Evolution Autoencodes Life's Interactions as Species that are Decoded into Ecosystems - arXiv
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Cohen and Marron, March 2022
Evolution Autoencodes Life’s Interactions as
Species that are Decoded into Ecosystems.
Irun R. Cohen∗1 and Assaf Marron2
1
Department of Immunology and Regenerative Biology, Weizmann Institute of Science,
Rehovot , 76100 Israel irun.cohen@weizmann.ac.il
2
Department of Computer Science and Applied Mathematics, Weizmann Institute of Science,
Rehovot, 76100 Israel , assaf.marron@weizmann.ac.il
Abstract The continuity of life and its evolution, we have proposed,
emerge from an interactive group process termed Survival-of-the-Fitted.
This process supplants the Darwinian theory of individual struggle and
arXiv:2203.11891v4 [cs.NE] 30 Jun 2022
Survival-of-the-Fittest as the primary mechanism of evolution. Here,
we propose that Survival-of-the-Fitted results from a natural process
functionally related to computer autoencoding. Autoencoding is a
machine-learning technique for extracting a compact representation of
the essential features of input data; dimensionality reduction by autoen-
coding establishes a code that enables a variety of applications based on
decoding of the relevant data.
We establish the following points:
1) We define a species by its species interaction code, which consists
of the fundamental, core interactions of the species with its external
and internal environments; core interactions are encoded by multi-scale
networks including molecules-cells-organisms.
2) Evolution proceeds by sustainable changes in species interaction
codes; these changing codes both reflect and construct the species
environment. The survival of species is computed by what we term
Natural Autoencoding: arrays of input interactions generate species
codes, which survive by decoding into networks of sustained ecosystem
interactions. DNA is only one element in Natural Autoencoding.
3) Natural Autoencoding and artificial autoencoding processes manifest
defined similarities and differences.
Survival-of-the-Fitted by Natural Autoencoding sheds a new light on the
mechanism of evolution and explains why a habitable biosphere requires
a diversity of fitted group interactions.
Keywords
Evolution, Interaction, Survival of the Fitted,
Species Interaction Code, Biosphere Autoencoding;
Artificial Autoencoding; Encoding; Decoding.
Page 1 of 15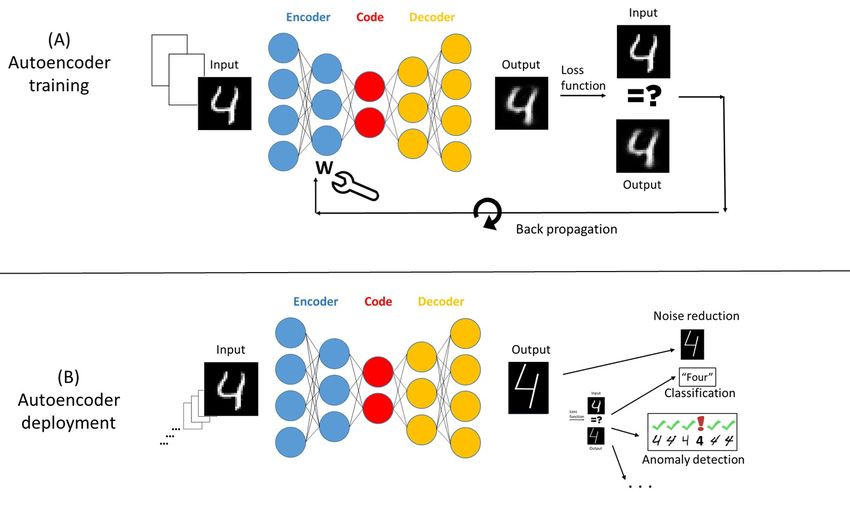
Cohen and Marron, March 2022
1 Evolution emerges from (termed interactors) in which the interactors transmit
or exchange energy, matter or information.
cooperative interactions; Sustainable types of interactions are characterized
basic definitions. by repetition and sequence. Metabolic interactions,
Evolution is the narrative of changes in species and for example, are organized in repeating, sequential
their interactions over time (Koonin, 2011). pathways—one interaction connected to the next in
Ever since Darwin (Darwin, 1860), the mechanism un- line (Judge and Dodd, 2020). Cycles of reproduction,
derlying evolution has been taught to be Natural Selec- growth, aging and death are accessible examples of the
tion, which is based on a continuous struggle of variant universality of repeated, sequential interactions.
individual organisms for survival and reproductive ad- Energy is the impetus behind motion and activ-
vantage in the face of limited resources; this struggle ity (Doige and Day, 2012), including the capacity to
leads to survival of the fittest variants and their domin- do work. Energy enables interactions.
ion over the less fit; predominating proliferation of the Information we define according to Shannon as a par-
winners determines the characters of species. ticular non-random structure or arrangement of entities
In the interval since Darwin, science has learned that or processes (Shannon, 1948; Cohen, 2006).
all living systems—cells, organisms, species (including Arrangements bear information; but an arrangement
Homo sapiens) and ecosystems—survive in extensive by itself has no meaning unless it interacts with other
networks of interaction and group cooperation (Cohen, arrangements to produce some effect (Cohen, 2006;
2016; Cohen and Marron, 2020; Sachs et al., 2004). A Cohen, 2000). The consequences of the interactions
few examples include the dependence of every multi- of structured information constitute the meaning of
cellular organism on a microbiome (Blaser, 2014); the the information. A sequence of DNA, for example,
symbiotic web of forest trees and fungi (Simard, 2018) ; bears information that only gains meaning through ex-
and the collaboration and symbiosis that create a coral pressed interactions including transcription and trans-
colony (Rosenberg et al., 2007). The biosphere is sus- lation (Cohen et al., 2016). Written words, too, have
tained by these interactions; the biosphere is a world no meaning unless somebody or some thing can read
wide web of interactions. them. Interaction extracts meaning from information.
Survival-of-the-Fitted is an alternative mechanism to Matter itself is a product of interaction: the nuclei of
account for evolution (Cohen and Marron, 2020; Co- atoms are created by interactions between fundamen-
hen, 2016; Cohen, 2000). Rather than the Darwinian tal particles; atoms are formed by interactions between
struggle for individual reproductive advantage, surviv- nuclei and electrons; and molecules are formed by in-
ing organisms and species are those that integrate into teractions between atoms.
networks of sustaining interactions; longevity and rates So we must conclude that anything made of atoms or
of reproduction are not individual achievements but are molecules, including living entities and the biosphere
largely encoded in the species. itself, is made of interactions. As stated by Feyn-
Survival-of-the-Fittest would claim that what wins, man (Gleick, 1993) and others (Rovelli, 2017), inter-
works; Survival-of-the-Fitted affirms that what works, actions constitute reality.
works.
Natural Selection embodies fitness as the mechanism
that drives evolution. What mechanism generates fit- 2 Species are formed by core
tedness? In the body of this paper, we shall demon- codes of interaction.
strate that Survival-of-the-Fitted is the outcome of a The concept of species is ancient, and species, since
process we term Natural Autoencoding. Darwin, are linked to evolution; this link is reflected in
the title of Darwin’s foundational work on evolution:
Interactions evolve The Origin of Species (Darwin, 1860).
The substance of evolution, we have proposed, is the The definition of species, despite the term’s wide use,
process of interaction (Cohen and Marron, 2020); in- is controversial: A search in Google Scholar for species
teractions are what evolve. For example, individual returns millions of publications; but there is not one
brain cells of humans and apes, indeed, of cats and universally accepted definition; scientists have proposed
mice, may be quite similar. The differences between dozens of different definitions based on morphology, ge-
mammalian species and between individuals within a netics, sexual reproduction, ecology, and other crite-
mammalian species can emerge from the different num- ria (Mallet, 1995).
bers and interaction networks of largely stereotypical The concept of species has interested humans since an-
cells (Striedter, 2005). tiquity. The Bible, for example, names species of ani-
mals that an observant person may or may not eat; the
Bible amends the list of names with a functional code:
Basic definitions a permitted species has cloven hoofs (a structural fea-
This paper uses the following terminology: ture) and chews its cud (a dynamic process) (Levit. 11,
We have already used the term interaction: Interactions Deuter. 14). Aristotle used codes, termed Vegetative,
are mutual relationships between two or more entities Sensitive, and Rational, to distinguish plants, animals
Page 2 of 15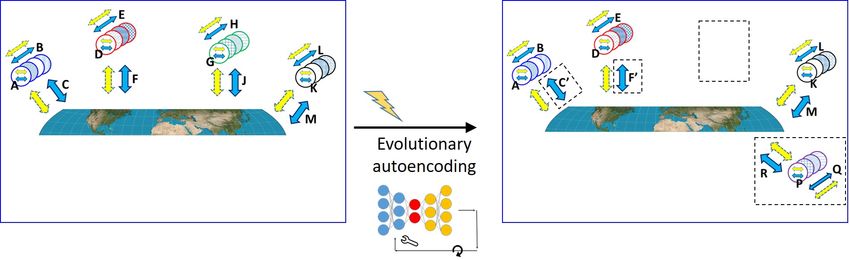
Cohen and Marron, March 2022
and humans (See Britannica entry for Aristotle). 2017); the flowers of a plant are structural and visual
People may differ about the names of species; however images of the plant’s specific pollinators (Hu et al.,
abstract, a code is objective and testable; recall Juliet’s 2008; Kritsky, 1991). Conceptually, these interactors
reference to smell in her code for defining a rose by any and their environments fit one another like locks and
name (Shakespeare’s Romeo and Juliet, Act 2). keys (Cohen, 2000).
In the spirit of codes, we here define a species as a Core codes not only fit species environments, they can
collective of organisms that share a core code of poten- actually construct them (Scott-Phillips et al., 2014):
tial, repetitive interactions with a jointly constructed beaver core interactions build dams to create beaver
external and internal environment, including both liv- environments (Westbrook et al., 2006); birds’ nests and
ing and non-living components, both structural and dy- ant hills fashion the environments of these species; ants
namic. Thus, a species interaction code of sunflowers domesticate aphids (Depa et al., 2020); interactions be-
would include interactions that are common to all or- tween wolves and humans created dog species environ-
ganisms, such as cell division, interactions that are com- ments (Nagasawa et al., 2015). Modern humans, in
mon to all plants, such as photosynthesis, and interac- contrast to now extinct versions of earlier humanoids,
tions that characterize sunflowers among others, such thrive in environments essentially constructed by mod-
as heliotropism. The core code of interactions emerges ern humans (Jablonka, 2011).
from the essential structures, processes and behaviors Barbieri has pointed out the importance of codes in
that characterize the members of a species. The core living systems generally; he proposed that life emerges
code enables the species to survive and thrive in its en- from codes that enable the maintenance and the de-
vironment. All interacting organisms express their own velopment of structures and processes, including the
species interaction codes. genetic code and its expression; on this basis, he de-
In principle, each species could be characterized by par- veloped the concept of “codepoiesis”, the idea that liv-
ticular core interactions. A detailed list of a species ing systems function to preserve organic codes and to
interaction code for even the simplest species of bacte- evolve by developing new codes. Barbieri defines a code
ria would challenge experts. We suggest that a pair- as “a mapping between the objects of two independent
wise or set perspective might provide a manageable worlds” (Barbieri, 2015; Barbieri, 2012). Species in-
solution: given two related or interacting species or teraction codes, in contrast, are not mappings between
sets of species, we might focus only on the interactions "independent worlds"; rather they are sets of mutually
that distinguish the pairs or the sets. Below, we shall dependent interactions that link organisms to their spe-
bring species of voles and crows to illustrate that re- cific environments and ecosystems.
lated species may be distinguished by only a few code An interactive system, like a species, that encodes
interactions. an internal representation of its environment can be
Living systems manifest a great variety of interactions; termed a cognitive system (Cohen, 2000). In this sense,
however, living systems exist by virtue of two essen- species are cognitive systems; each, thanks to its core
tial properties: their ability to reproduce their kind code, bears an intrinsic representation of how it lives
and their ability to metabolize the energy they require and where it lives. This representation of the species
for maintenance and reproduction in their particular environment enables a species to integrate within a suit-
environment. Quite simply, species whose constituent able ecosystem. Below, we shall describe the integra-
organisms do not reproduce and metabolize cannot tion of species into sustaining ecosystems as a decoding
live (Dupré and O’Malley, 2013). The details of these of species interaction codes. Cognition can thus be seen
essential interactions are encoded in species interaction as an element in fittedness — as a way to fit into an
codes. appropriate environment.
Matter alone, unlike living species, is not cognitive;
other than its level of energy, matter bears no intrinsic
Core codes reflect and construct the representations of how or where it exists.
species environment
In addition to describing the shared program that es-
tablishes organisms in a species, the core code ex- Core species interactions may be shared
presses an image of the environments with which the and/or specialized
constituent organisms of the species interact (Cohen, Some core interactions may be common to many dif-
2000). This image of the environment is intrinsic to ferent species: all organisms share cell division; all
the species. For example, the shedding of their broad multi-cellular organisms interact with resident micro-
leaves by deciduous trees is an image of the recur- biomes (Blaser, 2014); all mammals, except for mar-
rent winters the trees have evolved to endure (Hill supials and monotremes (Graves, 1996), interact with
and Broughton, 2009); nitrogen-fixing plants manifest their fetuses by way of placentas (Wildman et al.,
structures and molecular processes that make possible 2006), but as far as we know, only H. sapiens, among
their interactions with particular species of nitrogen- mammals, features core codes that include social and
fixing bacteria—likewise, the bacteria engage in core in- conceptual interactions using verbal language (Chris-
teractions with their host plants (Vitousek et al., 2002); tiansen and Kirby, 2003).
an animal’s teeth echo the animal’s diet (Melstrom, The core code of interactions may include differences
Page 3 of 15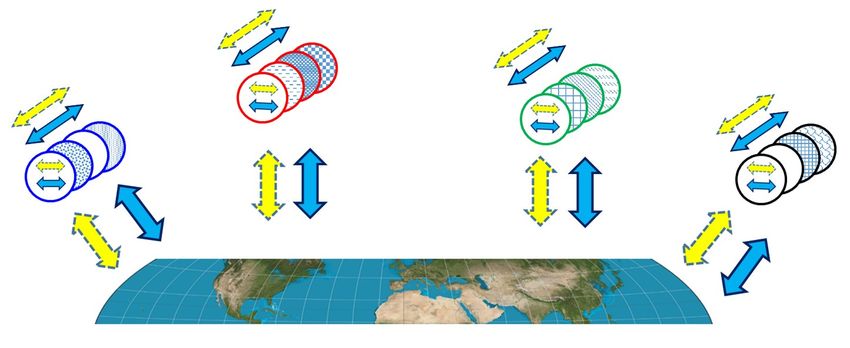
Cohen and Marron, March 2022
between individuals or classes within a species; for ex- work, or complex system, express and process more in-
ample, males and females perform different interac- formation or manifest properties not previously present
tions; only queen bees produce fertile eggs; some collec- in its constituent components (Crutchfield, 1994; Co-
tives are controlled by dominant individuals; a variety hen and Harel, 2006).
of specializations exist within particular species. Importantly, the scales of interactions in living sys-
Because of the specialization of organisms within a tems are interwoven composites, one hidden within the
species, not every organism need perform all the in- other—molecules in cells, cells in organisms, organisms
teractions in the species’ list of core interactions. in species, and species in ecosystems.
Repeated patterns of interactions, be they biochemical
Circumstantial interactions can precede or social, form additional levels, albeit more dynamical
than structural, within this multi-scale architecture.
core interactions Molecular networks, emerging from the chemical evo-
Beyond a code of core interactions, many interactions
lution of matter, evolved into a primordial type of liv-
are circumstantial; for example, most humans today
ing cell that further evolved into families of prokaryote
interact with computers, but this interaction is only
cells (Puigbò et al., 2010). Prokaryotes, then and now,
recent; the hooded crows in Rehovot interact with hu-
survive in cooperative ecosystems (Keller and Segel,
mans and cats to eat cat food and human garbage, but
1970; O’Toole et al., 2000).
only recently. These circumstantial interactions are not
Some billion years after their appearance, some
core codes, but may be derived secondarily from core
prokaryote cells (bacteria and archea) merged through
codes—for example, human interactions with comput-
endosymbiotic interactions to initiate a higher scale of
ers evolved from the core code of human language; cir-
evolution—the eukaryote cell (Margulis, 1981), which
cumstantial crow interactions with humans and cats
features complex organelles including nuclei, mitochon-
evolved from crow core social intelligence (Kurosawa
dria, and more.
et al., 2003).
Yet higher-scale multi-cellular organisms arose from the
Some core interactions are likely to arise de novo from
evolution of cell differentiation, a process that enables
a critical change, such as a mutation; for example, ge-
single eukaryote cells to specialize into variously dif-
netic changes in the human larynx and voice box, which
ferent functional types and band together by activat-
are absent in the Neanderthals (Gokhman et al., 2020),
ing selective segments of their DNA (Slack and Dale,
may have had a significant influence on the core inter-
2021); skin, muscles, bones, brains, guts and the rest of
actions mediated by human speech.
the cell types that constitute multi-cellular organisms
Other core species interactions may first have arisen
differentiate from stem cells.
as circumstantial interactions; the phenomenon of ge-
These more complex multi-cellular organisms continue
netic assimilation described by Waddington is an
to interact with earlier forms of life that may not
example (Waddington, 1953): Waddington exposed
undergo differentiation; as we mentioned above, all
Drosophila (fruit flies) to heat shock which led to a
multi-cellular organisms depend on an essential micro-
change in the structure of the insects’ wings; repeating
biome (Blaser, 2014).
the circumstantial heat shock over several generations
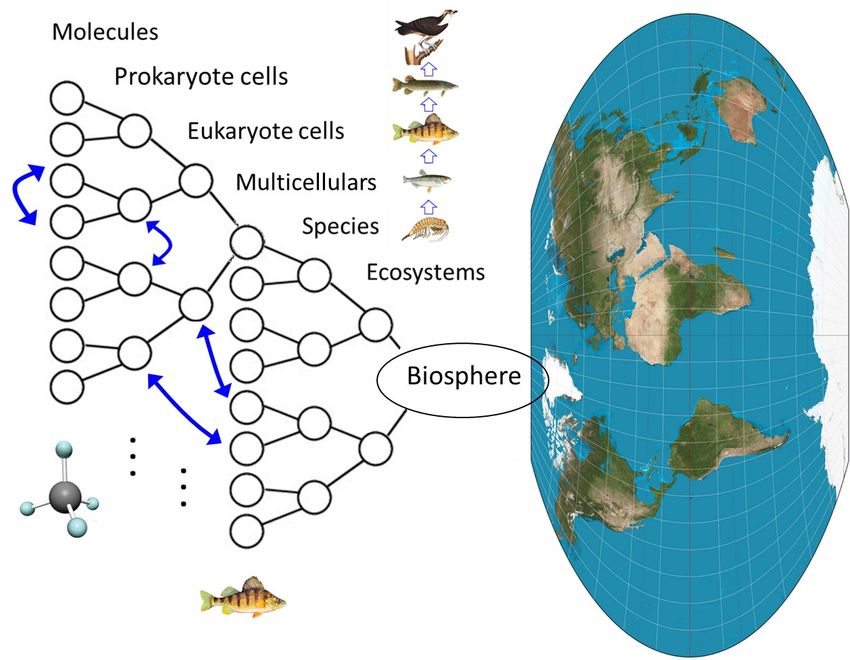
Figure 2 depicts networks of interactions both within
eventually led to genetic fixation of the variant wing
and between networks of molecules, prokaryote and eu-
form, even in the absence of circumstantial heat shock.
karyote cells, and organisms, culminating in species,
Many core genetic interactions are likely to have arisen
ecosystems and the biosphere itself. Cells and organ-
from a history of circumstances. Figure 1 schematically
isms evolved sequentially over eons of time, but all life
summarizes the structure of the biosphere manifested
is now integrated into one multi-scaled biosphere.
through species codes of interactions.
This multi-scale, interaction-based architecture and the
constraints it induces on its constituents suggested to
The evolution of species is the evolution us that the biosphere might perform a type of autoen-
of species interaction codes coding. First, we shall briefly describe computer au-
A species becomes extinct in nature when, due to a toencoding, and then we shall apply the autoencoding
lack of energy or offspring, an insufficient number of concept to the natural autoencoding of evolution.
member organisms remain to fulfill the original species
interaction code (Purvis et al., 2000). The evolution of
a new species interaction code signifies the birth of a 4 Artificial autoencoding extracts
new species. the essential features of data
sets.
3 The biosphere has evolved Autoencoding is a term associated with artificial in-
interactions across scales of telligence, machine learning, and artificial neural net-
works (Kramer, 1991)(Goodfellow et al., 2016, Chapter
complexity. 14)
Life’s interactions are scaled. We designate a higher An autoencoder is a computer program that learns
scale when the interactions of entities in a given net- defining features of the individuals in a given popu-
Page 4 of 15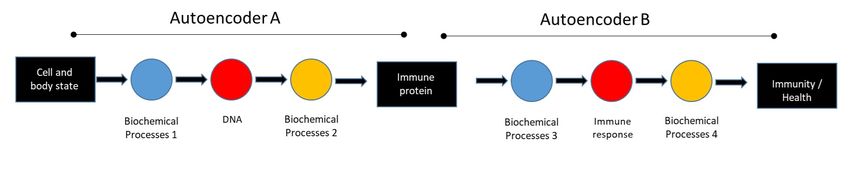
Cohen and Marron, March 2022
lation, and then represents each individual encoded as face recognition, cleaning out image data by removing
a value in a feature vector, or array. This code and its insignificant "noise", anomaly detection, classification
associated encoding, generated by autoencoding, con- and more.
stitute a compact representation of the population and Figure 3 depicts the training and deployment of a typ-
its individuals. For example, a 1000-by-1000 pixel pic- ical artificial autoencoding operation.
ture of a straight line segment, may be encoded using
just start and end coordinates, line width, and color.
Encoding and decoding occur in nature:
An autoencoder is constructed by an iterative train-
ing process that discovers the exact computations that
Music, for example
yield both a compact code and a faithful reconstruction Below, we shall apply autoencoding and encod-
of each encoded individual. ing/decoding, to biological interactions and to their
evolution; here, we convey the concepts of encoding and
The machinery of an artificial autoencoder includes
decoding from the computer to the natural world by
three elements: (1) The encoder receives as input raw
considering the example of human music (Fitch, 2006):
data about selected individuals, such as pixels of an im-
age, an audio signal, or measurement data from some A musical score is almost always the result of selec-
problem domain; the encoder outputs the learned fea- tive encoding by a composer of complex inputs present
ture vector with individual value assignments; this fea- in his or her brain of emotions, training, life experi-
ture vector constitutes (2) The code. (3) The decoder ence, innate talent, previous compositions and other
accepts the code representing the encoding of a particu- factors including constraints of time, budget and op-
lar individual, and reconstructs the original data, such portunity. The musical score is the resulting code,
as the image or the speech segment. which is decoded by the performers into actual music,
which is then encoded by listeners into their own brain
In the training process that builds the encoder and de-
state, which is then decoded into the listeners’ behav-
coder, the inputs and outputs are compared using a loss
ior, which might then be encoded into a state of family
function to determine how close the reconstructed out-
and social interactions.
puts are to the respective inputs. The internal parame-
ters of encoder and decoder, which are commonly built This chain of ongoing encoding-decoding-encoding in-
using neural networks, are then adjusted and tuned, in teractions sets the stage for appreciating the autoen-
a process termed backpropagation, usually carried out coding enterprises that mark biology and the evolution
by gradient descent, to minimize the loss function. of the biosphere.
Training an autoencoder is unsupervised: the data are
not labeled, so the autoencoder does not know what 5 The evolution of the biosphere
it is encoding. It is only required that the outputs be
very similar to the corresponding inputs. manifests autoencoding.
Once trained on a representative sample of a popula- In everyday use, the word decoding refers to the ex-
tion, the autoencoder is able to encode and faithfully traction of the meaning of some coded information. In
reconstruct many inputs from this population. Further- the realm of machine-learning with autoencoding, as
more, certain autoencoders, termed variational, can use in communications protocols and encryption, decoding
the code to generate new entities to be included in that focuses on re-creating the input.
population. Encoding and decoding in the context of the biosphere,
The array of features that comprises the numerical code in contrast, refers to the indefinite repetition of biolog-
may or may not include traits that a human observer ical interactions on planet earth. The realization that
would intuitively use to compactly describe the individ- the biosphere is sustained by continuous encoding and
ual. Hence this code vector is often referred to as the decoding and the transformation of species codes leads
latent, invisible vector, where only a properly trained to the definition of Natural Autoencoding (Section 4).
decoder can "understand" its features. Despite their differences, both the biosphere and partic-
Consequently, artificial autoencoding can function ular machine-learning computational processes extract
without its human operators assigning meaning to the and decode hidden codes in diverse populations. This
details of the process, that is, how the multitude of con- discovery is the basis of the present paper.
nection weights and non-linear functions relate to the Each molecule, cell, and organism emerges from the set
problem at hand. Autoencoding, as it were, takes place of its constituent interactions. In addition, the set of
in a "black box" in which humans choose the network interactions that the entity may perform is a program,
architecture and the activation functions, select the in- not unlike a computer program, which is described by
put, and develop the loss function. Autoencoder inter- its functions—a set of algorithms for reactive behavior.
pretability and explainability are still areas of research. Conceptually, we have separated Natural Autoencod-
The opacity of autoencoding is important for our un- ing into distinct encoding and decoding processes. The
derstanding of Natural Autoencoding of biosphere evo- interactions of living systems, however, are integrated
lution, described below; biosphere autoencoding takes into functioning composites; thus it would be difficult
place without any access of the biosphere to external to label a particular reproductive, developmental or
representations of the process. metabolic process as purely encoding or purely decod-
Artificial autoencoders enable many uses, including ing.
Page 5 of 15Cohen and Marron, March 2022
For example, a particular sequence of DNA is decoded maintains two closely related species in the same physi-
into a linear amino acid sequence, which itself is a code cal environments? It turns out that the 0.28 percent ge-
that encodes a functionally folded protein. This protein netic difference between the species encodes the degree
then serves as a code that is subsequently decoded, in and pattern of feather pigmentation (Poelstra et al.,
the case of an antibody, into an immune response, that 2014); the two species, distinguished only by their ap-
can help encode a higher scale immune reaction that pearance (carrions are totally black; hoodeds are partly
protects the organism from an infection. Indeed, a ma- grey), live in peace, defying Darwinian competition. It
chine learning process has been invoked to account for seems that crows prefer to mate with partners who look
the encoding of the state of the body by the mammalian like their parents (Metzler et al., 2020). Thus, the two
immune system (Cohen and Efroni, 2019). More gener- species are distinguished by a single interaction code
ally, every interaction, input, code, or output of a given determinant of what Darwin has termed sexual selec-
autoencoder may also serve a function in another au- tion (Darwin, 1860); what works, works.
toencoder (See Figure 5). Chained autoencoders may
also be viewed in the aggregate, forming a higher scale
autoencoding process. Constraints channel encoding and decod-
Encoding and decoding processes can be used to de- ing
scribe essentially all biological interactions that emerge Interactions in general are organized by limitations, or
from genomic DNA including metabolic networks; constraints, imposed on the interactors and on the en-
growth, development and differentiation; the activities vironment; no structures can emerge when degrees of
of the nervous, endocrine, cardiovascular, renal, respi- freedom are not limited (Grotzinger et al., 1995). In-
ratory and digestive systems; reproduction and aging, teractions result from constraints that channel the in-
and more. teractors to meet and interact. Moreover, interactions
Note that the DNA codes of living entities actually themselves generate new constraints on what may fol-
emerge from the very biological structures and inter- low; interactions constrain degrees of freedom. The cell
actions encoded in DNA; the synthesis, sequence, re- membrane, for example, is formed by interactions be-
combination, and expressions of DNA emerge from bi- tween lipids, proteins and other molecules that effec-
ological interactions that encode and decode DNA. A tively establish the boundaries of the cell.
sequence of DNA is a code only if it is decoded; a code Encoding and decoding, in this sense, are no differ-
is defined as such by the decoding interactions that de- ent from other types of interactions; each instance of
ploy it. A sequence of DNA, like a number, a speech encoding and decoding is guided and organized by its
sound, or a shape, is not a code unless it is decoded. own landscape of constraints.
Any code is realized only by its decoding.
A few genes encoding interactions can Housekeeping autoencoding maintains
distinguish species: voles and crows existing species
Voles: The species termed prairie vole (Microtus ochro- We distinguish housekeeping autoencoding from evolu-
gaster ) and meadow vole (M. pennsylvanicus) look very tionary autoencoding.
much alike, but the two species differ markedly in re- Housekeeping autoencoding refers to existing species
productive and social behaviors: prairie vole males interactions that have not been perturbed by innova-
are largely monogamous and social while meadow vole tions that change species codes. Biological "business
males are polygamous and solitary (Gruder-Adams and as usual" is housekeeping—maintaining the house.
Getz, 1985). These interaction patterns are compo- Natural housekeeping autoencoding is similar to the er-
nents of the core codes that distinguish the two species. ror correction and noise reduction applications of ar-
But meadow voles can be induced to express prairie vole tificial autoencoding in that the decoding process re-
interactions: Experimental insertion of a vasopressin stores the integrity of the input. In artificial autoen-
receptor transgene into a specific site in the ventral fore- coding such noise reduction is computed in a trained
brain of adult male meadow voles changes their repro- autoencoder using the learned weights of the neural
ductive and social behavior into those that mark prairie net (see Figure 3). In natural housekeeping, the out-
voles; they become monogamous and friendly (Lim put is generated by subjecting the input to genetic pro-
et al., 2004). The results of this experiment illustrate grams for DNA repair (Sancar et al., 2004), cancer sup-
that two existing species can be distinguished by a core pressor mechanisms (Soussi, 2000), immune reactions
code difference related to a change in the local expres- and programmed death (Cohen, 2000), among others.
sion of a single gene; what works, works. These interaction programs are included in the existing
Crows: Hooded crows (Corvus cornix ) and carrion species core interaction code. Housekeeping autoencod-
crows (C. corone) are very similar genetically (99.72 ing maintains the state of the individual by restoring a
percent identical) to the point that they can produce healthy, sustainable body state; but restoration of the
fertile hybrid offspring (Wolf et al., 2010). The two individual is not evolution.
species are in contiguity in large areas of Europe; why Evolutionary autoencoding refers to the evolution of
has not one species dominated the other? What factor new species interaction codes.
Page 6 of 15Cohen and Marron, March 2022
Evolutionary autoencoding follows inno- into ecosystems is not a strategic response to change;
vations the encoding/decoding and autoencoding processes are
Changes in species interaction codes emerge from in- the essence of the interactive existence of the biosphere.
novations that lead to novel interactions of encoding The decoding of species interaction codes into ecosys-
and decoding. Innovations are changes that are not tems is a manifestation of survival of the fitted. We
included in the existing core codes. here propose natural autoencoding of innovations as
Conceptually, there are two broad classes of innova- the underlying principle that accounts for evolution by
tions: either the environment has changed in a way Survival-of-the-Fitted (Cohen, 2016; Cohen and Mar-
that renders the core code ineffective in maintaining the ron, 2020). Extinction results from a failure to decode
species, or the core code has changed so that it is no effectively.
longer relevant to the present environment. Of course, Figure 5 depicts an aspect of the simultaneous, parallel
changes can take place both in the present code and and overlapping flow of encoding and decoding interac-
in the environment—for example, genetically variant tions among multiple living entities.
members of a hominoid species migrated out of Africa Recently, Vanchurin and colleagues have used machine
to evolve into the Neanderthal species in the European learning concepts and thermodynamic principles to de-
environment (Mellars, 2004). velop a theory of evolution (Vanchurin et al., 2022a;
Vanchurin et al., 2022b). This theory proposes that the
An innovation can enter the biosphere at any scale,
individual organism "learns" its degree of evolutionary
be it a molecular mutation, an infecting pathogen, an
fitness by a process of machine learning.
invading species, a cancer cell, a change in nutrients
Our concept of housekeeping autoencoding could be
or in solar radiation, a natural cataclysm or a social
aligned with Vanchurin and colleagues regarding the
or a technological invention; witness, for example, the
focus on the individual. But our concept of the evolu-
industrial revolution and global warming (Rosenzweig
tionary autoencoding of species interaction codes dif-
et al., 2008).
fers: Evolution of species as groups emerges from inno-
If the innovation is not integrated into a fitted configu- vations not included in an existing species code. Evo-
ration within a networked species interaction code, then lution cannot be guided by the reconstitution of the
an unfitted interaction state can emerge, which may initial species input; evolution proceeds by the integra-
negatively affect molecules, cells, organisms, species, tion of new species interaction codes.
and ecosystems; the unfitted innovation will ultimately
fail to survive and become extinct. Figure 4 shows the
effects of innovations on the evolution of the biosphere. 6 Artificial and natural autoen-
Evolutionary autoencoding differs from housekeeping
autoencoding in that the outputs of evolution are new
coding manifest significant dif-
core interaction codes. Evolutionary autoencoding also ferences.
differs from artificial autoencoding in that there is no Here, we list essential and technical differences between
restoration of the original input, no measure of a loss natural and artificial autoencoding. The comparison
function and no form of back propagation. can help clarify both mechanisms.
The transition from individual housekeeping autoen-
coding to new species interaction codes is a regular fea- 1. Internal Operation: Typical artificial autoen-
ture of the evolution of species; the serial generation coders iteratively compute and optimize a loss
of new interaction codes from existing codes and the function (Goodfellow et al., 2016, Chapter 14):
extinction of previous codes constitute the evolution of each input is encoded, its representation is then de-
life. coded, and the differences between inputs and cor-
How does the biosphere decide which species interac- responding reconstructed outputs are computed.
tions to keep and which to discard? The results and the computed gradients are used
In nature there is no species optimization step as in to drive the adjustment and learning of the autoen-
artificial autoencoding; instead, many individual inter- coder. In Natural Autoencoding, evolving species
actors are formed and many individual interactions are interaction codes stabilize around working interac-
triggered, and those that happen to repeat are retained tion patterns. The natural autoencoder works with
in the species by the fact that they work, not by a the present state without assessing how "good" it
choice based on an external criterion such as the value is, or whether it is or has been sustainable. As we
of a loss function, or on a Darwinian "optimum" such stated above, what works, works. Viewed differ-
as a reproductive advantage (Darwin, 1860). Species ently, artificial autoencoding is devised by humans
interaction codes, including species-specific life spans to perform an assigned function (Bank et al., 2020;
and reproductive rates, are decoded by the integration Goodfellow et al., 2016), and hence it can involve
of a species into its particular ecosystem arrangement. measurements of success or progress. Natural au-
Once formed, sustaining ecosystems can continue to toencoding processes are driven inevitably by the
grow, adding materials, organisms, species and energy laws of nature and the state of the biosphere; there
flows. are no preset goals, that can be used for assess-
Autoencoding of interactions into species and decoding ment.
Page 7 of 15Cohen and Marron, March 2022
A particular aspect of these operations is that of selected entities (Goodfellow et al., 2016, Section
feedback-based repair. Both artificial autoencod- 14.2.2) Biosphere autoencoding focuses on each en-
ing and natural housekeeping autoencoding can tity’s interactions with other entities.
use existing codes for repair(Goodfellow et al.,
2016, Section 14.2.2). However, natural evolu- 6. Handling deviations and innovations: Artifi-
tionary autoencoding shapes species interaction cial autoencoding is usually designed to avoid or
codes following innovations whose impact cannot trim changes, deviations and innovations; where
be learned or predicted from past experience. variations are allowed, they are constrained to a
desired distribution (Goodfellow et al., 2016, Sec-
2. Cross connectivity of scales: In typical arti- tions 3.14 and 20). Biosphere autoencoding ac-
ficial autoencoding, nodes at a particular network commodates and retains diverse innovations that
layer are connected only to nodes in the next higher happen to be sustainable; this is evolution.
layer(Goodfellow et al., 2016, Chapter 14; Open-
ing paragraph); more general connectivity can be Despite the differences listed above, artificial and natu-
found in Boltzmann Machine architecture (Ack- ral autoencoding share the defining property of an en-
ley et al., 1985), but this design does not include coding process that reduces the dimensionality of the
multiple built-in scales or hierarchies. Natural au- input to generate a code amenable to decoding. Evolu-
toencoding features interactions both between and tion, it seems, produced Natural Autoencoding millions
within scales—for example, in addition to the in- of years before humans developed artificial autoencod-
teractions between a cell and its host organism, ing. We introduced the concept of autoencoding by
a cell can interact with other cells in the same referring to a version of artificial autoencoding. How-
organism, and with other organisms as shown in ever, from the perspective of evolutionary time, arti-
Figure 2. ficial autoencoding, is a "non-conventional" variant of
the natural process that proceeded it.
3. Recursion and reflectivity: In artificial autoen-
coding, the autoencoder itself is usually not part
of the input; where recursion is applied, it is by 7 Autoencoding clarifies the dy-
iterative feeding of intermediate results into the
autoencoder (Socher et al., 2011); reflection is in
namics of evolution.
the backpropagation process, which exists in a dif- Darwin saw evolution as a gradual process; moreover,
ferent realm than the input and output data. In he believed that evolution in abrupt steps would leave
Natural Autoencoding, the biosphere incorporates an opening for creationist arguments (Darwin, 1860,
the inputs, the outputs and the very machinery of Chapter 9).
autoencoding; the autoencoder recursively receives Neo-Darwinians, too, supported gradualism. Fisher,
copies of earlier versions of itself as part of the in- for example, claimed that the probability of a mutation
put. increasing the "Darwinian fitness coefficient" of an or-
ganism decreases proportionately with the magnitude
Another aspect of this recursion is input prepa- of the mutation; Survival-of-the-Fittest assumes that
ration. Artificial autoencoding features a prelimi- any existing species must be close to its peak of fitness
nary representation, a "pre-encoding" step, to con- in a given environment; when you are already near the
vert the subject of interest into a numerical com- peak, too great a step becomes a descent. Hence, any
puter input format. The inputs into natural au- abrupt genetic change is likely to result in decreased fit-
toencoding are biosphere interactions; there is no ness (Fisher, 1958). Evolution just had to proceed only
pre-encoding. For example, artificial autoencod- in small, gradual steps (Pigliucci and Müller, 2010).
ing of a visual scene, starts with a numerical pixel- However, Gould and Eldredge introduced the term
based representation of an image. The input to punctuated equilibria to describe long periods of seem-
natural autoencoding of that scene are the direct ing stasis interrupted by bursts of new species in the
effects of light waves emanating from the real world fossil record (Gould and Eldredge, 1972). The authors
objects on biological receptors. interpreted the observation of stasis as a sign of silent
4. Selection of training input: Artificial autoen- equilibrium punctuated, as it were, by discrete periods
coding is generally unsupervised, but inputs are of jumps in evolutionary activity—or "saltations", as
most often preselected from a designated class described earlier by Goldschmidt (Goldschmidt, 1940;
within a defined domain (Bank et al., 2020, Section Dietrich, 2003).
4). The inputs into Natural Autoencoding are en- It is well documented that DNA mutations occur
tire ecosystems, with all their diverse entities and constantly and proceed at uniform rates, actually
dynamic interactions. establishing a biological clock of uniformly gradual
change (Lynch, 2010). Environments, too, constantly
5. Features of interest: Artificial autoencoding is change (Lindsey et al., 2013).
driven by optimization of the similarity of each The observation of stasis along with saltations raises
output to its corresponding input, relying on ob- a question: How can continuous genetic and environ-
servations of specific features or properties of the mental change be reconciled, on the one hand, with
Page 8 of 15Cohen and Marron, March 2022
an apparently static equilibrium of species and, on the References
other hand, with sudden changes and saltations in the Ackley, D. H., Hinton, G. E., and Sejnowski, T. J.
fossil record? (1985). A learning algorithm for boltzmann ma-
Autoencoding provides a solution to the paradox: on- chines. Cognitive science, 9(1):147–169.
going environmental changes and biological mutations
will not affect a species unless they violate or modify Atkins, P. W. (1984). The second law. Scientific Amer-
the species core code in its present ecosystem. Autoen- ican Library.
coding can maintain evolutionary stasis at the species Auerswald, P. E. and Branscomb, L. M. (2003). Valleys
macro-scale despite continuous changes at the micro- of death and Darwinian seas: Financing the inven-
scale involving, for example, ongoing DNA mutations tion to innovation transition in the united states.
and local environmental perturbations. The Journal of Technology Transfer, 28(3):227–
239.
Bank, D., Koenigstein, N., and Giryes, R. (2020). Au-
8 Sustaining the biosphere de- toencoders. arXiv preprint arXiv:2003.05991.
mands changes in collective Barbieri, M. (2012). Codepoiesis–the deep logic of life.
human behavior. Biosemiotics, 5(3):297–299.
The rapid expansion of the human population in the Barbieri, M. (2015). Code biology: A new science of
past ten thousand years owes its onset to the domesti- life. Springer.
cation of species of plants and animals by human inter-
vention; humans chose to select for innovations affect- Bergman, J. (2014). The Darwin Effect: It’s influence
ing other species and environments. on Nazism, Eugenics, Racism, Communism, Cap-
Whether or not one accepts Darwin’s idea of Nat- italism & Sexism. New Leaf Publishing Group.
ural Selection as a "law of nature" (Bradley, 2022; Blaser, M. J. (2014). The microbiome revolution. The
Reed, 1981; Byerly, 1983), the "natural right" of dom- Journal of Clinical Investigation, 124(10):4162–
ination by the most fit has influenced many aspects 4165.
of human culture including ethics, economics, gover-
nance, racial relations, social organization, and educa- Bradley, B. (2022). Natural selection according to dar-
tion (Mayr, 2000; Atkins, 1984; Wyllie, 1959; Auer- win: cause or effect? History and Philosophy of
swald and Branscomb, 2003; Browning et al., 2017; the Life Sciences, 44(2):1–26.
Bergman, 2014). The centrality of domination in Natu- Browning, L., Thompson, K., and Dawson, D. (2017).
ral Selection is problematic both to our understanding From early career researcher to research leader:
and to our behavior. survival of the fittest? Journal of Higher Edu-
Thomas Kuhn has pointed out the blinding power of cation Policy and Management, 39(4):361–377.
entrenched paradigms in science (Kuhn, 1970). Tra-
ditional studies of evolution assume Survival-of-the- Byerly, H. C. (1983). Natural selection as a law: Prin-
Fittest in some form as a given. Misunderstanding ciples and processes. The American Naturalist,
alone, however, is tolerable; the spirit of legitimate 121(5):739–745.
domination underlies much of the irresponsible human Christiansen, M. H. and Kirby, S. E. (2003). Language
behavior that is now changing the biosphere. evolution. Oxford University Press.
Natural Selection is not sufficiently sensitive to the
world wide web of cooperative interactions among Cohen, I. R. (2000). Tending Adam’s Garden: evolving
species and environments required to maintain a bio- the cognitive immune self. Elsevier.
sphere friendly to the well-being of the human species. Cohen, I. R. (2006). Informational landscapes in art,
We hope that an appreciation of Natural Autoencoding science, and evolution. Bulletin of Mathematical
and Survival-of-the-Fitted will provide a rationale for a Biology, 68(5):1213–1229.
revision of human interactions with the biosphere.
Cohen, I. R. (2016). Updating Darwin: information and
entropy drive the evolution of life. F1000Research,
5.
9 Future Research
Cohen, I. R., Atlan, H., and Efroni, S. (2016). Genet-
The role of encoding and decoding in evolution has in-
ics as Explanation: Limits to the Human Genome
spired us to embark on the formulation of new archi-
Project. eLS. John Wiley & Sons, Ltd: Chichester.
tectures and algorithms in generative machine learning.
Computer models of Natural Autoencoding can extend Cohen, I. R. and Efroni, S. (2019). The immune sys-
our understanding of the biosphere and might even pro- tem computes the state of the body: crowd wis-
vide new tools in computer science. dom, machine learning, and immune cell reference
repertoires help manage inflammation. Frontiers
in immunology, 10:10.
Page 9 of 15REFERENCES Cohen and Marron, March 2022
Cohen, I. R. and Harel, D. (2006). Explaining a com- Graves, J. A. M. (1996). Mammals that break the rules:
plex living system: dynamics, multi-scaling and genetics of marsupials and monotremes. Annual
emergence. Journal of the Royal Society Interface, review of genetics, 30(1):233–260.
4(13):175–182.
Grotzinger, J. P., Bowring, S. A., Saylor, B. Z., and
Cohen, I. R. and Marron, A. (2020). The evolution of Kaufman, A. J. (1995). Biostratigraphic and
universal adaptations of life is driven by universal geochronologic constraints on early animal evolu-
properties of matter: energy, entropy, and interac- tion. Science, 270(5236):598–604.
tion. F1000Research, 9.
Gruder-Adams, S. and Getz, L. L. (1985). Comparison
Crutchfield, J. P. (1994). The calculi of emergence:
of the mating system and paternal behavior in mi-
computation, dynamics and induction. Physica D:
crotus ochrogaster and m. pennsylvanicus. Journal
Nonlinear Phenomena, 75(1-3):11–54.
of Mammalogy, 66(1):165–167.
Darwin, C. (1860). On the Origin of Species by
Means of Natural Selection, or the Preservation of Hill, R. and Broughton, R. K. (2009). Mapping the un-
Favoured Races in the Struggle for Life. John Mur- derstorey of deciduous woodland from leaf-on and
ray. leaf-off airborne LiDAR data: A case study in low-
land britain. ISPRS Journal of Photogrammetry
Depa, Ł., Kaszyca-Taszakowska, N., Taszakowski, A., and Remote Sensing, 64(2):223–233.
and Kanturski, M. (2020). Ant-induced evolu-
tionary patterns in aphids. Biological Reviews, Hu, S., Dilcher, D. L., Jarzen, D. M., and Taylor, D. W.
95(6):1574–1589. (2008). Early steps of angiosperm–pollinator co-
evolution. Proceedings of the National Academy of
Dietrich, M. R. (2003). Richard Goldschmidt: hope-
Sciences, 105(1):240–245.
ful monsters and other’heresies’. Nature Reviews
Genetics, 4(1):68–74. Jablonka, E. (2011). The entangled (and con-
Doige, C. A. and Day, T. (2012). A typology of under- structed) human bank. Philosophical Transac-
graduate textbook definitions of heat across sci- tions of the Royal Society B: Biological Sciences,
ence disciplines. International Journal of Science 366(1566):784–784.
Education, 34(5):677–700.
Judge, A. and Dodd, M. (2020). Metabolism. Essays
Dupré, J. and O’Malley, M. A. (2013). Varieties of in Biochemistry, 64(4):607–647.
living things: life at the intersection of lineage
and metabolism. In Vitalism and the Scientific Keller, E. F. and Segel, L. A. (1970). Initiation of slime
Image in Post-Enlightenment Life Science, 1800- mold aggregation viewed as an instability. Journal
2010, pages 311–343. Springer. of theoretical biology, 26(3):399–415.
Fisher, R. A. (1958). The genetical theory of natural Koonin, E. V. (2011). The Logic of Chance: the Nature
selection. Ripol Classic. and Origin of Biological Evolution. FT press.
Fitch, W. T. (2006). The biology and evolution of
Kramer, M. A. (1991). Nonlinear principal compo-
music: A comparative perspective. Cognition,
nent analysis using autoassociative neural net-
100(1):173–215.
works. AIChE journal, 37(2):233–243.
Gleick, J. (1993). Genius: The life and science of
Richard Feynman. Vintage. In Prologue. Kritsky, G. (1991). Darwin’s Madagascan hawk moth
prediction. American Entomologist, 37(4):206–
Gokhman, D., Nissim-Rafinia, M., Agranat-Tamir, 210.
L., Housman, G., García-Pérez, R., Lizano, E.,
Cheronet, O., Mallick, S., Nieves-Colón, M. A., Li, Kuhn, T. S. (1970). The structure of scientific revolu-
H., et al. (2020). Differential DNA methylation of tions, volume 111. Chicago University of Chicago
vocal and facial anatomy genes in modern humans. Press.
Nature communications, 11(1):1–21.
Kurosawa, R., Kono, R., Kondo, T., and Kanai, Y.
Goldschmidt, R. (1940). The material basis of evolu- (2003). Diet of jungle crows in an urban landscape.
tion. Yale University Press. ISBN 0-300-02823-7. Global Environmental Research—English Edition,
Goodfellow, I., Bengio, Y., and Courville, A. (2016). 7(2):193–198.
Deep Learning. MIT Press. http://www.
Lim, M. M., Wang, Z., Olazábal, D. E., Ren, X., Ter-
deeplearningbook.org.
williger, E. F., and Young, L. J. (2004). Enhanced
Gould, S. J. and Eldredge, N. (1972). Punctuated equi- partner preference in a promiscuous species by ma-
libria: an alternative to phyletic gradualism. Mod- nipulating the expression of a single gene. Nature,
els in paleobiology, 1972:82–115. 429(6993):754–757.
Page 10 of 15REFERENCES Cohen and Marron, March 2022
Lindsey, H. A., Gallie, J., Taylor, S., and Kerr, B. Rosenberg, E., Koren, O., Reshef, L., Efrony, R., and
(2013). Evolutionary rescue from extinction is con- Zilber-Rosenberg, I. (2007). The role of microor-
tingent on a lower rate of environmental change. ganisms in coral health, disease and evolution. Na-
Nature, 494(7438):463–467. ture Reviews Microbiology, 5(5):355–362.
Lynch, M. (2010). Evolution of the mutation rate. Rosenzweig, C., Karoly, D., Vicarelli, M., Neofotis, P.,
Trends in Genetics, 26(8):345–352. Wu, Q., Casassa, G., Menzel, A., Root, T. L., Es-
trella, N., Seguin, B., et al. (2008). Attributing
Mallet, J. (1995). A species definition for the mod-
physical and biological impacts to anthropogenic
ern synthesis. Trends in Ecology & Evolution,
climate change. Nature, 453(7193):353–357.
10(7):294–299.
Rovelli, C. (2017). Carlo Rovelli — All Reality Is
Margulis, L. (1981). Symbiosis in Cell Evolution: Life
Interaction
and its Environment on the Early Earth. W. H.
. https://www.wnyc.org/story/
Freeman.
59a21fbf4616dd86cb8fc341; WNYC Radio
Mayr, E. (2000). Darwin’s influence on modern Program; Accessed March 2022.
thought. Scientific American, 283(1):78–83.
Sachs, J. L., Mueller, U. G., Wilcox, T. P., and Bull,
Mellars, P. (2004). Neanderthals and the mod- J. J. (2004). The evolution of cooperation. The
ern human colonization of Europe. Nature, Quarterly review of biology, 79(2):135–160.
432(7016):461–465.
Sancar, A., Lindsey-Boltz, L. A., Ünsal-Kaçmaz, K.,
Melstrom, K. M. (2017). The relationship between diet and Linn, S. (2004). Molecular mechanisms of
and tooth complexity in living dentigerous sauri- mammalian dna repair and the dna damage check-
ans. Journal of morphology, 278(4):500–522. points. Annual review of biochemistry, 73(1):39–
85.
Metzler, D., Knief, U., Peñalba, J. V., and Wolf,
J. B. (2020). Assortative mate choice and epistatic Scott-Phillips, T. C., Laland, K. N., Shuker, D. M.,
mating-trait architecture induces complex hybrid- Dickins, T. E., and West, S. A. (2014). The niche
zone movement. bioRxiv. construction perspective: a critical appraisal. Evo-
lution, 68(5):1231–1243.
Nagasawa, M., Mitsui, S., En, S., Ohtani, N., Ohta,
M., Sakuma, Y., Onaka, T., Mogi, K., and Shannon, C. E. (1948). A mathematical theory of com-
Kikusui, T. (2015). Oxytocin-gaze positive loop munication. The Bell system technical journal,
and the coevolution of human-dog bonds. Science, 27(3):379–423.
348(6232):333–336.
Simard, S. W. (2018). Mycorrhizal networks facili-
O’Toole, G., Kaplan, H. B., and Kolter, R. (2000).
tate tree communication, learning, and memory.
Biofilm formation as microbial development. An-
In Memory and learning in plants, pages 191–213.
nual Reviews in Microbiology, 54(1):49–79.
Springer.
Pigliucci, M. and Müller, G. B. (2010). Elements of an
Slack, J. M. and Dale, L. (2021). Essential developmen-
extended evolutionary synthesis. Evolution: The
tal biology. John Wiley & Sons.
extended synthesis, pages 3–17.
Poelstra, J. W., Vijay, N., Bossu, C. M., Lantz, H., Socher, R., Pennington, J., Huang, E. H., Ng, A. Y.,
Ryll, B., Müller, I., Baglione, V., Unneberg, P., and Manning, C. D. (2011). Semi-supervised re-
Wikelski, M., Grabherr, M. G., et al. (2014). cursive autoencoders for predicting sentiment dis-
The genomic landscape underlying phenotypic in- tributions. In Proceedings of the 2011 conference
tegrity in the face of gene flow in crows. Science, on empirical methods in natural language process-
344(6190):1410–1414. ing, pages 151–161.
Puigbò, P., Wolf, Y. I., and Koonin, E. V. (2010). The Soussi, T. (2000). The p53 tumor suppressor gene: from
tree and net components of prokaryote evolution. molecular biology to clinical investigation. Annals
Genome biology and evolution, 2:745–756. of the New York Academy of Sciences, 910(1):121–
139.
Purvis, A., Gittleman, J. L., Cowlishaw, G., and
Mace, G. M. (2000). Predicting extinction risk Striedter, G. F. (2005). Principles of brain evolution.
in declining species. Proceedings of the royal so- Sinauer associates.
ciety of London. Series B: Biological Sciences,
267(1456):1947–1952. Vanchurin, V., Wolf, Y. I., Katsnelson, M. I., and
Koonin, E. V. (2022a). Toward a theory of evolu-
Reed, E. S. (1981). The lawfulness of natural selection. tion as multilevel learning. Proceedings of the Na-
The American Naturalist, 118(1):61–71. tional Academy of Sciences, 119(6):e2120037119.
Page 11 of 15REFERENCES Cohen and Marron, March 2022
Vanchurin, V., Wolf, Y. I., Koonin, E. V., and Katsnel-
son, M. I. (2022b). Thermodynamics of evolution
and the origin of life. Proceedings of the National
Academy of Sciences, 119(6):e2120042119.
Vitousek, P. M., Cassman, K., Cleveland, C., Crews,
T., Field, C. B., Grimm, N. B., Howarth, R. W.,
Marino, R., Martinelli, L., Rastetter, E. B., et al.
(2002). Towards an ecological understanding
of biological nitrogen fixation. Biogeochemistry,
57(1):1–45.
Waddington, C. H. (1953). Genetic assimilation of an
acquired character. Evolution, pages 118–126.
Westbrook, C. J., Cooper, D. J., and Baker, B. W.
(2006). Beaver dams and overbank floods influence
groundwater–surface water interactions of a rocky
mountain riparian area. Water resources research,
42(6).
Wildman, D. E., Chen, C., Erez, O., Grossman, L. I.,
Goodman, M., and Romero, R. (2006). Evolu-
tion of the mammalian placenta revealed by phy-
logenetic analysis. Proceedings of the National
Academy of Sciences, 103(9):3203–3208.
Wolf, J. B., Bayer, T., Haubold, B., Schilhabel, M.,
Rosenstiel, P., and Tautz, D. (2010). Nucleotide di-
vergence vs. gene expression differentiation: com-
parative transcriptome sequencing in natural iso-
lates from the carrion crow and its hybrid zone
with the hooded crow. Molecular ecology, 19:162–
175.
Wyllie, I. G. (1959). Social Darwinism and the busi-
nessman. Proceedings of the American Philosoph-
ical Society, 103(5):629–635.
Acknowledgments
We thank Gil Egozi, Guy Frankel, David Harel, Eu-
gene Koonin, Antonio López-Pinto, Eugene Rosenberg,
and Smadar Szekely for valuable discussions and sug-
gestions.
Author contributions
IRC and AM contributed equally to the paper.
Competing interests
The authors have no competing interests.
Page 12 of 15You can also read

















































