Glucocorticoids coordinate changes in gut microbiome composition in wild North American red squirrels
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
www.nature.com/scientificreports
OPEN Glucocorticoids coordinate changes
in gut microbiome composition
in wild North American red
squirrels
Lauren Petrullo1*, Tiantian Ren2, Martin Wu2, Rudy Boonstra3, Rupert Palme4, Stan Boutin5,
Andrew G. McAdam6 & Ben Dantzer1,7*
The gut microbiome impacts host health and fitness, in part through the diversification of gut
metabolic function and pathogen protection. Elevations in glucocorticoids (GCs) appear to reduce gut
microbiome diversity in experimental studies, suggesting that a loss of microbial diversity may be a
negative consequence of increased GCs. However, given that ecological factors like food availability
and population density may independently influence both GCs and microbial diversity, understanding
how these factors structure the GC-microbiome relationship is crucial to interpreting its significance
in wild populations. Here, we used an ecological framework to investigate the relationship between
GCs and gut microbiome diversity in wild North American red squirrels (Tamiasciurus hudsonicus). As
expected, higher GCs predicted lower gut microbiome diversity and an increase in metabolic taxa.
Surprisingly, but in line with prior empirical studies on wild animals, gastrointestinal pathogens
decreased as GCs increased. Both dietary heterogeneity and an upcoming food pulse exhibited direct
effects on gut microbiome diversity, whereas conspecific density and reproductive activity impacted
diversity indirectly via changes in host GCs. Our results provide evidence of a gut–brain axis in wild
red squirrels and highlight the importance of situating the GC-gut microbiome relationship within an
ecological framework.
The intimate symbiosis between animals and their microbiomes has become a major area of focus for ani-
mal behavior, ecology, and evolution research over the last decade. In vertebrates, the gut microbiome inter-
acts strongly with other host physiological systems1. Gut microbiota are sensitive to changes in host immune
function2, brain development and behavior3,4, circadian rhythms5, and metabolism6. Beyond these effects,
the gut microbiome also responds to the host endocrine system. In wild female primates, reproductive hor-
mones like estrogen and progesterone are associated with differences in gut microbiome composition7,8. In
humans, both androgens and estrogens, as well as metabolic hormones like insulin, are linked to variation in gut
microbiota9,10. Such connections reflect a larger “gut–brain axis” through which the gut microbiota and nervous
system communicate11.
Recently, glucocorticoids (GCs) have emerged as a central component of the bidirectional gut–brain a xis12.
GCs are metabolic hormones produced via the activation of the hypothalamic–pituitary–adrenal axis. They
are involved in energy regulation and the physiological stress r esponse13, and can induce adaptive phenotypic
plasticity in response to environmental c hange14. For example, elevated GCs can enhance fitness by facilitating
transitions between life history stages15, supporting the energetic demands of reproduction16, and improving
survival in response to fluctuating temperatures and food a vailability17. GCs may also induce adaptive plasticity
in the gut microbiome, a host microbial community that responds rapidly to changes in ecology. Shifts in gut
microbiome composition can regulate energy balance as ambient temperatures rise and fall18, and can enhance
1
Department of Psychology, University of Michigan, Ann Arbor, MI 48108, USA. 2Department of Biology, University
of Virginia, Charlottesville, VA 22904, USA. 3Department of Biological Sciences, Centre for the Neurobiology of
Stress, University of Toronto Scarborough, Toronto, ON M1C 1A6, Canada. 4Unit of Physiology, Pathophysiology
and Experimental Endocrinology, Department of Biomedical Sciences, University of Veterinary Medicine Vienna,
Veterina ̈rplatz 1, 1210 Vienna, Austria. 5Department of Biological Sciences, University of Alberta, Edmonton,
AB T6G 2E9, Canada. 6Department of Ecology and Evolutionary Biology, University of Colorado, Boulder,
CO, USA. 7Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI 48108,
USA. *email: petrullo@umich.edu; dantzer@umich.edu
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 1
Vol.:(0123456789)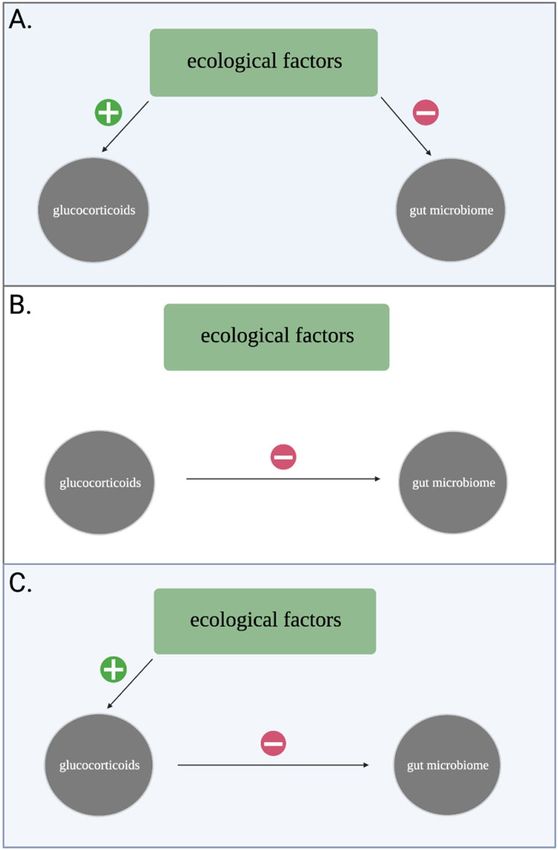
www.nature.com/scientificreports/
Figure 1. Conceptual model demonstrating how ecological factors structure the relationship between
glucocorticoids and the gut microbiome. (A) Environmental covariance results in direct effects on both variables
and diminishes a detectable effect of GCs on gut microbiome diversity. (B) Ecology does not influence either
variable and a direct effect of GCs on gut microbiome diversity is preserved. (C) Ecological factors influence gut
microbiome diversity indirectly via host GCs. Note that the three scenarios are not mutually exclusive, such that
a combination of direct (A) and indirect (B) effects may result in the appearance or absence of a relationship
between GCs and gut microbiome diversity.
Study Species N GCs Effect Ecological factors included?
Noguera et al. (2018)24 Gulls (Larus michahellis) 29 Experimental ↓ ɑ-diversity No
Stothart et al. (2019)26 Gray squirrels (Sciurus carolinensis) 29 Natural No effect No
Uren Webster et al. (2020)25 Atlantic salmon (Salmo salar) 168 Experimental ↓ ɑ-diversity No
Vlčková et al. (2018)27 Gorillas (Gorilla gorilla gorilla) 42 Natural No effect No
Table 1. Selected prior experimental and correlational studies on the relationship between glucocorticoids
and gut microbiome alpha diversity in captive and wild vertebrates.
digestion efficiency as an animal’s energetic demands increase (e.g., during reproduction) or as resource avail-
ability fluctuates19–21.
One measure of gut microbiome composition—alpha diversity, or the taxonomic diversity within a sin-
gle community—appears particularly sensitive to changes in host GCs. A taxonomically diverse microbiome
confers community stability and resilience, whereas a loss of diversity is presumed to have detrimental conse-
quences via increased host susceptibility to pathogenic infection22,23. Animal studies in which GCs have been
experimentally elevated have documented reduced gut microbiome diversity in response to elevated GCs24,25,
while studies in unmanipulated populations have found no r elationship26,27. This inconsistency may indicate that
the link between GCs and gut microbiome diversity is modified by ecological factors (Fig. 1), yet these are rarely
included in such analyses (Table 1). For example, an increase in food availability can cause transient elevations in
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 2
Vol:.(1234567890)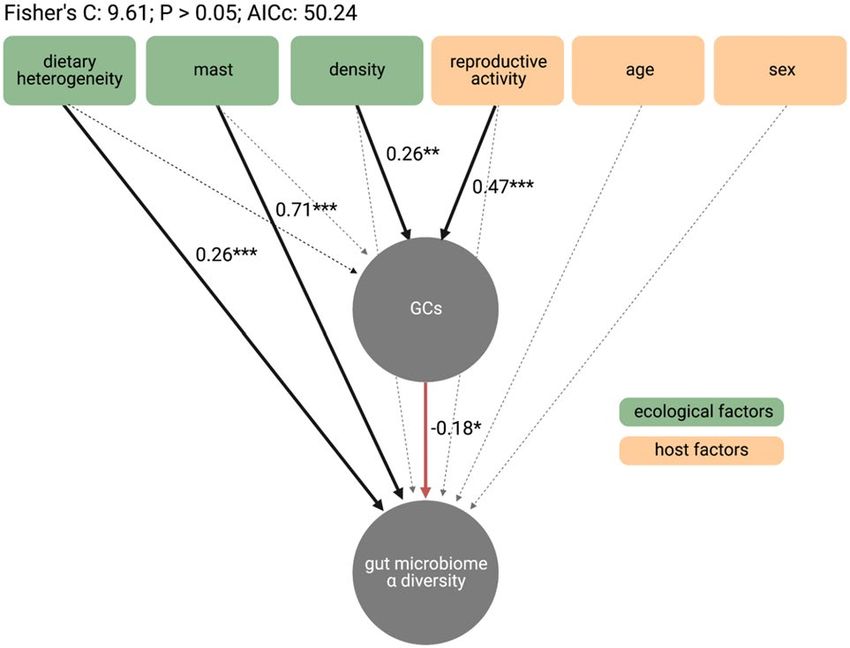
www.nature.com/scientificreports/
GCs if conspecific density also increases due to that elevation in food availability28. If elevated density results in
more frequent social interactions, it may enhance microbiome diversity directly via increased microbial trans-
mission among c onspecifics29. Such environmental covariance may drive the absence of a relationship between
GCs and microbiome diversity in unmanipulated populations (Fig. 1)30–32, necessitating an integrative approach
to determine how GCs impact the gut microbiome in wild animals.
In this study, we test the hypothesis that ecological factors structure the effects of GCs on gut microbiome
alpha diversity in wild North American red squirrels (Tamiasciurus hudsonicus) living in the Yukon, Canada.
Red squirrels are highly territorial animals that experience dramatic shifts in food availability and population
density as a result of fluctuations in their preferred food source, seeds from white spruce trees (Picea glauca)33.
Squirrels incorporate other food sources into their diet when seasonally available (e.g., fungi, bark, leaves, flow-
ers)34, resulting in changes in dietary diversity that may directly impact gut microbiome diversity. However,
spruce seeds comprise the majority of their d iet34 despite their episodic availability. Masting events occur every
4–6 years in white spruce, resulting in the production of a superabundance of cones containing seeds that become
available in the autumn. By contrast, few to no cones are available in non-mast y ears33,35. In anticipation of an
upcoming spruce mast, squirrels exhibit an extended breeding season and concomitant behavioral changes: ter-
ritoriality breaks down and conspecific interactions increase due to increased breeding frequency and infanticidal
behavior36,37. An upcoming spruce mast may thus exert direct positive effects on gut microbiome diversity via
more frequent social interactions, which leads to greater horizontal microbial t ransmission29.
Squirrel densities also fluctuate in parallel with food pulses, with densities at their lowest in the months prior
to a mast and highest in the spring following a m ast28,38. Although sociality is expected to increase gut microbi-
ome diversity in group-living animals , this effect may not occur in territorial species39. For example, elevated
29
conspecific densities result in increased frequency of long-range territorial vocalizations emitted by red squirrels
in our study p opulation40, which can in turn reduce interaction frequency by deterring territorial i ntrusions39,41.
Indeed, conspecific interactions in squirrels do not appear to vary with density, and the number of territorial
intruders has been both negatively c orrelated40 and u nrelated41 to density. Given that both actual and perceived
increases in density cause GC elevations in red s quirrels28,42, density may have indirect rather than direct effects
on microbiome diversity due to the psychosocial stress of anticipating greater c ompetition28.
We predicted to find an overall negative association between host GCs and gut microbiome alpha diversity,
along with an increase in pathogenic taxa and taxa involved in host metabolism with increasing GCs. Similar to
prior studies in wild animals (Table 1), we focused here on the unidirectional effects of GCs on microbial diversity
to understand gut–brain axis function in the absence of an experimental manipulation, and because previously
documented sensitivity of GCs to intrinsic and extrinsic factors in our p opulation28 suggests it may mediate
downstream effects on the microbiome. We used a multivariate structural equation modeling approach43,44 to
integrate GCs, the gut microbiome, and exogenous ecological and host biological variables into a single causal
network45. We tested a set of a priori hypothesized relationships related to the direct and indirect effects of dietary
heterogeneity, an upcoming spruce mast, and conspecific density on both GCs and gut microbiome diversity
(Fig. S1). We expected to find that dietary heterogeneity and an upcoming spruce mast would have direct posi-
tive effects on gut microbiome diversity. Conversely, we predicted that density would have an indirect negative
effect on diversity by way of GC elevations. We additionally included biological factors (reproductive activity,
sex, age) in our analysis, given their potential effects on GCs and microbiome composition46–48. We predicted
that reproductive activity had positive direct effects on both GCs and gut microbiome diversity, as a result of
increased energetic d emands49,50 and conspecific interactions, respectively. We also expected that older age would
predict lower microbiome diversity, and that males would exhibit greater microbiome diversity due to travel
across territories for multiple mating in the breeding s eason51.
Results
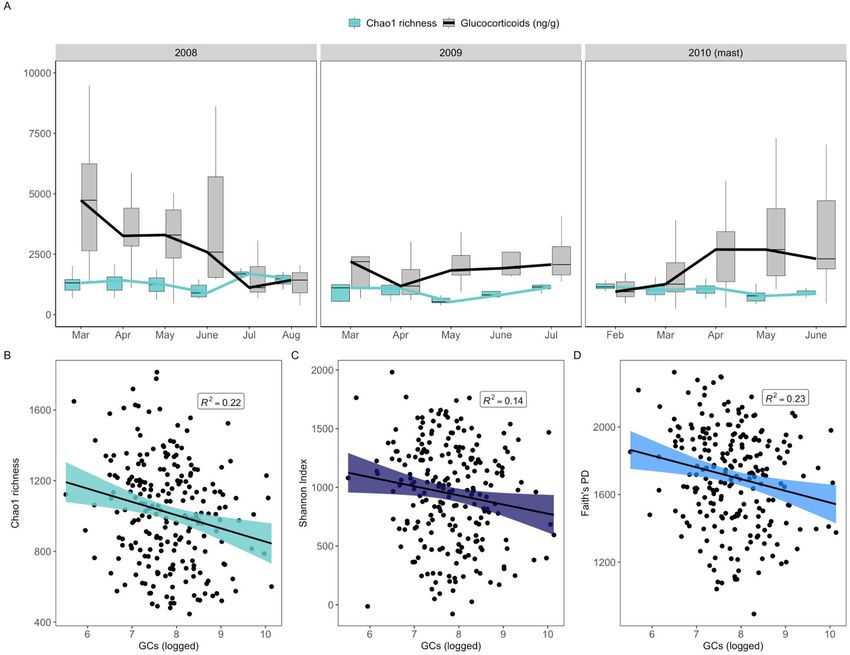
Gut microbiome diversity is negatively associated with glucocorticoids. Both gut microbiome
alpha diversity and GCs were highly variable across seasons in each of our sampling years, with gut microbial
diversity reaching its maxima during the summer months of July and August (Fig. 2A), coinciding with increased
dietary diversity34,52. GCs were highest in early spring (March), with the exception of the mast year of 2010 in
which GCs steadily increased across the first part of the year (Fig. 2A). Consistent with our predictions and in
line with prior studies in which GCs were experimentally manipulated (Table 1), GCs were negatively associated
with gut microbiome alpha diversity. Individuals with greater GC concentrations exhibited relatively lower taxo-
nomic diversity (i.e., species richness, Chao1: β = − 75.05 ± 25.91, t = − 2.90, P = 0.004; Fig. 2B). Greater GCs were
also associated with lower Shannon Indices, a composite measure of species richness and evenness (Shannon:
β = − 77.64 ± 36.00, t = − 2.16, P = 0.03, Fig. 2C), as well as lower phylogenetic diversity in the gut microbial com-
munity (Faith’s PD: β = − 66.97 ± 26.71, t = − 2.51, P = 0.01, Fig. 2D). The negative relationship between GCs and
gut microbiome alpha diversity was robust to individual variation in GC production, with higher individually-
averaged GCs similarly predicting lower species richness (β ± SE: − 0.027 ± 0.011, t = − 2.45, P = 0.02).
Glucocorticoids predict changes in gut microbial taxa. We constructed a series of negative binomial
linear mixed-effects models to determine how reduced gut microbiome alpha diversity was reflected in changes
at the taxonomic level and identify taxa whose relative abundances changed with increasing GCs. We found
that elevated GCs were associated with changes in gut microbiome composition at both the family (Fig. 3A)
and genus (Fig. 3B) levels. Increased GCs predicted shifts in the relative abundances of 15 bacterial families,
predominantly decreases in rare bacterial families (i.e., taxa that contribute < 0.01% relative abundance to the
microbial community) (Fig. 3A; Table S1). An exception was a reduction in Elusimicrobiaceae, which contrib-
uted an average of 0.13% relative abundance to the gut microbiome community (β = − 0.81, P FDR < 0.0001). By
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 3
Vol.:(0123456789)www.nature.com/scientificreports/
Figure 2. Host production of glucocorticoids predicts gut microbiome alpha diversity. (A) Boxplot and line
graph showing the opposing relationship between mean fecal glucocorticoid metabolites (GCs, scaled) and
median gut microbiome diversity (Chao1 richness) across each month of the three sampling years. Outliers
removed from plot for visualization purposes. (B) Partial residual plots (points represent individual samples)
showing the relationship between gut microbiome taxonomic richness (Chao1), (C) taxonomic richness and
evenness (Shannon Index), and (D) phylogenetic diversity (Faith’s PD) and matched fecal glucocorticoid
concentrations (GCs) (N = 227). R2 values (i.e., coefficients of determination) were obtained from linear mixed-
effect models.
contrast, elevated GCs were associated with an increase in Coriobacteriaceae (β = 0.57, PFDR = 0.007), Strepto-
coccaceae (β = 1.0, PFDR < 0.0001), Dermabacteraceae (β = 2.81, PFDR < 0.0001), and Ruminococcaceae (β = 0.12,
PFDR = 0.02) (Fig. 3A). Ruminococcaceae, a family of largely cellulolytic and fibrolytic bacteria, is an abundant
(~ 25% relative abundance) and core taxa in the red squirrel gut microbiome52.
At the genus level, the relative abundances of 22 bacterial genera were significantly reduced with increas-
ing host GCs. Similar to changes at the family level, the majority of bacterial genera reductions were rare taxa
(Fig. 3B) with the exception of Brochothrix (mean 0.02% relative abundance). Contrary to our predictions but
in line with a prior study on b irds24, we found that two potentially pathogenic genera—Yersinia (β = − 1.09,
PFDR = 0.004) and Salmonella (β = − 10.98, PFDR < 0.0001)54—decreased in relative abundance with increasing host
53
GCs. Conversely, a greater proportion of abundant taxa were found to increase with increasing GCs (Fig. 3B).
Greater host GCs predicted greater relative abundances of Clostridium (β = 0.28, PFDR = 0.008), Butyricicoccus
(β = 0.35, PFDR = 0.004), Oscillospira (β = 0.60, PFDR < 0.0001), YRC22 (β = 0.65, P
FDR = 0.004), and Lachnospira
(β = 0.67, PFDR = 0.02).
The gut‑brain axis mediates ecological effects on the gut microbiota. To determine how eco-
logical and host factors contributed to the effects of GCs on gut microbiome alpha diversity, we fit a structural
equation model (SEM) based on a set of a priori hypothesized pathways (Fig. S1). The SEM was constructed to
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 4
Vol:.(1234567890)www.nature.com/scientificreports/
Figure 3. Gut microbial taxa that shift with increasing host glucocorticoids. Barplots depict bacterial families
(A) and genera (B) whose relative abundance was significantly (Benjamini–Hochberg adjusted P < 0.05)
predicted by changes in host glucocorticoid concentrations. Bold taxa exhibited a mean relative abundance
> 0.01%. Effects of GCs reflect model estimates generated by negative binomial mixed models testing the effect
of GCs on the relative abundance of each bacterial taxa, controlling for collection date, food supplementation
status, and individual ID. Black bars represent a decrease in relative abundance with increasing GCs; grey bars
represent an increase with increasing GCs. Taxa depicted at the bottom of Panel B (Kribbella, Propionibacterium)
exhibited model estimates ~ 10× larger than the rest of the taxa and are therefore separated from the main plot
for visualization purposes.
test the relative direct and indirect effects of three ecological factors (dietary heterogeneity, an upcoming spruce
mast, and conspecific density) and three host factors (reproductive activity, age, and sex) on GCs and gut micro-
biome diversity. Overall, the SEM revealed direct and indirect pathways by which ecological and host factors
exert cascading effects on microbial diversity (Figure 4). In line with our predictions, dietary heterogeneity and
the presence of an upcoming spruce mast both exhibited direct effects on gut microbiome diversity, such that a
more heterogeneous diet (standardized β = 0.26, P < 0.001) and an upcoming spruce masting event (standard-
ized β = 0.71, P < 0.001) led to greater microbial diversity. Tests of directed separation revealed that there was
no effect of either dietary heterogeneity or an upcoming spruce mast on host GCs. By contrast, conspecific
density and reproductive activity indirectly, but not directly, affected gut microbiome diversity via changes to
host GCs. Higher squirrel densities (standardized β = 0.26, P = 0.006) and reproductive activity (standardized
β = 0.47, P < 0.001) predicted greater GC concentrations, which in turn reduced microbial diversity (standard-
ized β = − 0.18, P = 0.01). There was no effect of age or sex on gut microbiome diversity, and tests of directed
separation similarly found no effect of age or sex on host GCs.
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 5
Vol.:(0123456789)www.nature.com/scientificreports/
Figure 4. Ecology and host biology influence gut microbiome alpha diversity via changes in host
glucocorticoids. Structural equation model assessing direct and indirect effects of ecological and host factors
on glucocorticoids (GCs) and gut microbiome alpha diversity (Chao1 richness). Solid black arrows represent
significant positive paths; solid red arrows represent significant negative paths; dotted arrows represent non-
significant paths. Text labels indicate standardized beta estimates (i.e., effect sizes) and significance (P < 0.05*,
P < 0.01**, P < 0.001***) for each of the predicted pathways tested in the SEM.
Discussion
Determining if ecological factors structure the relationship between glucocorticoids (GCs) and gut microbiome
alpha diversity is crucial to interpreting the adaptive value of the GC-microbiome connection in wild animals.
Here, we demonstrate that the link between GCs and gut microbiome alpha diversity is robust to ecological fac-
tors that directly influence the gut microbiome in wild populations. Our findings additionally suggest that GCs
can integrate changes in ecology and host biology to induce plasticity in the gut microbiome.
Gut microbiome alpha diversity varied across seasons and peaked in summer, coinciding with the period of
eterogeneity34,52 (Fig. 2A). GCs exhibited greater variability overall: GCs reached their peak in
greatest dietary h
the early spring in non-mast years (2008, 2009), but in summer of the mast year (2010) (Fig. 2A). Despite this
variability, there was a significant negative association between GCs and gut microbiome diversity such that
individuals with greater GCs exhibited lower alpha diversity across three separate measures (species richness,
evenness, and phylogenetic diversity) (Fig. 2B–D). Though prior studies in unmanipulated populations did not
detect a relationship between GCs and gut microbiome alpha diversity (Table 1), but s ee55 for oral microbiome),
our results align with experimental studies in which GCs were manipulated. This suggests that gut–brain axis
communication, particularly between GCs and gut microbiome composition, is under strong selection in our
population.
As GCs increased, the taxa that decreased in the gut microbiome were overwhelmingly rare taxa that contrib-
uted < 0.01% in relative abundance to the overall community (e.g., Odoribacteriaceae, Sporichthyaceae) (Fig. 3).
This finding aligns with expectations about the effects of disturbances on gut microbiome composition from
an ecological perspective56. Resilience to microbial disturbances is greater among abundant taxa than non-
abundant taxa57, likely due to divergent patterns of colonization and succession. Abundant taxa retain their
position in microbial communities via selective filtering and by occupying core n iches56. By contrast, rare taxa
are incorporated into the microbiome largely via stochastic processes, though they contribute significantly to
community alpha diversity m easures58. A reduction in rare taxa with increasing GCs in our population may
therefore indicate that the effects of elevated GCs on rare bacteria in the gut mimic those expected by broader
microbial disturbances (e.g., antibiotics, infection)59.
A decrease in the relative abundance of rare bacteria may also serve to reorganize host metabolic priorities
through replacement by core taxa that can better support changes in host energetic demands. Overall, we found
that increases in host GCs were accompanied by increases in bacteria involved in host metabolism (Fig. 3). Both
Oscillospira, which correlates with the consumption of spruce buds in the late spring52, and Coriobacteriaceae, a
common rodent gut microbe involved in energy metabolism60, increased in relative abundance with increasing
GCs. In experimental settings, housing stress caused an increase in Coriobacteriaceae61, suggesting that it may
similarly contribute to maintaining energy balance in wild rodents facing challenging environmental conditions.
We additionally found that individuals with elevated GCs exhibited greater relative abundances of Ruminococ-
caceae, a bacterial family of cellulolytic and fibrolytic bacteria involved in dietary acclimation in wild a nimals20,62.
Together, our results suggest that in our study population, GCs may coordinate adaptive shifts in gut microbiome
composition in response to increased energetic demands, seasonal changes in diet, or both.
Resistance to pathogens has been proposed as one of the major evolutionary advantages conferred by host
microbial communities63 and butyrates, compounds produced via fermentation by microbiota in the large
intestine64, are particularly critical to preventing intestinal pathogen invasion65. We found that the butyrate-
producing bacteria Butyriciococcus (family Ruminococcaceae) and Clostridium were elevated in the red squirrel
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 6
Vol:.(1234567890)www.nature.com/scientificreports/
gut microbiome as GCs increased (Fig. 3). In line with this finding, elevated GCs were associated with lower
relative abundances of two potentially pathogenic genera: Salmonella and Yersinia. Salmonella are rod-shaped,
Gram-negative bacteria that can cause gastroenteritis in both rodents and humans upon infection66. Similarly,
Yersinia (including Y. pestis, Y. pseudotuberculosis, and Y. enterocolitica) are commonly harbored in the gut
microbiota of wild rodents and can lead to enteric and systemic d isease54,67, though we note there is currently
no evidence of Yersinia disease in our population. While a loss of these taxa with increasing GCs contradicts
theoretical expectations of pathogen susceptibility as microbial diversity decreases, our findings align with a
prior study in free-living birds in which elevated GCs similarly reduced the relative abundance of intestinal
pathogens24, and in piglets in which GCs reduced Salmonella, specifically68. These data suggest that elevations
in GCs may confer short-term protection against intestinal pathogens, potentially through transient increases in
gut immune function or butyrate production. Confirmation of the pathogenicity of these taxa, however, requires
high-resolution, strain-level genomic data outside the scope of the present study.
To disentangle the effects of ecological and host factors on the relationship between GCs and the gut microbi-
ome, we constructed a structural equation model (SEM) based on a set of causal a priori hypothesized pathways
(Fig. S1)43,69. As predicted, an upcoming spruce mast had a direct positive effect on gut microbiome alpha diver-
sity, and this path was the strongest path in the SEM (Fig. 4). An increase in gut microbiome diversity in the
mast year compared to the non-mast years aligns with our expectation of territorial breakdown and increased
social interactions due to an extended breeding season in the months leading up to a masting e vent36,37. Given
that the positive link between sociality and gut microbiome diversity is well-supported at least in some taxa8,29,70,
squirrels may exhibit increased gut microbiome diversity due to greater horizontal transmission of microbes
between conspecifics as social interactions become more frequent.
Similar to the effects of an upcoming spruce mast, dietary heterogeneity had a direct positive effect on
microbial diversity, though this effect was approximately 2.5× weaker than that of the upcoming mast (stand-
ardized β = 0.26). Gut microbiome alpha diversity was greatest in the months in which the food available to red
squirrels was most heterogeneous (e.g., fungi, buds, and seeds)34. Indeed, a varied diet is expected to increase
microbiome diversity through greater substrate selection for diverse ecological niches71. This effect of dietary
heterogeneity on gut microbiome diversity, coupled with prior work on the relationship between diet and gut
microbiome composition in this p opulation52, indicates that the red squirrel gut microbiome responds rapidly
to shifts in food availability.
By contrast, conspecific density indirectly, but not directly, impacted gut microbiome diversity by way of GCs,
lending support to previous findings that the frequency of social interactions (and thus horizontal transmission)
is not related to squirrel density in this population40. In line with our expectations, elevated conspecific densities
predicted increased host production of G Cs28, which in turn predicted reduced gut microbiome diversity (Fig. 4).
Red squirrels are highly sensitive to changes in density, and signals of both actual and perceived elevated den-
sity lead to GC increases independent of other ecological factors that covary with it (e.g., food)28. That elevated
density reduced gut microbiome diversity via increasing GCs aligns with our understanding of the regulation of
the gut–brain axis by the social environment, stress, and psychological state in laboratory rodents72. In the wild,
vocalizations can buffer individuals from physical interactions with conspecifics even in times of high densities
in highly territorial animals like red squirrels. Our results suggest that the indirect effects of increased density on
gut microbiome diversity likely reflect the psychosocial stress of increased competition, demonstrating a novel
link between social stress and the gut–brain axis in a wild mammal.
Reproductive state was the only host factor to exhibit an effect on gut microbiome diversity, and the effect
was indirect and the strongest path to GCs in the model. As predicted, being reproductively active (e.g., scrotal
for males, and breeding, gestating, or lactating for females) predicted greater GCs, and in turn a reduction in
microbial diversity. In both males and females, reproduction increases host metabolic demands b roadly50and,
in females in our population, GCs s pecifically49. A consequent reduction in microbiome diversity may therefore
better support host energy balance by increasing the relative abundance of core microbiota at the expense of rare
taxa that contribute less to the metabolic functions of the community. Contrary to our predictions, we found
no effect of sex or age on microbial diversity, and the SEM did not identify an effect of either on GCs via tests of
directed separation69. Studies in humans and other mammals have found mixed effects of age on both GCs and
microbial diversity48,73,74. Sex effects on microbiome composition related to hormones and behavior have been
documented in experimental rodent m odels47,75, but studies in wild populations have not typically found sex
differences in gut microbiome c omposition76.
A number of potential pathways may explain how host GCs impact gut microbiota. First, GCs can alter lipid
metabolism, leading to lipid accumulation in the gut77. Increased lipid metabolism reduces taxonomic diversity
in the gut microbiome of laboratory rodents as some bacteria exhibit sensitivity to lipid accumulation78. Sec-
ond, elevated GCs can disrupt host circadian rhythms in laboratory s ettings78, reducing gut microbiome alpha
diversity79. Finally, increased GCs can decrease mucin synthesis, which is integral to the stability of the gut
microbiome and largely determines its c omposition80. Impaired mucin synthesis as a result of elevated GCs may
therefore disrupt gut microbiome stability by reducing the resilience of the mucosal layer, leading to a reduction
in non-core bacteria and an overall loss of community diversity81.
Of note is the bidirectionality of the gut–brain axis demonstrated in laboratory rodent s tudies82,83, and thus
the potential bidirectionality of the GC-microbiome relationship in wild animals. Here, we focused on the unidi-
rectional effects of GCs on gut microbiome diversity similar to prior studies (Table 1). However, gut microbiota
can themselves regulate the hypothalamic–pituitary–adrenal axis, such that shifts in gut microbiome composition
may directly modulate host production of G Cs84 and contribute to a feedback loop between the two s ystems85.
Statistical constraints inherent to structural equation modeling prevented us from incorporating a bidirectional
relationship between GCs and microbial diversity into this s tudy69. Nonetheless, the bidirectionality of the
gut–brain axis has important implications for the evolution of the relationship between GCs and microbial
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 7
Vol.:(0123456789)www.nature.com/scientificreports/
ammals11. We encourage future research on wild populations to implement experimental
diversity in wild m
frameworks whenever possible to better characterize the complexity of the GC-gut microbiome relationship.
Methods
Ethics statement. All methods were carried out in accordance with relevant guidelines and regulations. All
research methods and protocols were conducted under animal ethics approvals from Michigan State University
(AUF#04/08-046-00), University of Guelph (AUP#09R006), and University of Michigan (PRO00005866). All
authors complied with the ARRIVE guidelines.
Study population. Subjects for this study were wild North American red squirrels (Tamiasciurus hudsoni-
cus) inhabiting a natural environment in southwest Yukon, Canada (61° N, 138° W). All subjects were continu-
ously monitored as part of the Kluane Red Squirrel Project, a long-term field study that has been conducting
a combination of live-trapping, focal behavioral observations, sampling, and experimental manipulations in
the area since 1 98738,86. Individual squirrels are marked with small metal ear tags and unique combinations
of colored wire threaded through the ear tags. Individuals are monitored from birth to death in each year of
study, roughly from March to October, using live-trapping and behavioral observations38. Individuals included
in our study lived on one of three grids (Agnes or AG, Kloo or KL, Sulphur or SU). On AG, individuals were
supplemented with peanut butter from October to May in each year for a separate experiment focused on experi-
mentally increasing squirrel population d ensity28,38. On KL and SU, no food supplementation was provided. All
models controlled for food supplementation status given its potential impacts on gut microbiome composition.
Sampling. We collected 227 samples from 88 individuals across three years (2008–2010). When individu-
als were captured, they were handled such that their unique identity could be determined (by reading ear tags),
sexed, and their reproductive condition could be recorded. Fecal samples were collected opportunistically dur-
ing live-trapping from underneath the traps using forceps. Following capture and handling of squirrels, fresh
fecal samples were collected from underneath the traps, kept on wet ice until they could be frozen at − 20 °C
within 5 h of collection in the field. Samples contaminated with urine were not collected and all samples were
kept at − 20 °C until analysis. We removed one fecal pellet from each sample using sterilized forceps for micro-
biome sequencing and then used the rest of the sample to measure fecal GC metabolites.
Age. The age of each squirrel was known as individual squirrels were uniquely tagged in their natal nest when
they were ~ 25 days of age and age is accordingly recorded at each trapping event38.
Reproductive activity. Squirrels were live-trapped regularly and handled using visualization to determine
reproduction state. During each trapping event simultaneous with the collection of microbiome and hormone
samples, the reproductive state of the individual was determined via abdominal p alpation86. Males were consid-
ered reproductively active if their testes were scrotal, and not reproductively active if testes were abdominal. For
females, pregnancy status was assessed via abdominal palpation for fetuses as well as by examining nipple condi-
tion. Females were determined to be reproductively active if they were gestating, lactating, or breeding based on
nipple condition. We have previously found that females that are reproductively active (pregnant or lactating)
have higher fecal GC metabolites than those that were not reproductively active whereas there were no differ-
ales49.
ences in the effects of reproductive activity (presence or absence of scrotal testes) in m
Dietary heterogeneity. Although red squirrels consume primarily white spruce seeds, they also consume
a number of other foods (e.g., spruce bark and needles, willow leaves and buds, fungi, and bearberry flowers) and
thus experience varying levels of seasonal dietary heterogeneity34. We coded dietary heterogeneity by ranking
the availability of these different foods across seasons from greatest (3) to least (1). Samples collected prior to
June of each year were ranked as 1, while samples collected in the month of June and late summer (July–August)
were ranked as 2 and 3, respectively.
Conspecific density. Densities (expressed as squirrels per hectare) for each grid of the study (KL, SU, AG)
were calculated separately for each year (2008, 2009, 2010) across the dataset using census data. In May of each
year, we determined the number of squirrels owning a territory on our study areas using a combination of live-
trapping and behavioral observations. Because squirrels are diurnal, regularly exhibit territorial calls, and their
territories are visually conspicuous, we were able to completely enumerate all squirrels living in our study areas.
Sequencing and bioinformatics. Microbiome data used in this study are a subset of previously pub-
lished data52. DNA extraction and sequencing was performed as described in Ren et al.52. Briefly, the V1–V3
hypervariable region of the 16S rRNA bacterial gene was amplified using two universal primers: 27F (5′-ARG
GTTTGATCMTGGCTCAG-3′) and 534R (5′-TTACCGCGGCTGCTGGCAC-3′). Samples were barcoded for
PCR amplification, pooled, gel purified, and then sequenced on an Illumina MiSeq using 300 bp paired-end
sequences. Sequences were then filtered, quality controlled, and reads were successfully merged using QIIME87.
Chimeras were removed using USEARCH88 and sequences determined to be non-chimeric by both de novo
and reference-based algorithms were retained. Reads were clustered to OTUs using U CLUST89 with an identity
threshold of 97% (genus-level). Mitochondria and chloroplast were removed, and samples were rarefied to 4000
reads per sample.
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 8
Vol:.(1234567890)www.nature.com/scientificreports/
Hormone metabolite analysis. The time period from collection in the field to freezing (~ 5 h) did not
impact fecal GC metabolites49. We measured fecal GC metabolites using previously validated protocols49,90.
Briefly, samples were lyophilized for 14–16 h, bathed in liquid nitrogen, and pulverized using a mortar and
pestle. A subsample (0.05 g) was then extracted using 80% methanol where the samples were vortexed at 1450
RPM for 30 min followed by centrifuging for 15 min at 2500g49. The supernatant was then used in an enzyme-
immunoassay that employed an antibody that measures GC metabolites with a 5α-3β,11β-diol s tructure91. We
have previously validated this assay and shown that the antibody can accurately measure increases in adrenal
production of GCs49. We have also shown that our measures of fecal GC metabolites are comparable across
assays92. Using pooled samples that were run repeatedly on different plates (n = 115) in our laboratory show that
the estimates of optical density for these pooled samples were highly repeatable (R = 0.851, 95% CI 0.543–0.925).
Using a linear mixed-effects model, we partitioned the variance in the optical density recorded for the pooled
samples that were run across these different plates and found that most of the variance was due to the sample
itself (85.1%) with little of it being explained by intra-assay variation as all samples were run in duplicate (4.9%)
or by inter-assay variation (9.9%).
Statistical analysis. All statistical analyses were conducted in R (v. 3.5.2.) (R. Core Team, 2015). OTU and
taxonomy tables were imported into R and merged into a phyloseq object for downstream analyses using the
ape93 and p
hyloseq94 packages. All figures were created in R, with the exception of the conceptual model (Fig. 1)
and the structural equation model figures (Figs. 4 and Fig. S1), which were created in bioRender (www.biore
nder.com).
Alpha diversity. The estimate_richness() function in the phyloseq package was used to calculate the
observed richness (Chao1) and Shannon Index of alpha diversity. Faith’s Phylogenetic Distances were calculated
using the pd() function on the phyloseq object in picante95. Linear mixed-effects models were used to assess the
relationship between bacterial diversity and fecal glucocorticoid metabolites (GCs), including individual ID as
a random intercept, and collection date and food supplementation as fixed effects. GCs concentrations were
log-transformed to improve model fit. Shannon Indices were Tukey transformed prior to analysis to achieve
residual normality. All models were assessed for multicollinearity among predictor variables by calculating vari-
ance inflation factors (VIF < 5).
Differential abundance testing. To identify the bacterial taxa whose relative abundances were signifi-
cantly associated with changes in host GCs, we constructed negative binomial mixed models and implemented
our analysis using the NBZIMM p ackage96. Negative binomial models outperform other traditional differential
abundance methods (e.g. DESeq) because they are better equipped to handle the zero-inflation and sparsity
common to microbiome count d ata96. Taxa included in differential abundance testing were filtered with a liberal
threshold of > 0.001% relative abundance to the overall microbiome community to avoid excluding rare taxa as
they contribute substantially to measures of community d iversity58,97. Models included the read count of each
bacterial taxa as the dependent variable, GCs (scaled to zero mean and unit variance) as a fixed effect, control-
ling for collection date (fixed), food supplementation (fixed), and individual id (random). Taxa whose negative
binomial models did not converge due to a high presence of zeroes were modeled instead with zero-inflated
negative binomial models using the glmer.zinb() function in the same package (NBZIMM). We controlled the
false discovery rate by applying a Benjamini–Hochberg FDR correction to all p-values. Adjusted p-values < 0.05
were considered statistically significant.
Structural equation modelling. To integrate ecological and host variables into our model framework
investigating the relationship between GCs and gut microbiome diversity, we constructed a structural equation
model using (SEM) using the piecewiseSEM package69. SEM is an effective way to evaluate direct and indirect
effects of multiple variables within complex ecological systems43. Unlike traditional variance covariance-based
SEM, piecewise SEM approaches allow for the inclusion of random effects, the construction of a single causal
network from multiple separate models, and the ability to handle small sample sizes and compare models using
Akaike information criterion (AIC)69.
Using piecewiseSEM, we investigated whether the relationship between GCs and gut microbiome diversity
(endogenous variables, i.e., variables of interest) was moderated by host and/or ecological factors (exogenous
variables, i.e., variance outside of the model structure). All categorical variables were converted to numeric vari-
ables prior to modeling. To build the SEM, we first constructed two component linear mixed-effect models. The
first model tested the effects of conspecific density, reproductive activity, dietary heterogeneity, and an upcoming
mast on GCs. The second model tested the effects of reproductive activity, dietary heterogeneity, an upcoming
mast, age, and GCs on gut microbiome diversity (Chao1 richness). Both component models included sample
collection date and food supplementation status as a fixed effect and individual ID as a random effect. However,
food supplementation did not affect GCs or gut microbiome diversity in either of the component models (effect
on GCs: β ± SE − 4.86 ± 3.08, t = − 1.58, P = 0.12; effect on gut microbiome alpha diversity: β ± SE − 29.14 ± 79.13,
t = − 0.37, P = 0.71), and was therefore removed from the SEM to improve model fit (AICc) and refine the stand-
ardized beta estimates. The overall fit of the SEM was evaluated using Shipley’s test of d-separation Fisher’s C
statistic and AICc.
Data availability
All sequences, hormone data, and R code related to this manuscript are available at figshare (10.6084/
m9.figshare.19077773 and https://figshare.com/s/a52886d8016cdd1f0dbb).
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 9
Vol.:(0123456789)www.nature.com/scientificreports/
Received: 23 August 2021; Accepted: 24 January 2022
References
1. Ley, R. E. et al. Evolution of mammals and their gut microbes. Science 320, 1647–1651 (2008).
2. Thaiss, C. A., Zmora, N., Levy, M. & Elinav, E. The microbiome and innate immunity. Nature 535, 65–74 (2016).
3. Ezenwa, V. O., Gerardo, N. M., Inouye, D. W., Medina, M. & Xavier, J. B. Microbiology. Animal behavior and the microbiome.
Science 338, 198–199 (2012).
4. Sampson, T. R. & Mazmanian, S. K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe.
17, 565–576 (2015).
5. Voigt, R. M., Forsyth, C. B., Green, S. J., Engen, P. A. & Keshavarzian, A. Circadian rhythm and the gut microbiome. Int. Rev.
Neurobiol. 131, 193–205 (2016).
6. Backhed, F. Programming of host metabolism by the gut microbiota. Endocr. Abstr. https://doi.org/10.1530/endoabs.32.s20.2
(2013).
7. Mallott, E. K., Borries, C., Koenig, A., Amato, K. R. & Lu, A. Reproductive hormones mediate changes in the gut microbiome
during pregnancy and lactation in Phayre’s leaf monkeys. Sci. Rep. 10, 9961 (2020).
8. Miller, E. A., Livermore, J. A., Alberts, S. C., Tung, J. & Archie, E. A. Ovarian cycling and reproductive state shape the vaginal
microbiota in wild baboons. Microbiome. 5, 8 (2017).
9. Gomez-Arango, L. F. et al. Connections between the gut microbiome and metabolic hormones in early pregnancy in overweight
and obese women. Diabetes 65, 2214–2223 (2016).
10. Shin, J.-H. et al. Serum level of sex steroid hormone is associated with diversity and profiles of human gut microbiome. Res.
Microbiol. 170, 192–201 (2019).
11. Burokas, A., Moloney, R. D., Dinan, T. G. & Cryan, J. F. Microbiota regulation of the mammalian gut–brain axis. Adv. Appl. Micro-
biol. 91, 1–62. (2015).
12. Sudo, N. The hypothalamic–pituitary–adrenal axis and gut microbiota. Gut–Brain Axis. https://d oi.o
rg/1 0.1 016/b
978-0-1 2-8 02304-
4.00013-x (2016).
13. Sapolsky, R. M., Romero, L. M. & Munck, A. U. How do glucocorticoids influence stress responses? Integrating permissive, sup-
pressive, stimulatory, and preparative actions. Endocr. Rev. 21, 55–89 (2000).
14. Hau, M., Casagrande, S., Ouyang, J. Q. & Baugh, A. T. Glucocorticoid-mediated phenotypes in vertebrates: Multilevel variation
and evolution. Adv. Stud. Behav. 48, 41–115 (2016).
15. Sprague, R. S. & Breuner, C. W. Timing of fledging is influenced by glucocorticoid physiology in Laysan Albatross chicks. Horm.
Behav. 58, 297–305 (2010).
16. Fletcher, Q. E., Dantzer, B. & Boonstra, R. The impact of reproduction on the stress axis of free-living male northern red backed
voles (Myodes rutilus). Gen. Comp. Endocrinol. 224, 136–147 (2015).
17. Romero, L. M. & Wikelski, M. Corticosterone levels predict survival probabilities of Galapagos marine iguanas during El Nino
events. Proc. Natl. Acad. Sci. USA. 98, 7366–7370 (2001).
18. Chevalier, C. et al. Gut microbiota orchestrates energy homeostasis during cold. Cell 163, 1360–1374 (2015).
19. Amato, K. R. et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 7,
1344–1353 (2013).
20. Baniel, A. et al. Seasonal shifts in the gut microbiome indicate plastic responses to diet in wild geladas. Microbiome. 9, 26 (2021).
21. Kohl, K. D., Amaya, J., Passement, C. A., Dearing, M. D. & McCue, M. D. Unique and shared responses of the gut microbiota to
prolonged fasting: A comparative study across five classes of vertebrate hosts. FEMS Microbiol. Ecol. 90, 883–894 (2014).
22. Stecher, B. et al. Like will to like: Abundances of closely related species can predict susceptibility to intestinal colonization by
pathogenic and commensal bacteria. PLoS Pathog. https://doi.org/10.1371/journal.ppat.1000711 (2010).
23. Das, B. & Nair, G. B. Homeostasis and dysbiosis of the gut microbiome in health and disease. J. Biosci. 44(5), 1–8 (2019). https://
www.ncbi.nlm.nih.gov/pubmed/31719226.
24. Noguera, J. C., Aira, M., Pérez-Losada, M., Domínguez, J. & Velando, A. Glucocorticoids modulate gastrointestinal microbiome
in a wild bird. R. Soc. Open Sci. https://doi.org/10.1098/rsos.171743 (2018).
25. UrenWebster, T. M., Rodriguez-Barreto, D., Consuegra, S. & GarciadeLeaniz, C. Cortisol-related signatures of stress in the fish
microbiome. Front. Microbiol. 11, 1621 (2020).
26. Stothart, M. R., Palme, R. & Newman, A. E. M. It’s what’s on the inside that counts: stress physiology and the bacterial microbiome
of a wild urban mammal. Proc. Biol. Sci. 286, 20192111 (2019).
27. Vlčková, K. et al. Impact of stress on the gut microbiome of free-ranging western lowland gorillas. Microbiology 164, 40–44 (2018).
28. Dantzer, B. et al. Density triggers maternal hormones that increase adaptive offspring growth in a wild mammal. Science 340,
1215–1217 (2013).
29. Sarkar, A. et al. Microbial transmission in animal social networks and the social microbiome. Nat. Ecol. Evol. 4, 1020–1035 (2020).
30. Kruuk, L. E. B., Merilä, J. & Sheldon, B. C. When environmental variation short-circuits natural selection. Trends Ecol. Evol. 18,
207–209 (2003).
31. Stinchcombe, J. R. et al. Testing for environmentally induced bias in phenotypic estimates of natural selection: Theory and practice.
Am. Nat. 160, 511–523 (2002).
32. Rausher, M. D. The measurement of selection on quantitative traits: Biases due to environmental covariances between traits and
fitness. Evolution 46, 616–626 (1992).
33. Lamontagne, J. M. & Boutin, S. Local-scale synchrony and variability in mast seed production patterns of Picea glauca. J. Ecol.
https://doi.org/10.1111/j.1365-2745.2007.01266.x (2007).
34. Fletcher, Q. E. et al. Reproductive timing and reliance on hoarded capital resources by lactating red squirrels. Oecologia https://
doi.org/10.1007/s00442-013-2699-3 (2013).
35. Fletcher, Q. E. et al. The functional response of a hoarding seed predator to mast seeding. Ecology 91, 2673–2683 (2010).
36. Boutin, S. et al. Anticipatory reproduction and population growth in seed predators. Science 314, 1928–1930 (2006).
37. Haines, J. A. et al. Sexually selected infanticide by male red squirrels in advance of a mast year. Ecology 99, 1242–1244 (2018).
38. Dantzer, B., McAdam, A. G., Humphries, M. M., Lane, J. E. & Boutin, S. Decoupling the effects of food and density on life-history
plasticity of wild animals using field experiments: Insights from the steward who sits in the shadow of its tail, the North American
red squirrel. J. Anim. Ecol. 89, 2397–2414 (2020).
39. Hestbeck, J. B. A Mathematical Model of Population Regulation in Cyclic Mammals. Population Biology 290–297 (Springer, 1983).
40. Dantzer, B., Boutin, S., Humphries, M. M. & McAdam, A. G. Behavioral responses of territorial red squirrels to natural and
experimental variation in population density. Behav. Ecol. Sociobiol. 66, 865–878 (2012).
41. Siracusa, E. et al. Familiarity with neighbours affects intrusion risk in territorial red squirrels. Anim. Behav. 133, 11–20 (2017).
42. Guindre-Parker, S. et al. Individual variation in phenotypic plasticity of the stress axis. Biol. Lett. 15, 20190260 (2019).
43. Laughlin, D. & Grace, J. Discoveries and novel insights in ecology using structural equation modeling. Ideas Ecol. Evol. https://
doi.org/10.24908/iee.2019.12.5.c (2019).
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 10
Vol:.(1234567890)www.nature.com/scientificreports/
44. Pugesek, B. H., Tomer, A. & von Eye, A. Structural Equation Modeling: Applications in Ecological and Evolutionary Biology (Cam-
bridge University Press, 2003).
45. Pearl, J. The causal foundations of structural equation modeling. (2012). https://doi.org/10.21236/ada557445
46. Leftwich, P. T., Clarke, N. V. E., Hutchings, M. I. & Chapman, T. Gut microbiomes and reproductive isolation in Drosophila. Proc.
Natl. Acad. Sci. USA. 114, 12767–12772 (2017).
47. Markle, J. G. M. et al. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 339,
1084–1088 (2013).
48. Reveles, K. R., Patel, S., Forney, L. & Ross, C. N. Age-related changes in the marmoset gut microbiome. Am. J. Primatol. https://
doi.org/10.1002/ajp.22960 (2019).
49. Dantzer, B. et al. Fecal cortisol metabolite levels in free-ranging North American red squirrels: Assay validation and the effects of
reproductive condition. Gen. Comp. Endocrinol. 167, 279–286 (2010).
50. Fletcher, Q. E. et al. Seasonal stage differences overwhelm environmental and individual factors as determinants of energy expendi-
ture in free-ranging red squirrels. Funct. Ecol. https://doi.org/10.1111/j.1365-2435.2012.01975.x (2012).
51. Lane, J. E., Boutin, S., Gunn, M. R., Slate, J. & Coltman, D. W. Female multiple mating and paternity in free-ranging North American
red squirrels. Anim. Behav. 75, 1927–1937 (2008).
52. Ren, T. et al. Seasonal, spatial, and maternal effects on gut microbiome in wild red squirrels. Microbiome. 5, 163 (2017).
53. Backhans, A., Fellström, C. & Lambertz, S. T. Occurrence of pathogenic Yersinia enterocolitica and Yersinia pseudotuberculosis
in small wild rodents. Epidemiol. Infect. 139, 1230–1238 (2011).
54. Bižanov, G. & Dobrokhotova, N. D. Experimental infection of ground squirrels (Citellus pygmaeus Pallas) with Yersinia pestis
during hibernation. J. Infect. 54, 198–203 (2007).
55. Stothart, M. R. et al. Stress and the microbiome: Linking glucocorticoids to bacterial community dynamics in wild red squirrels.
Biol. Lett. 12, 20150875 (2016).
56. Costello, E. K., Stagaman, K., Dethlefsen, L., Bohannan, B. J. M. & Relman, D. A. The application of ecological theory toward an
understanding of the human microbiome. Science 336, 1255–1262 (2012).
57. Rocca, J. D., Simonin, M., Bernhardt, E. S., Washburne, A. D. & Wright, J. P. Rare microbial taxa emerge when communities collide:
Freshwater and marine microbiome responses to experimental mixing. Ecology https://doi.org/10.1002/ecy.2956 (2020).
58. Shade, A. et al. Conditionally rare taxa disproportionately contribute to temporal changes in microbial diversity. MBio https://doi.
org/10.1128/mbio.01371-14 (2014).
59. Dinan, T. G. & Cryan, J. F. The microbiome–gut–brain axis in health and disease. Gastroenterol. Clin. N. Am. 46, 77–89 (2017).
60. Claus, S. P. et al. Colonization-induced host–gut microbial metabolic interaction. MBio 2, e00271-e310 (2011).
61. Bangsgaard Bendtsen, K. M. et al. Gut microbiota composition is correlated to grid floor induced stress and behavior in the BALB/c
mouse. PLoS ONE 7, e46231 (2012).
62. Amato, K. R. et al. The gut microbiota appears to compensate for seasonal diet variation in the wild black howler monkey (Alouatta
pigra). Microb. Ecol. 69, 434–443 (2015).
63. McLaren, M. R. & Callahan, B. J. Pathogen resistance may be the principal evolutionary advantage provided by the microbiome.
Philos. Trans. R Soc. Lond. B Biol. Sci. 375, 20190592 (2020).
64. Donohoe, D. R. et al. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell
Metab. 13, 517–526 (2011).
65. Rivera-Chávez, F. et al. Depletion of butyrate-producing clostridia from the gut microbiota drives an aerobic luminal expansion
of salmonella. Cell Host Microbe. 19, 443–454 (2016).
66. Meerburg, B. G. & Kijlstra, A. Role of rodents in transmission of Salmonella and Campylobacter. J. Sci. Food Agric. https://doi.
org/10.1002/jsfa.3004 (2007).
67. Jalal, M. S. et al. Antibiotic resistant zoonotic bacteria in Irrawaddy squirrel (Callosciurus pygerythrus). Vet. Med. Sci. 5, 260–268
(2019).
68. Petrosus, E., Silva, E. B., Lay, D. Jr. & Eicher, S. D. Effects of orally administered cortisol and norepinephrine on weanling piglet
gut microbial populations and Salmonella passage. J. Anim. Sci. 96, 4543–4551 (2018).
69. Lefcheck, J. S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol.
Evol. https://doi.org/10.1111/2041-210X.12512 (2016).
70. Raulo, A. et al. Social networks strongly predict the gut microbiota of wild mice. ISME J. https://doi.org/10.1038/s41396-021-
00949-3 (2021).
71. Phillips, C. D. et al. Microbiome structural and functional interactions across host dietary niche space. Integr. Comp. Biol. 57,
743–755 (2017).
72. Galley, J. D. et al. Exposure to a social stressor disrupts the community structure of the colonic mucosa-associated microbiota.
BMC Microbiol. 14, 189 (2014).
73. Wu, C.-S. et al. Age-dependent remodeling of gut microbiome and host serum metabolome in mice. Aging 13, 6330–6345 (2021).
74. Altmann, J., Gesquiere, L., Galbany, J., Onyango, P. O. & Alberts, S. C. Life history context of reproductive aging in a wild primate
model. Ann. NY Acad. Sci. https://doi.org/10.1111/j.1749-6632.2010.05531.x (2010).
75. Sylvia, K. E., Jewell, C. P., Rendon, N. M., St John, E. A. & Demas, G. E. Sex-specific modulation of the gut microbiome and behavior
in Siberian hamsters. Brain Behav. Immun. 60, 51–62 (2017).
76. Grieneisen, L. E., Livermore, J., Alberts, S., Tung, J. & Archie, E. A. Group living and male dispersal predict the core gut microbiome
in wild baboons. Integr. Comp. Biol. 57, 770–785 (2017).
77. Peckett, A. J., Wright, D. C. & Riddell, M. C. The effects of glucocorticoids on adipose tissue lipid metabolism. Metabolism 60,
1500–1510 (2011).
78. Wu, T. et al. Chronic glucocorticoid treatment induced circadian clock disorder leads to lipid metabolism and gut microbiota
alterations in rats. Life Sci. 192, 173–182 (2018).
79. Deaver, J. A., Eum, S. Y. & Toborek, M. Circadian disruption changes gut microbiome taxa and functional gene composition. Front.
Microbiol. 9, 737 (2018).
80. Schroeder, B. O. Fight them or feed them: How the intestinal mucus layer manages the gut microbiota. Gastroenterol. Rep. 7, 3–12
(2019).
81. Huang, E. Y. et al. Using corticosteroids to reshape the gut microbiome: Implications for inflammatory bowel diseases. Inflamm.
Bowel Dis. 21, 963–972 (2015).
82. Carabotti, M., Scirocco, A., Maselli, M. A. & Severi, C. The gut–brain axis: Interactions between enteric microbiota, central and
enteric nervous systems. Ann. Gastroenterol. Hepatol. 28, 203–209 (2015).
83. de Weerth, C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neurosci. Biobehav.
Rev. 83, 458–471 (2017).
84. Luo, Y. et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl.
Psychiatry. 8, 187 (2018).
85. Cryan, J. F. et al. The microbiota–gut–brain axis. Physiol. Rev. 99, 1877–2013 (2019).
86. McAdam, A. G., Boutin, S., Sykes, A. K. & Humphries, M. M. Life histories of female red squirrels and their contributions to
population growth and lifetime fitness. Ecoscience 14, 362–369 (2007).
87. Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 7, 335–336 (2010).
Scientific Reports | (2022) 12:2605 | https://doi.org/10.1038/s41598-022-06359-5 11
Vol.:(0123456789)You can also read

















































