, 258–271. [CrossRef]
49. Luka, V.; Riegert, J. Apodemus mice as the main prey that determines reproductive output of tawny owl (Strix aluco) in central
Europe. Popul. Ecol. 2018, 60, 237–249. [CrossRef]
50. Jensen, T.S. Seed-seed predator interactions of European beech, Fagus silvatica and forest rodents, Clethrionomys glareolus and
Apodemus flavicollis. Oikos 1985, 44, 149–156. [CrossRef]
51. Nopp-Mayr, U.; Kempter, I.; Muralt, G.; Gratzer, G. Seed survival on experimental dishes in a central European old-growth
mixed-species forest—Effects of predator guilds, tree masting and small mammal population dynamics. Oikos 2012, 121, 337–346.
[CrossRef]
52. Zwolak, R.; Crone, E.E. Quantifying the outcome of plant-granivore interactions. Oikos 2012, 121, 20–27. [CrossRef]
53. Imholt, C.; Reil, D.; Plašil, P.; Rödiger, K.; Jacob, J. Long-term population patterns of rodents and associated damage in German
forestry. Pest Manag. Sci. 2017, 73, 332–340. [CrossRef]
54. Suchomel, J.; Šipoš, J.; Ouředníčková, J.; Skalský, M.; Heroldová, M. Bark gnawing by rodents in orchards during the growing
season—Can we detect relation with forest damages? Agronomy 2022, 12, 251. [CrossRef]
55. Heyman, P.; Mele, R.V.; Smajlovic, L.; Dobly, A.; Cochez, C.; Vandenvelde, C. Association between habitat and prevalence of
Hantavirus infections in bank voles (Myodes glareolus) and wood mice (Apodemus sylvaticus). Vector Borne Zoonotic Dis. 2009,
9, 141–146. [CrossRef]
56. Zöldi, V.; Papp, T.; Reiczigel, J.; Egyed, L. Bank voles show high seropositivity rates in a natural TBEV focus in Hungary. Infect.
Dis. 2015, 47, 178–181. [CrossRef]
57. Schmitt, M. Das Apodemus-Problem. Eulen Rundblick 2021, 71, 127–129.
58. Dubey, S.; Michaux, J.; Brünner, H.; Hutterer, R.; Vogel, P. False phylogenies on wood mice due to cryptic cytochrome-b
pseudogene. Mol. Phylogenet. Evol. 2009, 50, 633–641. [CrossRef]
59. Groenenberg, D.S.J.; Dekker, R.W.R.J. A mouse’s tail: How to settle an insurance dispute. Forensic Sci. Int. 2011, 207, e24–e27.
[CrossRef]
60. Martin Cerezo, M.L.; Kucka, M.; Zub, K.; Chan, Y.F.; Bryk, J. Population structure of Apodemus flavicollis and comparison to
Apodemus sylvaticus in northern Poland based on RAD-Seq. BMC Genom. 2020, 21, 241. [CrossRef] [PubMed]
61. Ancillotto, L.; Mori, E.; Sozio, G.; Solano, E.; Bertolino, S.; Russo, D. A novel approach to field identification of cryptic Apodemus
wood mice: Calls differ more than morphology. Mamm. Rev. 2017, 47, 6–10. [CrossRef]
62. Gasperini, S.; Mortelliti, A.; Bartolommei, P.; Bonacchi, A.; Manzo, E.; Cozzolino, R. Effects of forest management on density and
survival in three forest rodent species. For. Ecol. Manag. 2016, 382, 151–160. [CrossRef]
63. Spitzenberger, F. Die Säugetierfauna Österreichs; Grüne Reihe des Bundesministeriums für Land- und Forstwirtschaft, Umwelt und
Wasserwirtschaft: Graz, Austria, 2001.
64. Willner, W.; Grabherr, G. Die Wälder und Gebüsche Österreichs: Ein Bestimmungswerk mit Tabellen; Springer: Berlin/Heidelberg,
Germany, 2007.
65. Soné, K.; Kohno, A. Acorn hoarding by the field Mouse, Apodemus speciosus Temminck (Rodentia: Muridae). J. For. Res. 1999, 4,
167–175. [CrossRef]
66. Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’Brien, T.; O’Connell, A.; Sanderson, J.; Swann, D.E.; Tobler, M.; et al.
Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [CrossRef]
67. Rooney, S.M.; Wolfe, A.; Hayden, T.J. Autocorrelated data in telemetry studies: Time to independence and the problem of
behavioural effects. Mamm. Rev. 1998, 28, 89–98. [CrossRef]
68. Korn, H. Changes in home range size during growth and maturation of the wood mouse (Apodemus syIvaticus) and the bank vole
(Clethrionomys glareolus). Oecologia 1986, 68, 623–628. [CrossRef] [PubMed]
69. Mori, E.; Sangiovanni, G.; Corlatti, L. Gimme Shelter: The effect of rocks and moonlight on occupancy and activity pattern of an
endangered rodent, the garden dormouse Eliomys quercinus. Behav. Process. 2020, 170, 103999. [CrossRef] [PubMed]
70. Lešo, P.; Lešová, A.; Kropil, R.; Kaňuch, P. Response of the dominant rodent species to close-to-nature logging practices in a
temperate mixed forest. Ann. For. Res. 2016, 59, 259–268. [CrossRef]Animals 2023, 13, 981 12 of 13
71. Bowman, J.C.; Sleep, D.; Forbes, G.J.; Edwards, M. The association of small mammals with coarse woody debris at log and stand
scales. For. Ecol. Manag. 2000, 129, 119–124. [CrossRef]
72. Martinez, R.; Morato, S. Thigmotaxis and exploration in adult and pup rats. Rev. Etol. 2004, 6, 49–54.
73. Fitak, R.R.; Johnsen, S. Bringing the analysis of animal orientation data full circle: Model-based approaches with maximum
likelihood. J. Exp. Biol. 2017, 220, 3878–3882. [CrossRef]
74. Landler, L.; Ruxton, G.D.; Malkemper, E.P. The Hermans–Rasson test as a powerful alternative to the Rayleigh test for circular
statistics in biology. BMC Ecol. 2019, 19, 30. [CrossRef]
75. Ridout, M.S.; Linkie, M. Estimating Overlap of Daily Activity Patterns from Camera Trap Data. J. Agric. Biol. Environ. Stat. 2009,
14, 322–337. [CrossRef]
76. Meredith, M.; Ridout, M. Overview of the Overlap Package. 2021. Available online: https://cran.r-project.org/web/packages/
overlap/vignettes/overlap.pdf (accessed on 14 December 2022).
77. Mazza, G.; Marraccini, D.; Mori, E.; Priori, S.; Marianelli, L.; Roversi, P.F.; Gargani, E. Assessment of color response and
activity rhythms of the invasive black planthopper Ricania speculum (Walker, 1851) using sticky traps. Bull. Entomol. Res. 2020,
110, 480–486. [CrossRef]
78. Finke, D.L.; Snyder, W.E. Niche partitioning increases resource exploitation by diverse communities. Science 2008, 321, 1488–1490.
[CrossRef] [PubMed]
79. Levine, S.H. Competitive interactions in ecosystems. Am. Nat. 1976, 110, 903–910. [CrossRef]
80. Savić, I.; Todorović, M.; Mikeš, M. Teriološka istraživanja u stacionarnim šumskim ekosistemima i agrobiocenozama. Ekologija
1976, 11, 167–179.
81. Gurnell, J. Woodland rodents communities. In The Ecology of Woodland Rodents; Flowerdew, J.R., Gurnell, J., Gipps, J.H.W., Eds.;
Symposia of the Zoological Society of London 55; Oxford University Press: Oxford, UK, 1985; pp. 377–411.
82. Brown, L.E. Field experiments on the activity of the small mammals, Apodemus, Clethrionomys and Microtus. Proc. Zool. Soc. Lond.
1956, 126, 549–564. [CrossRef]
83. Wójcik, J.M.; Wołk, K. The daily activity rhythm of two competitive rodents: Clethrionomys glareolus and Apodemus flavicollis. Acta
Theriol. 1985, 30, 241–258. [CrossRef]
84. Kikkawa, J. Movement, activity and distribution of the small rodents Clethrionomys glareolus and Apodemus sylvaticus in woodland.
J. Anim. Ecol. 1964, 33, 259–299. [CrossRef]
85. Bergstedt, B. Distribution, Reproduction, growth and dynamics of the rodent species Clethrionomys glareolus (Schreber), Apodemus
flavicollis (Melchior) and Apodemus sylvaticus (Linné) in southern Sweden. Oikos 1965, 16, 132–160. [CrossRef]
86. Miller, R.S.; Elton, C. Activity rhythms in the wood mouse, Apodemus sylvaticus and the bank vole, Clethrionomys glareolus. Proc.
Zool. Soc. Lond. 1955, 125, 505–519. [CrossRef]
87. Grodzinski, W. Seasonal changes in the circadian activity of small rodents. Ekol. Polska Ser. B 1963, 9, 3–17.
88. Ashby, K.R. Patterns of daily activity in mammals. Mamm. Rev. 1972, 1, 171–185. [CrossRef]
89. Dell’omo, G.; Shore, R.F.; Lipp, H.-P. An automated system, based on microchips, for monitoring individual activity in wild small
mammals. J. Exp. Zool. 1998, 280, 97–99. [CrossRef]
90. Yoccoz, N.G.; Mesnager, S.; Mesnager, S. Are alpine bank voles larger and more sexually dimorphic because adults survive better?
Oikos 1998, 82, 85–98. [CrossRef]
91. Connell, J.H. On the prevalence and relative importance of interspecific competition: Evidence from field experiments. Am. Nat.
1983, 122, 661–696. [CrossRef]
92. Molsher, R.L.; Gifford, E.J.; McIlroy, J.C. Temporal, spatial and individual variation in the diet of red foxes (Vulpes vulpes) in
central New South Wales. Wildl. Res. 2000, 27, 593–601. [CrossRef]
93. Orrock, J.L. Rodent foraging is affected by indirect, but not by direct, cues of predation risk. Behav. Ecol. 2004, 15, 433–437.
[CrossRef]
94. Kołakowski, M.; Jancewicz, E.; Kielan, E. Czasowa i przestrzenna aktywność nornicy rudej Myodes (Clethrionomys) glareolus i
myszy leśnej Apodemus flavicollis w siedlisku gradowym
˛ Puszczy Białowieskiej. Sylwan 2018, 162, 1029–1037.
95. Montgomery, W.I. Seasonal variation in numbers of Apodemus sylvaticus, A. flavicollis and Clethrionomys glareolus. J. Zool. 1979,
188, 283–286. [CrossRef]
96. Montgomery, W.I. The use of arboreal runways by the woodland rodents, Apodemus sylvaticus (L.), A. flavicollis (Melchior) and
Clethrionomys glareolus (Schreber). Mamm. Rev. 1980, 10, 189–195. [CrossRef]
97. Fasola, M.; Canova, L. Asymmetrical competition between the bank Vole and the wood Mouse, a removal experiment. Acta
Theriol. 2000, 45, 353–365. [CrossRef]
98. Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, I.S. The effects of predator odors in mammalian prey
species: A review of field and laboratory studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [CrossRef]
99. Navarro-Castilla, Á.; Barja, I. Does predation risk, through moon phase and predator cues, modulate food intake, antipredatory
and physiological responses in wood mice (Apodemus sylvaticus)? Behav. Ecol. Sociobiol. 2014, 68, 1505–1512. [CrossRef]
100. Navarro-Castilla, Á.; Barja, I.; Díaz, M. Foraging, feeding, and physiological stress responses of wild wood mice to increased
illumination and common genet cues. Curr. Zool. 2018, 64, 409–417. [CrossRef]
101. Randler, C.; Kalb, J. Predator avoidance behavior of nocturnal and diurnal rodents. Behav. Process. 2020, 179, 10421. [CrossRef]
[PubMed]Animals 2023, 13, 981 13 of 13
102. Gasperini, S.; Bonacchi, A.; Bartolommei, P.; Manzo, E.; Cozzolino, R. Seasonal cravings: Plant food preferences of syntopic small
mammals. Ethol. Ecol. Evol. 2018, 30, 12–25. [CrossRef]
103. Canova, L. Resource partitioning between the bank vole Clethrionomys glareolus and the wood mouse Apodemus sylvaticus in
woodland habitats. Ital. J. Zool. 1993, 60, 193–198. [CrossRef]
104. Daan, S.; Aschoff, J. Circadian contributions to survival. In Vertebrate Circadian Systems; Aschoff, J., Daan, S., Groos, G.A., Eds.;
Springer: Berlin/Heidelberg, Germany, 1982; pp. 305–321.
105. Halle, S. Diel pattern of predation risk in microtine rodents. Oikos 1993, 68, 510–518. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual
author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to
people or property resulting from any ideas, methods, instructions or products referred to in the content.You can also read

















































