High-altitude tree growth responses to climate change across the Hindu Kush Himalaya - Oxford University Press
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of
Plant Ecology
Research Article
High-altitude tree growth responses to climate change
across the Hindu Kush Himalaya
Lili Zheng1,2, , Narayan Prasad Gaire3,4 and Peili Shi1,2,*
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
1
Lhasa Plateau Ecosystem Research Station, Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic
Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China, 2College of Resources and
Environment, University of Chinese Academy of Sciences, Beijing 100190, China, 3CAS Key Laboratory of Tropical Forest Ecology,
Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, Mengla, Yunnan 666303, China, 4Central
Department of Environmental Science, Tribhuvan University, Kirtipur, Kathmandu, Nepal
*Corresponding author. E-mail: shipl@igsnrr.ac.cn
Handling Editor: Christian Schöb
Received: 30 December 2020, First Decision: 26 January 2021, Accepted: 10 March 2021, Online Publication: 2 April 2021
Abstract
Aims Rapid warming at high altitudes may lead to a higher sensitivity in tree growth to temperature. The key
factors constraining tree radial growth and to what extent regional tree growth has suffered from climatic changes
are unclear.
Methods Tree-ring width data were collected from 73 sites across the Hindu Kush Himalaya (HKH), including
three dominant genera (Abies, Juniperus and Picea) at high altitudes over 3000 m. Dynamic time warping was
introduced to develop subregional chronologies by considering the synchrony of annual tree growth among
different sites. We quantified the contribution of the climate variables, and analyzed the spatiotemporal variation
of the growth–climate relationship.
Important Findings The site chronologies were grouped into three clusters, corresponding to the three distinct
bioclimatic zones, i.e. the western HKH, central-eastern HKH and southeastern Tibetan Plateau (TP). Tree growth
was positively correlated to winter and spring precipitation in the drier western HKH, and to winter temperature
and spring precipitation in the humid southeastern TP. Tree growth was markedly constrained by the minimum
temperature, especially in winter, with its importance increasing from the west toward the east. As shown by
moving correlation analysis, the signal of winter temperature in tree growth was weakened in the western and
central-eastern HKH, while it was enhanced in the southeastern TP following rapid warming since the 1980s.
Our results highlight that continuous warming may cause forest recession due to warming-induced moisture
deficit in the western HKH, but forest expansion in the southeastern TP.
Keywords tree ring, high-altitude forests, key climate factors, tree growth–climate relationships, growth trends,
climate sensitivity
兴都库什喜马拉雅地区高海拔树木生长对气候变化的响应
摘要:高海拔地区快速升温可能导致树木对温度响应更为敏感,而限制高海拔地区树木生长的关键气候
因子以及气候变化对树木生长产生多大程度的影响尚不清楚。本研究在兴都库什喜马拉雅地区收集了73
个样点的树轮数据,包括3个优势属的树种(Abies属、Juniperus属和Picea属),样点海拔均在3000 m以上。
© The Author(s) 2021. Published by Oxford University Press on behalf of the Institute of Botany, Chinese Academy of Sciences and the Botanical Society of China.
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial License (http://creativecommons.org/
licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited.
For commercial re-use, please contact journals.permissions@oup.com
JOURNAL OF PLANT ECOLOGY | doi:10.1093/jpe/rtab035 829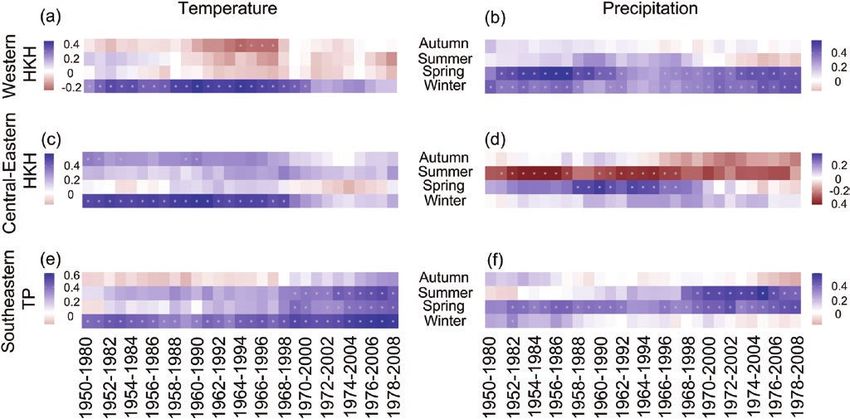
将时间动态规整(dynamic time warping)的方法用于建立亚区域年表,以考虑不同站点年表之间变化的同步
性。同时,定量分析了气候因子对树木生长的贡献以及树木生长与气候因子关系的时空动态。研究结果
发现,73个站点年表可以聚为3类,且与其所处的生物气候区相对应,即西喜马拉雅地区,中东喜马拉雅
地区和藏东南地区。在干旱的西喜马拉雅地区,树木生长与冬、春两季的降水呈正相关关系,而在湿润
的藏东南地区,树木生长与冬季温度和春季降水呈正相关关系。树木生长受最低温度的影响最大,特别
是冬季温度,其重要性从西到东呈现递增趋势。滑动窗口相关分析表明,在中西喜马拉雅地区,影响树
木生长的冬季温度信号在减弱,然而在藏东南地区该信号随着1980年以来的快速升温而增强。本研究结
果表明,若该地区升温持续,在西喜马拉雅地区可能会因变暖引起的水分亏缺而造成森林衰退,而在藏
东南地区因树木生长得益于变暖而使得森林扩张。
关键词:树轮,高海拔森林,关键气候因子,树木生长-气候的关系,生长趋势,气候敏感性
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
INTRODUCTION means. In practice, mean temperature is used most
widely because of its representativeness (Fan et al.
Climate warming is an undoubted fact (IPCC 2007,
2009; Zhao et al. 2011). Maximum temperature is
2014), and the most rapid warming rates have been
usually relevant in warm and drier areas, e.g. the
observed at high altitudes (Schickhoff et al. 2015).
It is expected that rapid warming will increase Mediterranean (Pellizzari et al. 2017), North America
temperature sensitivity of tree growth, and thus (Copenheaver et al. 2020) and Western Himalayas
substantially alter forest ecosystem composition (Borgaonkar et al. 2011). Minimum temperature
and functioning due to low temperature limitation is a critical factor closely related to cambial activity
at high altitudes (Körner 2012). However, the and the onset and end of xylogenesis (Li et al.
impact of such warming on tree growth is uncertain 2017a), and plays a significant role in cold and
because of different climatic drivers, environmental humid environments (Liang et al. 2010; Shi et al.
heterogeneity, species discrepancy and divergent 2020). Additionally, some studies have highlighted
responses (Babst et al. 2019; Girardin et al. 2016; the importance of growing season temperatures
Porter and Pisaric 2011; Rapp et al. 2012). Despite at high altitudes and high latitudes (Körner and
temperature as the most important climate factor Paulsen 2004) as well as summer, winter and spring
affecting tree growth, it is not clear what the key temperatures in determining tree growth (Francon
climatic factors constraining tree growth are, and et al. 2017; Panthi et al. 2017). Besides temperature,
to what extent regional tree growth has suffered precipitation is also an important factor affecting
from climate warming. Additionally, temperature or mediating tree growth, especially in drier
sensitivity of tree growth may be mediated and environments (Crawford et al. 2015). Several studies
vary with water conditions (Jevšenak et al. 2020). in the Himalayas have reported that tree growth at
Therefore, it is crucial to understand temperature high altitudes can be either limited by winter and/
sensitivities of tree growth and their temporal trends or spring temperatures (Aryal et al. 2020; Gaire et al.
under different precipitation regimes at a regional 2020) or controlled by premonsoon precipitation
scale. Quantifying tree growth responses to climatic (Dawadi et al. 2013; Gaire et al. 2017a; Liang et al.
variation along geographical gradients helps to better 2014; Yadav et al. 2014). However, little is known
understand the spatiotemporal patterns of climate- about the key factors and their possible interactions
driven tree growth and its climate sensitivity to in constraining tree growth at a regional scale.
ongoing climate change. Disentangling these factors has great implications to
Previous studies have exhibited that climate determine their roles in influencing the sensitivity of
factors may influence tree growth differently and tree growth to future climate change.
their effects may vary with environmental gradients. High-altitude ecosystems are extremely sensitive
Temperatures are generally considered as predictive and vulnerable to the most rapid climate warming
variables of tree growth responses in terms of seen in the past decades (IPCC 2007; Schickhoff et al.
maximum, minimum and mean temperatures, 2015). Tree growth at high altitudes may therefore
as well as the growing and nongrowing seasonal respond actively to ongoing climate warming,
830 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842
leading to tree growth enhancement (Shi et al. confounded by unbalanced precipitation variations
2020) and alpine treeline advancement (Harsch across the HKH?
et al. 2009). However, there are still inconsistent
results showing that a warming climate causes a
decrease in tree growth (Chen et al. 2017; Pellizzari MATERIALS AND METHODS
et al. 2017), tree dieback (Jump et al. 2017) and
even forest mortality (Allen and McDowell 2015), Study area and climate
which may be attributed to warming-induced The study area was located in the HKH, extending
water deficit. One of the reasons for such insistency more than 2500 km along the Himalayan mountain
is that most studies are performed in specific sites arc from the Pamirs in the west to the Hengduan
at relatively small scales such as local or landscape Mountains in the southeastern Tibetan Plateau (TP).
levels. It is not yet clear what the spatial patterns The complex topography and uplifting massif have
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
of warming-driven tree growth are in response to modified regional climate with distinct bioclimatic
climate change at a regional scale. zones. Temperatures are very low at high altitudes
The Hindu Kush Himalaya (HKH) has and even remain below 0 °C during summer, while
experienced the most rapid and elevation- dramatic climate warming has been observed since
dependent warming, along with an increase in the 1980s, especially in winter (Schickhoff et al.
extreme climate events, especially since the 1980s 2015). Rapid warming is particularly intense near
(Ren et al. 2017; Shrestha et al. 2012; Sun et al. 2017; high-altitude treelines (Liang et al. 2011; Schickhoff
Zhan et al. 2017). Large geographical gradients, et al. 2015). Precipitation decreases from the east
the most rapid warming climate and the highest to the west across the HKH (Kumar and Jain 2010;
high-altitude forests make it an excellent platform Ren et al. 2017; Sun et al. 2017; Zhan et al. 2017),
to study regional variation in the tree growth– although there are some high precipitation pocket
climate relationships under the scenario of climate areas in the central Himalayas due to the intense
change. The responses of tree growth to climate Indian monsoon. Despite an overall increase in
warming could present high spatial heterogeneity temperature and rapid warming since the 1980s,
due to the modulation of water conditions. With there is no significant trend in precipitation variations
climate warming, water stress could be inevitable across the entire HKH (see Fig. 3c). An analysis of the
in this region (Gaire et al. 2017a, 2019; Panthi et al. seasonal climatic variables showed that spring and
2017). Therefore, we cannot ignore the effects of summer precipitation increased overall, but autumn
warming-induced water deficit on tree growth in and winter precipitation decreased slightly in the
the HKH while considering the warming effect central and eastern Himalaya. Meanwhile, spring
on tree growth. Despite a large number of studies precipitation reduced, while summer precipitation
that have reported different results in drivers and increased significantly in the Western Himalaya
responses of tree growth at the stand and landscape (Shrestha et al. 2012).
scale, the climate sensitivity of tree growth across
Data sources and methods
the HKH region remains largely unknown.
Here, we compiled tree-ring growth data at Data sources
high altitudes in the HKH mountain arc to analyze We collected raw tree-ring width data from three
spatiotemporal variations in climate-driven tree dominant genera (i.e. Abies, Juniperus and Picea) at
growth patterns from 1950 to 2008. The extensive high altitudes across the HKH (24°–36° N, 70°–104°
dataset allows us to identify the key drivers of tree E) from the International Tree-Ring Data Bank
growth and quantify to what extent tree growth at (ITRDB; https://www.ncdc.noaa.gov/). In total, we
high altitudes has responded to climate warming selected 73 sites above 3000 m a.s.l. (Figs 1 and 2;
under different precipitation regimes at a regional Supplementary Table S1). Further exclusion rules
scale. Specifically, we address the following were applied (please see section ‘Clustering using the
questions: (i) What are the decisive climate factors dynamic time warping method’ in detail).
to drive the tree growth patterns at a subregional We downloaded TS v.4.01 gridded monthly
scale? (ii) What are the spatial patterns of the tree climate records including the maximum
growth sensitivities to climate warming? and (iii) temperature (Tmax), minimum temperature (Tmin),
Can temporal warming effects on tree growth be mean temperature (Tmean) and monthly
JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 831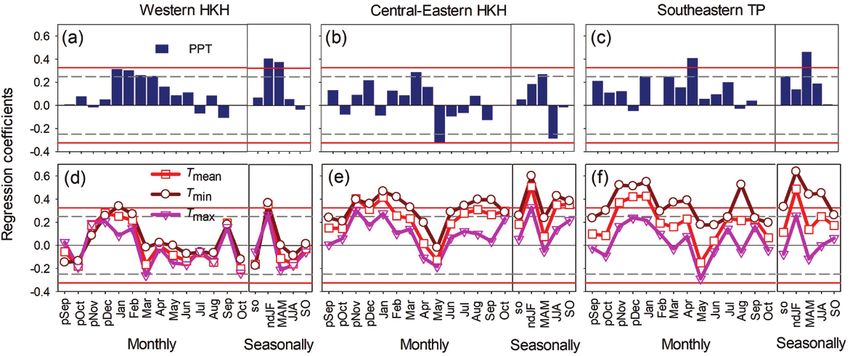
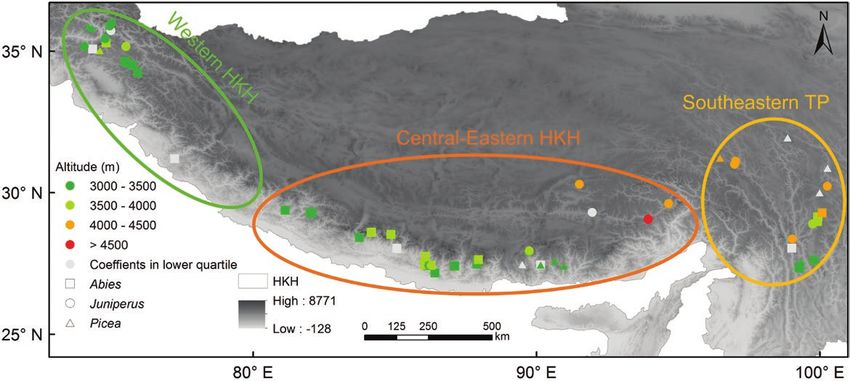
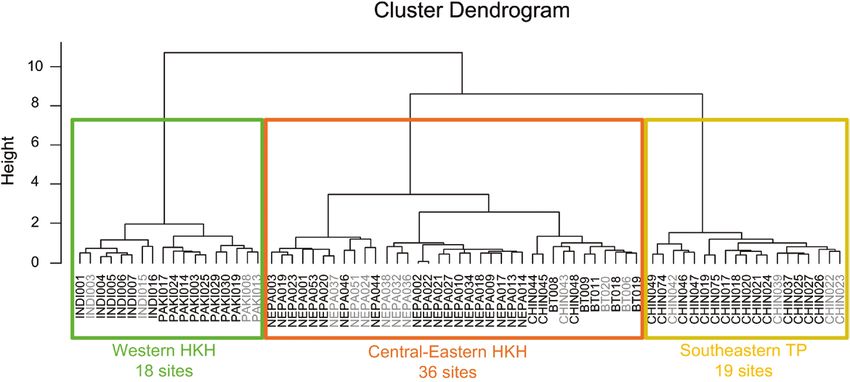
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021 Figure 1: Sites with tree-ring data and their clustering into three subregions in the HKH. Points with different colors from green to red indicate the altitudinal ranges of corresponding sites, in which the ones with light gray colors are excluded in the final analysis because their growth–winter temperature correlation coefficients are in the lower quartile. Different shapes of sites denote the three dominant genera, i.e. square for Abies, circle for Juniperus and triangle for Picea. Ellipses with different colors delineate the rough areas of three subregional chronologies derived from the DTW method, in which the green, orange and yellow represent subregions of the western HKH, central-eastern HKH and southeastern TP, respectively. Figure 2: Cluster dendrogram derived from the DTW method. Rectangles in three different colors indicate different clusters belonging to the three subregions in the HKH. Names of sampling sites in gray are those excluded before combining subregional composite chronologies due to their correlation coefficients of site chronologies with winter temperature being in the lower quartile within each cluster. precipitation from 1901 to 2016 from the Climate meteorological stations were established after the Research Unit (CRU; http://www.cru.uea.ac.uk/) 1950s, the accuracy of the interpolated climate data with a resolution of 0.5° × 0.5°. Then, we extracted has improved to a large extent since then. Thus, we climate data for each sampling site according to focused on investigating the tree growth–climate its geographic coordinates. Since the majority of relationships after the 1950s. 832 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842
Data processing that may vary in time or speed (Surhone et al. 2010).
At each site, an average of two cores from each This method has been widely used in econometrics
individual tree (if trees were sampled two cores (Franses and Wiemann 2020), disease monitoring
per tree in any study), and then detrended age- (Aghanavesi et al. 2020), speech recognition (Ismail
related growth trends using spline with a rigidity et al. 2020) and clustering of general time series digging
of 240 years, which equaled the average length (Ouivirach and Dailey 2010; Petitjean et al. 2011). Since
of the entire series (Fang et al. 2017), and finally each site chronology can be regarded as time-series
combined them into a site chronology using a data, it is recommended to try this method to detect
robust mean (Cook 1985). When assembling each the variations or similarities of site chronologies before
site chronology, we only kept the most reliable combining them into a single regional chronology.
period of the chronology, maintaining a subsample DTW is a method which computes the path
between two sequences and minimizing the distance
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
signal strength (SSS) greater than 0.85 (Fang et al.
2009, 2017). between the two sequences. This method allows for
an elastic shifting of the time axis to accommodate
Clustering using the dynamic time warping method sequences that are similar but out of phase (Keogh and
Dendrochronology is an interdisciplinary subject Ratanamahatana 2005). Thus, it can stretch or compress
widely applied in ecology, forestry and climatology, the sequences along the time axis (Zhang et al. 2018).
among others (Speer 2010). Therefore, a growing DTW was used to quantify synchronization among the
number of studies have been published on tree growth– 73 site chronologies by mapping each annual tree-ring
climate relationships using the traditional method of index of a site chronology to one or more neighboring
dendrochronology in the context of global warming. years’ tree-ring indices of another site chronology, to
This method combines individual tree-ring data into a find the minimum difference between the two site
site chronology following standard procedures (Cook chronologies (Zhang et al. 2018). Therefore, the DTW
1985; Fang et al. 2009; Liu et al. 2013). Indeed, this distance of two site chronologies can be calculated as:
Œ
traditional method has been widely and maturely used
2
D (X, rest(Y ))
to determine the limiting climatic factors of tree growth DTW(X, Y ) = D2 (f irst(X), f irst(Y )) + min D2 (rest(X), Y )
D2 (rest(X), rest(Y ))
(Rayback et al. 2017) and to reconstruct past climate
(Gaire et al. 2017a; Li et al. 2017b; Stahle et al. 2016). (1)
Such results have greatly improved our knowledge where X and Y represent two site chronologies, first(X)
of tree growth and its climatic sensitivity. However, a stands for the ‘current’ first element in X, and the
robust mean is generally used among different sites in rest(X) represent the remnant elements in X except for
order to combine site chronologies into a regional one the first(X). Obviously, this is an iteration procedure
using the traditional method (Panthi et al. 2017; Sohar in which the ‘min’ term repeats the DTW algorithm.
et al. 2017). In this way, the data process did not consider Meanwhile, the first and remaining sequences are
spatial distances and asynchrony among sampling sites. kept updating by the first() and rest() functions (Zhang
Pearson’s correlations are used to quantify confidence et al. 2018; Zhu and Shasha 2003). We then scaled the
levels when the chronologies from different sites are calculation of DTW(X, Y) and calculated the similarity
combined into the regional chronology (Lopez et al. between the two site chronologies as:
2019; Panthi et al. 2017). Consequently, many studies
DTW (X, Y ) = 1 − DTW(X, Y )
(2)
use the traditional method to detect the relationship
between regional climate and chronology whether where scaled DTW(X, Y) is used in Equation (2);
it is appropriate or not. In addition, some studies use DTW′(X, Y) stands for the similarity between the two
the principal component analysis to check the climatic site chronologies, i.e. the higher the value of DTW′(X,
response instead of simply taking the averaged regional Y) is, the more similar the two site chronologies are.
chronology (Cook et al. 2003; Yadav and Bhutiyani Simultaneously, the spatial distance between the
2013). Although some other combined methods have sites was also considered to keep spatial continuity,
been proposed to develop regional chronologies (Fang as indicated in Equation (3):
et al. 2017; Zhang et al. 2011), new interdisciplinary
Dis (X, Y ) = 1 − Dis(X, Y )
(3)
methods can further enrich the dendrochronology
toolbox. Dynamic time warping (DTW) is an algorithm where Dis(X, Y) refers to the scaled spatial distance
for calculating the similarity between two sequences (Euclidean distance) between site (X) and site (Y),
JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 833while Dis′(X, Y) represents the scaled inverse distance between the standardized subregional chronologies
between site (X) and site (Y), i.e. the higher the value and the standardized corresponding monthly
of Dis′(X, Y) is, the closer the two sites are located. and seasonal climate factors (Tmin, Tmean, Tmax and
We then averaged Dis′(X, Y) and DTW′(X, Y) precipitation) were then calculated. Because Tmin
to calculate a unique Index(X, Y) to conduct the was the key factor to limit tree growth (see section
following clustering hierarchy: ‘Relationships between the subregional chronologies
and climate factors’ and Fig. 4d–f), we simultaneously
DTW (X, Y ) + Dis (X, Y )
Index(X, Y ) =
(4) calculated the moving window correlations of
2
subregional chronologies with seasonal Tmin and
According to the results of the clustering hierarchy, precipitation, with a 31-year moving window and
we obtained three clusters (Figs 1 and 2). To expand 1-year window interval to check the temporal
the key climate signals and to minimize signal noise, variation in the growth–climate relationships.
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
we calculated monthly/seasonal growth–climate Following the convention in the studies across the
correlation coefficients for each site chronology and HKH, we treated March, April and May as spring;
found that winter temperature played an important June, July and August as summer; September and
role in all three clusters. Therefore, we excluded the October as autumn; with November and December
sampling sites in the lower quartile of the correlation of the previous year and January and February of the
coefficients with winter temperatures within each current year as winter.
cluster (Figs 1 and 2; Supplementary Table S1). The linear regressions were conducted using SPSS
Ultimately, 56 site chronologies were retained for the 20.0 software (IBM SPSS Statistics for Windows, IBM
final analyses. Corp., Armonk, NY, USA). DTW distance and moving
window correlations were determined using the ‘dtw’
Developing subregional chronologies
and ‘treeclim’ packages in the R language (R version
A robust mean was used to develop a subregional 3.5.2; Giorgino 2009; R Development Core Team
chronology for each cluster. The site chronologies 2018; Zang and Biondi 2015).
were then sorted into three subregions, namely,
western HKH chronology (14 sites), central-eastern
HKH chronology (27 sites) and southeastern TP RESULTS
chronology (15 sites) (see Figs 1 and 2; Supplementary
Table S1). Subregional chronologies of the HKH
We used the complete time series of tree-ring We developed three subregional chronologies
data during the detrending process and developing representing the western HKH (western India and
site chronologies. When tree growth–climate Pakistan), central-eastern HKH (eastern Tibet, most of
relationships were assessed at subregions, a part of Bhutan and Nepal) and southeastern TP (Hengduan
each subregional chronology was truncated to match Mountains) (Figs 1 and 2). In the past six decades,
the confident climate factors available (1950–2008). from 1950 to 2008, mean temperature anomalies
Because the tree-ring data have been published except annual precipitation displayed significantly
previously, and the SSS of all the site chronologies increasing trends in the three subregions (Fig. 3b
were above 0.85 after 1950, the data sources of the and c). Tree growth showed a decreasing, relatively
developed subregional chronologies were considered stable and overall increasing trend in the western
credible (Cook and Kairiukstis 1990). HKH, central-eastern HKH and southeastern TP,
respectively (Fig. 3a). Moreover, it is worth noting
Statistical analysis that tree growth showed a rapid increase after 1982
To verify the confidence levels (or representativeness) in the southeastern TP (Fig. 3a).
of the combined subregional chronologies and to
detect the influence of the different climatic variables Relationships between the subregional
on tree growth in the HKH, the relationships between chronologies and climate factors
the climate factors and the derived subregional Generally, tree growth was more responsive to
chronologies were tested. Prior to testing, the site temperature than to precipitation at high altitudes.
climate factors affiliated to the three clusters were Moreover, the minimum temperature rather than
also averaged to obtain three corresponding regional the mean temperature was the key climate variable
climate factors. The linear regression coefficients influencing tree growth in the HKH (Fig. 4).
834 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021 Figure 3: Variation of annual tree-ring indices (a), annual mean temperature anomalies (b) and annual total precipitation (c) in the three subregions of the HKH. Green, orange and yellow colors represent the subregions in the western HKH, central-eastern HKH and southeastern TP, respectively. R2 and P values of fitted linear curves in the bottom two panels are also given in corresponding colors. However, climate variables influenced subregional (September–October) and winter were found to tree growth differently. In the western HKH, the enhance tree growth (Fig. 4e). Exceptionally, tree lower monthly maximum temperature in March growth was associated positively with a wet spring but and higher precipitations in January through April, negatively with heavy rainfall in the summer (Fig. 4b). winter (previous November–February) and spring In the southeastern TP, the importance of the minimum (March–May) significantly favored tree growth and mean temperatures were both increased, with the (Fig. 4a and d). Meanwhile, the higher winter former being more critical in influencing tree growth. temperature was beneficial for tree growth regardless Specifically, tree growth was positively correlated with of its mean, maximum or minimum (Fig. 4d). In monthly temperatures in November and December the central-eastern HKH, tree-ring indices showed of the previous year, in January of the current year, higher correlations with the minimum temperature and the winter temperature (Fig. 4f). Higher monthly than mean and maximum temperatures. Higher precipitation in January, May and spring could favor temperatures in summer (June–August), autumn tree growth (Fig. 4c). JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 835
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021 Figure 4: Linear regression coefficients of subregional chronologies with monthly/seasonal climate factors for the western HKH (a and d), central-eastern HKH (b and e) and southeastern TP (c and f), upper panels for monthly/seasonal precipitation (abbreviated as PPT), lower panels for monthly/seasonal mean (Tmean), maximum (Tmax) and minimum (Tmin) temperatures. Time scale begins from previous September to current October. The small letter ‘p’ before month abbreviations means the month in the previous year. ’so’ stands for previous September and October (previous autumn), ‘ndJF’ for previous November through current February (winter), ‘MAM’ for March–May (spring), ‘JJA’ for June–August (summer) and ‘SO’ for September–October (autumn). Gray dashed and red solid lines show 95% and 99% confident intervals, respectively. Figure 5: Moving window correlations of seasonal climate-growth relationships for the three subregions in the HKH. (a), (c) and (e) for seasonal minimum temperature; (b), (d) and (f) for seasonal precipitation. Squares with different colors indicate moving correlation coefficients. Asterisks in the colored squares show significant correlation at P < 0.05 level. Moving window size is 31 years and interval is 1 year. Winter: previous November through current February, spring: March-May, summer: June-August, autumn: September-October. Temporal variations of the growth–climate positively correlated with winter temperature (Tmin) relationships and spring precipitation in the region as a whole, but Moving window correlation analysis revealed that varied remarkably within the three subregions (Fig. interannual growth–climate relationships were not 5). With increasing climate warming, the signal of stable during the entire study period in the three winter temperature was weakened in the western and subregions of the HKH. Overall, tree growth was central-eastern HKH, but was gradually intensified in 836 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842
the southeastern TP since the 1980s (Fig. 5a, c and e). Low temperatures are the critical limitation
Furthermore, spring and summer temperatures began to plant biochemical processes and physiological
to play positive roles in promoting tree growth in the activities (Larcher and Bauer 1981). There are several
southeastern TP since the 1980s as well (Fig. 5e). explanations for the decisive roles of minimum winter
For precipitation, tree radial growth was positively temperatures in influencing tree growth. First, the
correlated with spring precipitation across the HKH. low temperature, especially minimum temperature,
Tree growth likely responded to winter and spring appears to be closely related to restrictions on root
precipitation more in the western HKH than in the water uptake, despite most plants being dormant in
central-eastern HKH, and to summer precipitation winter (Jolly et al. 2005). At high altitudes, plants
negatively in central-eastern HKH (Fig. 5b and d). above the snow cover are exposed to low temperature
In contrast, the signal of spring precipitation in tree stress during the winter. Evergreen conifers with
growth was stable in the southeastern TP. Notably, small tracheids are especially prone to embolism
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
summer precipitation began to play a positive role in when cold stress induces low water potentials (Mayr
enhancing tree growth with the increasing climate et al. 2019). In addition, many field studies show that
warming after 1980 (Fig. 5f). low temperature reduces the biosynthetic activities of
plants. So early season temperature controls cambial
activity and total tree-ring width (Lenz et al. 2013).
DISCUSSION Moreover, colder winters can trigger physiological
All selected sites from the HKH mountain arc were desiccation and subsequent freezing injury in roots
roughly grouped into three subregions, corresponding and shoots, and consequently influence tree growth
to the three bioclimatic zones controlled by atmospheric in the growing season (Tranquillini 1979). The
circulation systems. The climate in the western HKH is minimum temperatures and long lengths of ambient
predominantly controlled by the westerlies, which are darkness during the nighttime may cause more
characterized by a drier climate with less precipitation frost if snow cover is scarce in drier environments.
(Schickhoff et al. 2015). Whereas the climate in the Therefore, warmer winters with a higher minimum
central-eastern HKH is dominated by the Indian temperature can increase an earlier snowmelt, in
monsoon, with heavy summer precipitation in most turn promoting the initiation of cambial activity,
of the areas. The southeastern TP is characterized by which may extend the growing season length and
moderate precipitation due to minor effects of the benefit the tree growth (Huang et al. 2019).
South Asian monsoon (Curio et al. 2015; Yao et al. However, winter warming can be a double-
2012). Findings showed that key climate variables edged sword; it can have both positive and negative
constraining tree growth at high altitudes shifted effects on tree growth. Considering low temperature
from temperature limitation in the southeastern TP limitations (Körner 2012), tree growth of montane
to more spring precipitation limitation in the western forests is known to be facilitated by warmer spring,
HKH, and subregional precipitation regimes greatly summer temperatures and extended growing seasons
mediated the tree growth responses to rapid climatic (Schickhoff et al. 2015). In contrast, our results
warming after the 1980s. indicated the opposite effects of ongoing warming
on tree growth between the western HKH and the
Temperature influence on high-altitude southeastern TP (Fig. 5a and c), which could be largely
tree growth attributed to different moisture conditions in these two
It has been a well-recognized notion that high- subregions (Panthi et al. 2017; Shi et al. 2020). Tree
altitude treelines are limited by growing season growth was commonly positively correlated with the
mean temperature (Körner and Paulsen 2004). Our winter minimum temperature. Following the rapid
findings show that tree growth is also affected by Tmin warming experienced since the 1980s, this positive
more than Tmean, especially following the change of effect of winter warming was continuously maintained
winter Tmin trends at high altitudes of the HKH (Figs and even enhanced in the southeastern TP. However,
4 and 5). Our results are consistent with previous such phenomena disappeared in the western and
studies showing that the positive effect of winter central-eastern HKH (Fig. 5a, c and e) where a dramatic
temperature is widely spread across the HKH (Cai increase of winter warming and a decrease of spring
et al. 2016; Chhetri and Cairns 2016; Fan et al. 2009; precipitation occurred (Shrestha et al. 2012). The
Gou et al. 2007; Huang et al. 2019; Rayback et al. legacy effect of winter warming-induced water deficit
2017; Zhang et al. 2015; Zhu et al. 2008). was also reflected by the negative role of maximum
JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 837temperature on tree growth in spring (e.g. March and limit tree growth (Fu et al. 2014; Lian et al. 2020).
May (Fig. 4d and f)). Previous studies have reported Moreover, winter warming can accelerate snow
the side effects of winter warming on tree growth. melting and reduce snow accumulation in winter
Thaw caused by winter warming can not only induce (You et al. 2016). Consequently, less snow may
winter desiccation, but may stimulate respiration impose water stress in the coming spring, while
more than photosynthesis at low temperatures, and spring precipitation can facilitate tree growth
consequently cause muted tree growth (Hamerlynck (Ahmad et al. 2020; Shah et al. 2018; Yadav 2013).
and Knapp 1996; Saxe et al. 2001). For example, If no warming occurs here, the relatively thicker
elevated winter temperature can increase spruce snowpack in the winter season would potentially
needle loss and reduce its growth in the following delay the start of the growing season and the
season (Skre and Nes 1996). Since the temperature in initiation of leaf sprouting, causing a decrease
winter is expected to increase more than in summer in tree growth. However, with the warming
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
under climate change (IPCC 2014), needle loss under climate, the melting snow could be an important
elevated winter temperatures may have a persistently water source for tree growth in the following
negative impact on forest productivity and cause spring season. Indeed, our results show winter
forest dieback. Facing warming winters, shallow roots precipitation (snow) can facilitate tree growth in
of conifers are limited by physiological drought caused the western HKH, suggesting that the positive effect
by damaged roots. Moreover, thaw–freeze cycles can of winter precipitation exceeds its negative effect
cause damage to fine roots due to a lack of protection on tree growth in this drier subregion (Figs 4a and
from insulating snowpack (Comeau et al. 2019; 5b). Interestingly, summer precipitation showed a
Schaberg et al. 2011). In addition, winter warming significantly negative effect before the 1990s in the
may stimulate earlier spring greening, which might moist central-eastern HKH (Figs 4b and 5b), probably
exacerbate summer soil drying (Lian et al. 2020). This due to dense cloud cover on rainy days blocking the
leads to moisture stress during the growing season, sunlight and causing a reduction in solar radiation
and eventual dieback and tree mortality (Hennon et al. (Grießinger and Bräuning 2006). Meanwhile, the
2012). air temperature on these consecutive rainy days
may be reduced during the monsoon season.
Precipitation influence on high-altitude Consequently, the photosynthesis of tree leaves
tree growth decreases, and the accumulation of carbohydrates
Although uneven seasonal precipitation induced reduces, and thus tree growth appears to slow
different effects on tree growth in high-altitude down. Previous tree-ring studies have detected
forests, tree radial growth is significantly correlated the heterogeneous and complex climate signals
with spring precipitation across the entire HKH of tree growth in the central-eastern HKH (Cook
(Figs 4a–c and 5b, d, f), in line with the results from et al. 2003; Gaire et al. 2017b, 2019; Panthi et al.
previous studies in the western HKH (Sohar et al. 2017; Sano et al. 2005). In the southeastern TP, the
2017; Yadav et al. 2014), central HKH (Dawadi et al. decisive role of spring precipitation in tree growth
2013; Gaire et al. 2017a; Panthi et al. 2017; Sigdel was shown despite relatively abundant precipitation
et al. 2018b; Tiwari et al. 2017) and eastern TP (Liang there. Furthermore, the positive signal of summer
et al. 2012; Ren et al. 2018; Zhang et al. 2003). This precipitation was also detected (Fig. 5f), likely due
is caused by a water deficit in spring resulting from to increasing warming-induced water requirements
evapotranspiration that cannot be compensated by for tree growth.
the scarce precipitation in the context of climate
warming. The asynchrony between increasing
warming and reduced moisture may cause a severe Regional tree growth trends in response to
water deficit and impede tree growth in spring (Ren recent climate change
et al. 2018). Premonsoon precipitation is an important The HKH has experienced a more rapid winter
water source to drive spring plant sprouting and the warming at high altitudes since the 1980s (Ren
onset of radial growth (Sigdel et al. 2018a). and Shrestha 2017; Singh et al. 2016). Our moving
In the western and central Himalayas, widespread correlation analysis indicated great differences
winter and spring warming (Shrestha et al. 2012) in the contributions of winter warming to the
may strengthen moisture stress for the early growing subregional tree growth, with a highly significant
season, exacerbate soil drying and subsequently contribution in the southeastern TP, whereas with
838 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842relatively lower contributions in the western and REFERENCES
central-eastern HKH since the 1980s, suggesting Aghanavesi S, Fleyeh H, Dougherty M (2020) Feasibility of
regionally divergent responses (Fig. 5). The using dynamic time warping to measure motor states in
contrasting results between subregions are largely Parkinson’s disease. J Sens 2020:3265795.
attributed to water stress imposed by rapidly Ahmad S, Zhu L, Yasmeen S, et al. (2020) A 424-year tree-
increasing climate warming (Bhutiyani et al. ring-based Palmer Drought Severity Index reconstruction
2007; Shrestha et al. 2012), in synchrony with a of Cedrus deodara D. Don from the Hindu Kush range
decrease in premonsoon precipitation, especially of Pakistan: linkages to ocean oscillations. Clim Past
16:783–798.
in northwestern Himalaya (Ren et al. 2017; Ren
Allen CD, McDowell NG (2015) Darcy’s law predicts
and Shrestha 2017). Climate warming-induced widespread forest mortality under climate warming. Nat
moisture deficit during the premonsoon season Clim Change 2015:H12B-02.
has become the major factor limiting tree growth,
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
Aryal S, Gaire NP, Pokhrel NR, et al. (2020) Spring season in
which is likely to spread more widely and bring Western Nepal Himalaya is not yet warming: a 400-year
about higher risks of growth decline and dieback. temperature reconstruction based on tree-ring widths of
Our results imply that continuous warming may Himalayan Hemlock (Tsuga dumosa). Atmosphere 11:132.
Babst F, Bouriaud O, Poulter B, et al. (2019) Twentieth century
cause forest recession in the western HKH, but
redistribution in climatic drivers of global tree growth. Sci
forest expansion in the southeastern TP.
Adv 5:eaat4313.
Bhutiyani MR, Kale VS, Pawar NJ (2007) Long-term trends in
maximum, minimum and mean annual air temperatures
CONCLUSIONS
across the Northwestern Himalaya during the twentieth
Our research found that tree growth at high century. Clim Change 85:159–177.
altitudes was mainly influenced by winter minimum Borgaonkar HP, Sikder AB, Ram S (2011) High altitude
temperatures and premonsoon precipitation across the forest sensitivity to the recent warming: a tree-ring
entire HKH. Despite the enhancement of tree growth analysis of conifers from Western Himalaya, India. Quat Int
236:158–166.
with increasing climate warming in the southeastern
Cai Q, Liu Y, Wang Y, et al. (2016) Recent warming evidence
TP, the benefits were offset by water stress induced by inferred from a tree-ring-based winter-half year minimum
rapid winter warming since the 1980s in the western temperature reconstruction in northwestern Yichang,
and central-eastern HKH. Based on our current South Central China, and its relation to the large-scale
results, we presume that continuous warming may circulation anomalies. Int J Biometeorol 60:1885–1896.
cause forest recession in the western HKH, but forest Chen L, Huang JG, Alam SA, et al. (2017) Drought causes
expansion in the southeastern TP. Our findings further reduced growth of trembling aspen in western Canada. Glob
advance the understanding of the responses of high- Change Biol 23:2887–2902.
Chhetri PK, Cairns DM (2016) Dendroclimatic response of
altitude tree growth to climate change.
Abies spectabilis at treeline ecotone of Barun Valley, eastern
Nepal Himalaya. J For Res 27:1163–1170.
Supplementary Material Comeau VM, Daniels LD, Knochenmus G, et al. (2019) Tree-
Supplementary material is available at Journal of rings reveal accelerated yellow-cedar decline with changes
Plant Ecology online. to winter climate after 1980. Forests 10:1085.
Table S1: Details of individual site chronologies from Cook ER (1985) A Time Series Analysis Approach to Tree Ring
the Hindu Kush Himalaya (HKH). Standardization. Arizona: University of Arizona.
Cook ER, Kairiukstis LA (1990) Methods of Dendrochronology:
Funding Applications in the Environmental Sciences. Dordrecht, The
This work was supported by National Natural Science Netherlands: Springer.
Foundation of China and ICIMOD Joint Research Cook ER, Krusic PJ, Jones PD (2003) Dendroclimatic signals
Project (no. 41661144045). in long tree-ring chronologies from the Himalayas of Nepal.
Int J Climatol 23:707–732.
Copenheaver CA, Shumaker KL, Butcher BM, et al. (2020)
Acknowledgements
Dendroclimatology of sugar maple (Acer saccharum):
We thank Huimeng Wang for her assistance with
climate-growth response in a late-successional species.
using dynamic time warping approach and appreciate Dendrochronologia 63:125747.
Yang Wang and Shoudong Zhao for their help in Crawford CJ, Griffin D, Kipfmueller KF (2015) Capturing
tree-ring data processing. season-specific precipitation signals in the northern Rocky
Conflict of interest statement. The authors declare that Mountains, USA, using earlywood and latewood tree rings.
they have no conflict of interest. J Geophys Res Biogeosci 120:428–440.
JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 839Curio J, Maussion F, Scherer D (2015) A 12-year high- Harsch MA, Hulme PE, McGlone MS, et al. (2009) Are treelines
resolution climatology of atmospheric water transport over advancing? A global meta-analysis of treeline response to
the Tibetan Plateau. Earth Syst Dyn 6:109–124. climate warming. Ecol Lett 12:1040–1049.
Dawadi B, Liang E, Tian L, et al. (2013) Pre-monsoon Hennon PE, D’Amore DV, Schaberg PG, et al. (2012) Shifting
precipitation signal in tree rings of timberline Betula utilis in climate, altered niche, and a dynamic conservation strategy
the central Himalayas. Quat Int 283:72–77. for yellow-cedar in the North Pacific Coastal Rainforest.
Fan ZX, Bräuning A, Cao KF, et al. (2009) Growth–climate BioScience 62:147–158.
responses of high-elevation conifers in the central Huang R, Zhu H, Liang E, et al. (2019) A tree ring-based winter
Hengduan Mountains, southwestern China. For Ecol Manage temperature reconstruction for the southeastern Tibetan
258:306–313. Plateau since 1340 CE. Clim Dyn 53:3221–3233.
Fang K, Gou X, Chen F, et al. (2009) Response of regional IPCC (2007) Climate change 2007: the physical science
tree-line forests to climate change: evidence from basis. In Solomon S, Qin D, Manning M, et al. (eds).
the northeastern Tibetan Plateau. Trees Struct Funct Contribution of Working Group I to the Fourth Assessment Report
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
23:1321–1329. of the Intergovernmental Panel on Climate Change. Cambridge
Fang K, Guo Z, Chen D, et al. (2017) Drought variation of University Press, Cambridge, UK.
western Chinese Loess Plateau since 1568 and its linkages IPCC (2014) Climate change 2014: synthesis report. In Core
with droughts in western North America. Clim Dyn Writing Team, Pachauri RK, Meyer LA (eds). Contribution
49:3839–3850. of Working Groups I, II and III to the Fifth Assessment Report
Francon L, Corona C, Roussel E, et al. (2017) Warm summers of the Intergovernmental Panel on Climate Change. Geneva,
and moderate winter precipitation boost Rhododendron Switzerland: IPCC.
ferrugineum L. growth in the Taillefer massif (French Alps). Ismail A, Abdlerazek S, El-Henawy IM (2020) Development
Sci Total Environ 586:1020–1031. of smart healthcare system based on speech recognition
Franses PH, Wiemann T (2020) Intertemporal similarity of using support vector machine and dynamic time warping.
economic time series: an application of dynamic time Sustainability 12:2403.
warping. Comput Econ 56:59–75. Jevšenak J, Tychkov I, Gričar J, et al. (2020) Growth-limiting
Fu YH, Piao S, Zhao H, et al. (2014) Unexpected role of winter factors and climate response variability in Norway spruce
precipitation in determining heat requirement for spring (Picea abies L.) along an elevation and precipitation gradients
vegetation green-up at northern middle and high latitudes. in Slovenia. Int J Biometeorol 65:311–324.
Glob Change Biol 20:3743–3755. Jolly WM, Nemani R, Running SW (2005) A generalized,
Gaire N, Bhuju D, Koirala M, et al. (2017a) Tree-ring based bioclimatic index to predict foliar phenology in response to
spring precipitation reconstruction in western Nepal climate. Glob Change Biol 11:619–632.
Himalaya since AD 1840. Dendrochronologia 42:21–30. Jump AS, Ruiz-Benito P, Greenwood S, et al. (2017) Structural
Gaire NP, Dhakal YR, Shah SK, et al. (2019) Drought (scPDSI) overshoot of tree growth with climate variability and the
reconstruction of trans-Himalayan region of central global spectrum of drought-induced forest dieback. Glob
Himalaya using Pinus wallichiana tree-rings. Palaeogeogr Change Biol 23:3742–3757.
Palaeoclimatol Palaeoecol 514:251–264. Keogh E, Ratanamahatana CA (2005) Exact indexing of
Gaire NP, Fan ZX, Shah SK, et al. (2020) Tree-ring record dynamic time warping. Knowl Inf Syst 7:358–386.
of winter temperature from Humla, Karnali, in central Körner C (2012) Alpine Treelines. Basel, Switzerland: Springer.
Himalaya: a 229 years-long perspective for recent warming Körner C, Paulsen J (2004) A world-wide study of high
trend. Geogr Ann A Phys Geogr 102:297–316. altitude treeline temperatures. J Biogeogr 31:713–732.
Gaire NP, Koirala M, Bhuju DR, et al. (2017b) Site- and species- Kumar V, Jain SK (2010) Trends in seasonal and annual
specific treeline responses to climatic variability in eastern rainfall and rainy days in Kashmir Valley in the last century.
Nepal Himalaya. Dendrochronologia 41:44–56. Quat Int 212:64–69.
Giorgino T (2009) Computing and visualizing dynamic time Larcher W, Bauer H (1981) Ecological significance of
warping alignments in R: the dtw package. J Stat Softw 31:1–24. resistance to low temperature. In Lange OL, Nobel PS,
Girardin MP, Bouriaud O, Hogg EH, et al. (2016) No growth Osmond CB, et al. (eds). Physiological Plant Ecology
stimulation of Canada’s boreal forest under half-century of I. Encyclopedia of Plant Physiology (New Series), Vol. 12.
combined warming and CO2 fertilization. Proc Natl Acad Sci Berlin, Germany: Springer.
U S A 113:E8406–E8414. Lenz A, Hoch G, Körner C (2013) Early season temperature
Gou X, Chen F, Jacoby G, et al. (2007) Rapid tree growth with controls cambial activity and total tree ring width at the
respect to the last 400 years in response to climate warming, alpine treeline. Plant Ecol Divers 6:365–375.
northeastern Tibetan Plateau. Int J Climatol 27:1497–1503. Li XX, Liang E, Gricar J, et al. (2017a) Critical minimum
Grießinger J, Bräuning A (2006) Late Holocene variations temperature limits xylogenesis and maintains treelines on
in monsoon intensity in the Tibetan Himalayan Region— the southeastern Tibetan Plateau. Sci Bull 62:804–812.
evidence from tree rings. J Geol Soc India 68:485–493. Li J, Shi J, Zhang DD, et al. (2017b) Moisture increase in
Hamerlynck E, Knapp AK (1996) Photosynthetic and stomatal response to high-altitude warming evidenced by tree-
responses to high temperature and light in two oaks at the rings on the southeastern Tibetan Plateau. Clim Dyn
western limit of their range. Tree Physiol 16:557–565. 48:649–660.
840 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842Lian X, Piao S, Li LZX, et al. (2020) Summer soil drying Ren P, Rossi S, Camarero JJ, et al. (2018) Critical temperature
exacerbated by earlier spring greening of northern and precipitation thresholds for the onset of xylogenesis of
vegetation. Sci Adv 6:eaax0255. Juniperus przewalskii in a semi-arid area of the north-eastern
Liang E, Dawadi B, Pederson N, et al. (2014) Is the growth of Tibetan Plateau. Ann Bot 121:617–624.
birch at the upper timberline in the Himalayas limited by Ren GY, Shrestha AB (2017) Climate change in the Hindu
moisture or by temperature? Ecology 95:2453–2465. Kush Himalaya. Adv Clim Change Res 8:137–140.
Liang E, Lu X, Ren P, et al. (2012) Annual increments of juniper Sano M, Furuta F, Kobayashi O, et al. (2005) Temperature
dwarf shrubs above the tree line on the central Tibetan variations since the mid-18th century for western Nepal,
Plateau: a useful climatic proxy. Ann Bot 109:721–728. as reconstructed from tree-ring width and density of Abies
Liang E, Wang Y, Eckstein D, et al. (2011) Little change in the spectabilis. Dendrochronologia 23:83–92.
fir tree-line position on the southeastern Tibetan Plateau Saxe H, Cannell MGR, Johnsen Ø, et al. (2001) Tree and forest
after 200 years of warming. New Phytol 190:760–769. functioning in response to global warming. New Phytol
Liang EY, Wang YF, Xu Y, et al. (2010) Growth variation in 149:369–399.
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
Abies georgei var. smithii along altitudinal gradients in the Schaberg PG, D’Amore DV, Hennon PE, et al. (2011) Do
Sygera Mountains, southeastern Tibetan Plateau. Trees limited cold tolerance and shallow depth of roots contribute
Struct Funct 24:363–373. to yellow-cedar decline? For Ecol Manage 262:2142–2150.
Liu J, Qin C, Kang S (2013) Growth response of Sabina tibetica Schickhoff U, Bobrowski M, Boehner J, et al. (2015) Do
to climate factors along an elevation gradient in south Tibet. Himalayan treelines respond to recent climate change?
Dendrochronologia 31:255–265. An evaluation of sensitivity indicators. Earth Syst Dyn
Lopez EL, Kerr SA, Sauchyn DJ, et al. (2019) Variation in tree 6:245–265.
growth sensitivity to moisture across a water-limited forest Shah SK, Pandey U, Mehrotra N (2018) Precipitation
landscape. Dendrochronologia 54:87–96. reconstruction for the Lidder Valley, Kashmir Himalaya
Mayr S, Schmid P, Rosner S (2019) Winter embolism and using tree-rings of Cedrus deodara. Int J Climatol
recovery in the conifer shrub Pinus mugo L. Forests 10:941. 38:E758–E773.
Ouivirach K, Dailey MN (2010) Clustering human behaviors Shi C, Schneider L, Hu Y, et al. (2020) Warming-induced
with dynamic time warping and hidden Markov models unprecedented high-elevation forest growth over the
for a video surveillance system. In: The 2010 ECTI monsoonal Tibetan Plateau. Environ Res Lett 15:054011.
International Conference on Electrical Engineering/Electronics, Shrestha UB, Gautam S, Bawa KS (2012) Widespread climate
Computer, Telecommunications and Information Technology change in the Himalayas and associated changes in local
(ECTI-CON2010). ecosystems. PLoS One 7:e36741.
Panthi S, Bräuning A, Zhou ZK, et al. (2017) Tree rings reveal Sigdel SR, Dawadi B, Camarero JJ, et al. (2018a) Moisture-
recent intensified spring drought in the central Himalaya, limited tree growth for a subtropical Himalayan conifer
Nepal. Glob Planet Change 157:26–34. forest in Western Nepal. Forests 9:340.
Pellizzari E, Camarero JJ, Gazol A, et al. (2017) Diverging Sigdel SR, Wang Y, Camarero JJ, et al. (2018b) Moisture-
shrub and tree growth from the Polar to the Mediterranean mediated responsiveness of treeline shifts to global warming
biomes across the European continent. Glob Change Biol in the Himalayas. Glob Change Biol 24:5549–5559.
23:3169–3180. Singh RB, Schickhoff U, Mal S (2016) Climate Change,
Petitjean F, Ketterlin A, Gançarski P (2011) A global averaging Glacier Response, and Vegetation Dynamics in the Himalaya.
method for dynamic time warping, with applications to Switzerland: Springer.
clustering. Pattern Recognit 44:678–693. Skre O, Nes K (1996) Combined effects of elevated winter
Porter TJ, Pisaric MFJ (2011) Temperature-growth divergence temperatures and CO2 on Norway spruce seedlings. Silva
in white spruce forests of Old Crow Flats, Yukon Territory, Fenn 30:135–143.
and adjacent regions of northwestern North America. Glob Sohar K, Altman J, Lehečková E, et al. (2017) Growth-climate
Change Biol 17:3418–3430. relationships of Himalayan conifers along elevational and
R Core Team (2018) R: A language and environment for statistical latitudinal gradients. Int J Climatol 37:2593–2605.
computing. Vienna, Austria: R Foundation for Statistical Speer JH (2010) Fundamentals of Tree Ring Research. Arizona:
Computing. University of Arizona Press.
Rapp JM, Silman MR, Clark JS, et al. (2012) Intra- and Stahle DW, Cook ER, Burnette DJ, et al. (2016) The Mexican
interspecific tree growth across a long altitudinal gradient Drought Atlas: tree-ring reconstructions of the soil moisture
in the Peruvian Andes. Ecology 93:2061–2072. balance during the late pre-Hispanic, colonial, and modern
Rayback SA, Shrestha KB, Hofgaard A (2017) Growth eras. Quat Sci Rev 149:34–60.
variable-specific moisture and temperature limitations Sun XB, Ren GY, Shrestha AB, et al. (2017) Changes in extreme
in co-occurring alpine tree and shrub species, central temperature events over the Hindu Kush Himalaya during
Himalayas, Nepal. Dendrochronologia 44:193–202. 1961–2015. Adv Clim Change Res 8:157–165.
Ren YY, Ren GY, Sun XB, et al. (2017) Observed changes in Surhone LM, Tennoe MT, Henssonow SF, et al. (2010) Dynamic
surface air temperature and precipitation in the Hindu Time Warping. Beau Bassin, Mauritius: Betascript Publishing.
Kush Himalayan region over the last 100-plus years. Adv Tiwari A, Fan ZX, Jump AS, et al. (2017) Gradual expansion
Clim Change Res 8:148–156. of moisture sensitive Abies spectabilis forest in the
JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842 841Trans-Himalayan zone of central Nepal associated with Zhang QB, Cheng G, Yao T, et al. (2003) A 2,326-year tree-ring
climate change. Dendrochronologia 41:34–43. record of climate variability on the northeastern Qinghai-
Tranquillini W (1979) Physiology Ecology of the Alpine Timberline— Tibetan Plateau. Geophys Res Lett 30:1739.
Tree Existence at High Altitudes with Special Reference to the Zhang Y, Shao X, Wilmking M (2011) Dynamic relationships
European Alps. Berlin, Germany: Springer. between Picea crassifolia growth and climate at upper
Yadav RR (2013) Tree ring-based seven-century drought treeline in the Qilian Mts., Northeast Tibetan Plateau,
records for the Western Himalaya, India. J Geophys Res Atmos China. Dendrochronologia 29:185–199.
118:4318–4325. Zhang X, Wang M, Liu K, et al. (2018) Using NDVI time series
Yadav RR, Bhutiyani MR (2013) Tree-ring-based snowfall to diagnose vegetation recovery after major earthquake
record for cold arid western Himalaya, India since A.D. based on dynamic time warping and lower bound distance.
1460. J Geophys Res Atmos 118:7516–7522. Ecol Indic 94:52–61.
Yadav RR, Misra KG, Kotlia BS, et al. (2014) Premonsoon Zhang RB, Yuan YJ, Wei WS, et al. (2015) Dendroclimatic
precipitation variability in Kumaon Himalaya, India over a reconstruction of autumn-winter mean minimum
Downloaded from https://academic.oup.com/jpe/article/14/5/829/6209400 by guest on 23 September 2021
perspective of ~300 years. Quat Int 325:213–219. temperature in the eastern Tibetan Plateau since 1600 AD.
Yao T, Thompson L, Wei Y, et al. (2012) Different glacier Dendrochronologia 33:1–7.
status with atmospheric circulations in Tibetan Plateau and Zhao W, Jiang Y, Dong M, et al. (2011) Relationship between
surroundings. Nat Clim Change 2:663–667. the radial growth of Picea meyeri and climate along elevations
You Q, Min J, Kang S (2016) Rapid warming in the Tibetan of the Luyashan Mountain in North-Central China. For Ecol
Plateau from observations and CMIP5 models in recent Manage 265:142–149.
decades. Int J Climatol 36:2660–2670. Zhu Y, Shasha D (2003) Warping indexes with envelope
Zang C, Biondi F (2015) treeclim: an R package for the transforms for query by Humming. In: Proceedings of the ACM
numerical calibration of proxy-climate relationships. SIGMOD International Conference on Management of Data.
Ecography 38:431–436. Zhu H, Zheng Y, Shao X, et al. (2008) Millennial temperature
Zhan YJ, Ren GY, Shrestha AB, et al. (2017) Changes in extreme reconstruction based on tree-ring widths of Qilian juniper
precipitation events over the Hindu Kush Himalayan region from Wulan, Qinghai Province, China. Chin Sci Bull
during 1961–2012. Adv Clim Change Res 8:166–175. 53:3914–3920.
842 JOURNAL OF PLANT ECOLOGY | 2021, 14:829–842You can also read

















































