Evaluating Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) Response to Methyl Eugenol: Comparison of Three Common Bioassay Methods
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Economic Entomology, 115(2), 2022, 556–564
https://doi.org/10.1093/jee/toac018
Advance Access Publication Date: 17 March 2022
Research
Ecology and Behavior
Evaluating Bactrocera dorsalis (Hendel) (Diptera:
Tephritidae) Response to Methyl Eugenol: Comparison of
Three Common Bioassay Methods
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
Sheina B. Sim,1, Keena M. Curbelo, Nicholas C. Manoukis, and Dong H. Cha
USDA Agricultural Research Service, Daniel K. Inouye US Pacific Basin Agricultural Research Center, Tropical Crop and Commodity
Protection Research Unit, 64 Nowelo Street, Hilo, HI, USA, and 1Corresponding author, e-mail: sheina.sim@usda.gov
Subject Editor: Nikolaos Papadopoulos
Received 3 May 2021; Editorial decision 17 January 2022
Abstract
Insect responses to chemical attractants are often measured using olfactory bioassays prior to testing in field
experiments. The attraction of sexually mature male Bactrocera dorsalis to methyl eugenol (ME) and the loss
of attraction by ME pre-fed males have been demonstrated in laboratory bioassays and field trapping studies.
It has been suggested that ME nonresponsiveness can be exploited to improve the effectiveness of B. dor-
salis management programs by protecting sterile males from ME-based control measures. Currently, work is
underway to identify alternatives that reduce or eliminate ME response. To support the development of com-
pounds and evaluation of their effect on B. dorsalis attraction to ME, we compared the effectiveness of three
common bioassay methods that have been used to measure lure response in Bactrocera flies under controlled
conditions (choice assays using Y-tube [Y], small-cage arena [SC], and rotating carousel field-cage [RC]) to
determine which bioassay method is efficient and reliable. A series of bioassays comparing ME-exposed and
ME-naïve wild-type and genetic sexing strain males showed that the RC and SC were effective at both observing
attraction to ME and detecting a significant reduction in ME response from ME-exposed males. However, the
male attraction to ME and a significant decrease in response to ME after ME feeding was not observed in our
Y-tube assays. These suggest that RC and SC are preferable options to evaluate ME non-responsiveness in
B. dorsalis, and that Y-tube tests are difficult to administer correctly.
Key words: olfaction, behavior, bioassay, olfactometer
The Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: the population, both depend on methyl eugenol (ME) [4-allyl-1,2-
Tephritidae), is a highly invasive pest of a broad range of agricultural dimethoxybenzene] (Howlett 1915, Metcalf et al. 1975, Manoukis
fruit crops. Because of its destructive nature, it has caused signifi- et al. 2019), a chemical compound naturally produced by some
cant direct crop loss in addition to indirect losses due to quarantine flowers and hypothesized to be a metabolic precursor to male sexual
restriction on potentially infested fruit which results in diminished maturity (Nishida et al. 1988, Shelly et al. 2005, Jang et al. 2013,
market access across the globe (Heather and Hallman 2008, Dohino Vargas et al. 2014). It is a compound that elicits a strong attraction
et al. 2016). Management options of B. dorsalis include chemical and phagostimulation response from male B. dorsalis (Wong et al.
insecticide application (Orankanok et al., 2007, Vargas et al. 2010) 1989), an attribute that is key to its utility as the attractant in detec-
and strict quarantine efforts (Follett and Armstrong 2004, Hsu et al. tion networks and male annihilation technique bait stations (Gilbert
2018), but the most effective tools against B. dorsalis have been et al. 2013). Because ME is such an effective attractant for male
combinations of early detection through male surveillance, and the B. dorsalis, sterile insect technique (Knipling 1955) has been applied
male annihilation technique (Steiner and Lee 1955). infrequently against B. dorsalis compared with other major tephritid
Early detection and the male annihilation technique, a method pests like Ceratitis capitata (Wiedermann) (Diptera: Tephritidae)
which consists of attracting and killing a large proportion of the sexu- and Zeugodacus (Bactrocera) cucurbitae (Coquillett) (Diptera:
ally mature males in a fruit fly population to the point of crashing Tephritidae) whose male lures are not as potent as methyl eugenol.
Published by Oxford University Press on behalf of Entomological Society of America 2022. 556
This work is written by (a) US Government employee(s) and is in the public domain in the US.Journal of Economic Entomology, 2022, Vol. 115, No. 2 557
For example, the successful eradication program in Okinawa and to ME and assess the ability of three bioassay formats to quantify
neighboring islands relied on the sterile insect technique and other ME attraction by B. dorsalis, sexually mature adult male B. dorsalis
measures to eliminate Z. cucuribitae, but mainly the male annihila- were exposed to ME as a treatment and their response to ME after
tion technique against B. dorsalis (Koyama et al 1984, 2004). ME treatment was quantified using Y-tube, small-cage, and rotating
The use of the sterile insect technique decreases reproduction in carousel bioassays. Control flies, which represented flies not previ-
wild populations through over-flooding by sterile flies resulting in a ously exposed to ME (ME-naïve), were also tested. Results from
suppressed or eradicated wild population (Knipling 1955). Though these series of tests showed agreement with the previous field studies
less common compared with other tephritid control programs as by Shelly and colleagues (1994, 2020), but differences between bio-
mentioned above, release of sterile B. dorsalis has been successful as assay types, and effects of interactions that have not been previously
a component of pest management programs in previous suppression described. The results from the work described have implications for
(Sutantawong et al. 2002, Orankanok et al. 2007) and eradication usage of various bioassay formats in future studies and pest manage-
efforts (Steiner et al. 1970, Habu et al. 1984, Koyama et al. 1984). ment which are discussed.
The sterile insect technique remains an important tool against fruit
flies because it reduces the need for chemical control, is environmen-
tally friendly, and is reversible. However, one of the limits to its appli- Materials and Methods
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
cation can be cost- which can be reduced via increased efficiency and Study Flies
combinations of control measures (Mumford 2005). Contemporary Wild-type B. dorsalis pupae and genetic sexing strain B. dorsalis of
fruit fly sterile insect technique in tephritids such as C. capitata and the same age were obtained from the Daniel K. Inouye U.S. Pacific
Anastrepha ludens (Loew) (Diptera: Tephritidae) is often facilitated Basin Agricultural Research Center insect colony in Hilo Hawai‘i.
by male-only releases of a genetic sexing strain, which harbor select- This wild-type line originates from B. dorsalis collected in Puna,
able phenotypes such as a sexually dimorphic color morph or con- Hawai’i island in 1984, and has been kept in the laboratory on arti-
ditional lethality that allow for the rapid separation of males from ficial diet with the occasional infusion of wild-collected flies since its
females en masse (Franz 2005). inception. Adults are held in mixed cages at a density of ~50,000 per
For B. dorsalis, the combination of the male annihilation tech- cage (0.6 × 1.18 × 1.32 m in size, w × h × d). By contrast, the genetic
nique with the sterile insect technique holds significant potential sexing strain originated from cross between an irradiated wild-type
to increase the efficacy and efficiency though at a minimum, ap- colony that was originally established from wild-collected flies from
plication of the two techniques must be separated in time and/or Pukalani, Maui that had been maintained in colony for approxi-
space. However, these two methods can be made less antagonistic if mately 60 generations and induced with a reciprocal autosomal
nonresponse to ME can be induced in sterile male B. dorsalis. Shelly and Y-chromosome translocation (McCombs and Saul 1995) and a
(1994, 2020) reported that prior ME feeding by adult males resulted white puparium strain (McCombs and Saul 1992) with subsequent
in a decrease in attraction to ME by sexually mature male wild-type generations being selected for a fixed and stable translocation re-
and genetic sexing strain B. dorsalis. sulting in sex-linkage of the pupal color phenotype. Adults of the
To evaluate the possibility of combining the male annihilation genetic sexing strain are maintained in 30 cm3 wire mesh cages at a
technique and the sterile insect technique, and also to test the cur- density of 500 adults per cage with the entire colony consisting of
rently available genetic sexing strain and the next generation of gen- several cages of nonoverlapping generations. For these experiments,
etic sexing strain (Sim et al. 2019) for their compatibility with the adults eclosed and were maintained in 30 cm3 wire mesh cages and
male annihilation technique, it is critical to be able to accurately separated by sex within 3 d post emergence. All flies were main-
screen their behavioral response to ME. The behavioral response tained on a diet of sugar and yeast hydrolyzed protein in a ratio of
of an individual male to ME will be influenced by several of the 3:1 (Vargas 1989). Flies were held at 24–28°C, 60–90% RH, and re-
steps in the chain of events involving physiological, transcriptional, ceived broad-spectrum artificial light under a photoperiod of 12:12
and other subindividual level processes in the context of the biotic (L:D) h and were maintained under identical conditions until con-
and abiotic environment and can be most accurately determined in current bioassays were administered.
the field. However, field screening of chemical attractants is time-
consuming and labor-intensive, and it may be prohibited due to pol-
icies related to the release of GMO or transgenic insects. In tephritid Bioassays for ME Response: Rotating-Carousel,
attractant research, a variety of olfactometers and bioassays have Small Cage, and Y-tube Olfactometer
been used to facilitate the quantification of behavioral response to Male B. dorsalis (13–16 days old) response to ME was evaluated
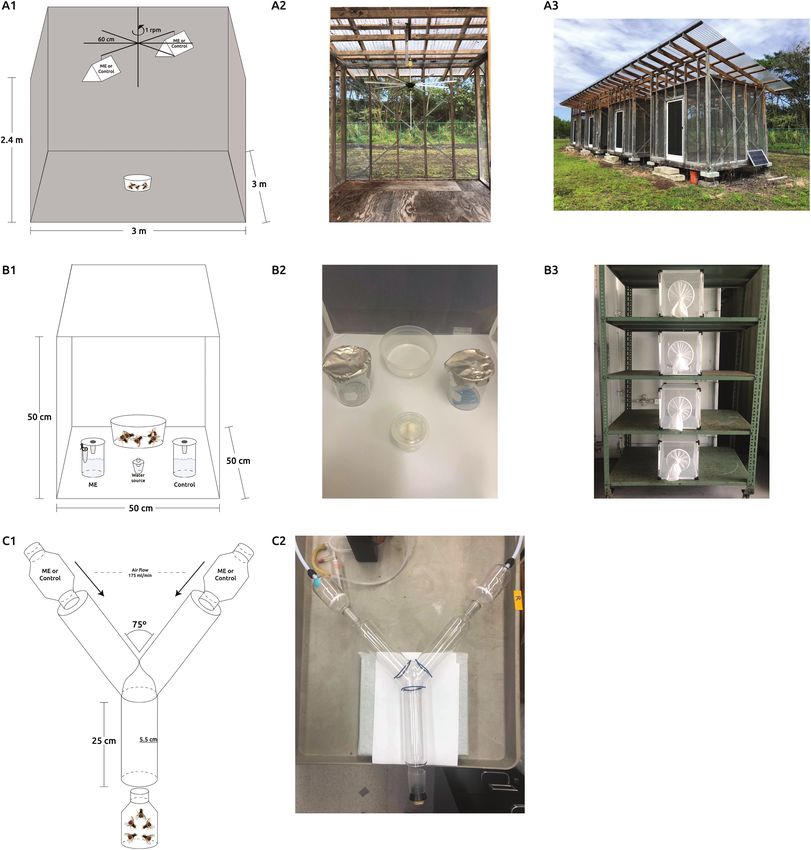
attractant, including bioassays administered via Y-tube (Manoukis using rotating-carousel (RC; Fig. 1A), small-cage (SC; Fig. 1B), and
et al. 2015), small cage (Cha et al. 2012, Roh et al. 2021), and the Y-tube (Fig. 1C) bioassays in 9 replicates with varying sample sizes
rotating carousel (Gothilf and Galun 1982, Fezza and Shelly 2020). of 100, 50, and 30 flies per replicate, for a total of 900, 450, and
The one prominent drawback of these assays is that while they are 270 flies tested in each of the bioassays respectively over an eight-
easier and faster to conduct than field testing, discrepancies between month period from August 2019 to April 2020. RC bioassays were
laboratory and field attraction of insects to semiochemical have been performed outside under a rain cover as described in (Fezza and
common (Knudsen et al. 2008, Cha et al. 2018). Thus, the overall Shelly 2020). Briefly, RC is 3 m L × 3 m W × 2.4 m H (20.7 m3)
goal of this study was to compare effectiveness of these common screened wood-frame with a translucent rain cover, containing a
bioassay methods in quantifying male B. dorsalis response to ME to motorized rotating six-armed ‘lazy Susan’ (diameter approximately
accurately match the response established in field trials. 60 cm) mounted near the ceiling of the cage. Two Jackson traps
Here we explore the behavioral attraction of B. dorsalis males with a bottom sticky panel were hung from opposite arms of the ca-
to ME using three behavioral bioassays: Y-tube, small cage, and rousel 60 cm apart, one baited with a cotton wick impregnated with
rotating-carousel. Attraction to ME was quantified for flies from two 20 μl of absolute ME. The other Jackson trap contained the wick
colonies: A wild-type colony and a genetic sexing strain. To quan- but no ME, and the carousel was set to spin at 1 rotation per mi-
tify the attraction of the wild-type strain and genetic sexing strain nute (0.06 m/s). For each replicate, 100 males from each treatment558 Journal of Economic Entomology, 2022, Vol. 115, No. 2 Fig. 1. Diagrams of the rotating-carousel (A1), small cage (B1), and Y-tube (C1) bioassays and relative locations of the methyl eugenol (ME) and the control trap Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022 or arm. Diagrams are not to scale. Photos of singular setups (A2, B2, and C2). Photos of concurrent bioassays and sample conditions (A3 and B3). were released, and after 24 h the number of flies in each trap was males from each treatment were released into the cage. After 24 h recorded. the locations of the individuals were recorded. The position of the The SC bioassays were conducted using 50 cm × 50 cm × 50 cm lure was switched between replicates. Outdoor environmental con- mesh cages (0.125 m3) as described previously (Cha et al. 2012) and ditions over the course of the experiment ranged from a temperature performed outside under a cover. Two 150 ml glass beakers were of 17°C–30°C with a mean of about 23.1°C and a relative humidity placed in the cage 25 cm apart, one containing a 1.5 ml tube with range of 65% to 95% and a mean of 81.3%. a cotton wick with 10 μl of absolute ME, the other without ME. Y-tube bioassays were conducted using Y-tube glass olfactometer The 1.5 ml tube had a hole (3 mm diameter) drilled into the cap (5.5 cm diameter and 25 cm arm and base length) in a laboratory to allow the ME plume to escape. Both 150 ml beakers contained with fluorescent full-spectrum overhead lighting, and temperature 50 ml of a solution containing 50 μl unscented dish soap dissolved in and humidity that ranged from 16.1°C to 25.2°C and 44.4% to 100 ml water and covered with aluminum foil with an entry tunnel 76.7% respectively with a mean of 21.0°C and 56.5% respectively which allowed the flies to enter but not exit. In each replicate, 50 across all replicates. The chamber of one arm contained a cotton
Journal of Economic Entomology, 2022, Vol. 115, No. 2 559
wick loaded with 5 μl of ME, a loading demonstrated to be effective colony source treatments as two fixed independent variables, and
in measuring behavior in previous studies (Manoukis et al. 2015), replicate as a random variable with a binomial distribution and logit
while the chamber of the other arm remained empty. Carbon-filtered link function and maximum likelihood estimation (Proc Glimmix;
air flow through each arm was set at 175 ml/min and allowed to SAS Institute 2009). The means were compared using the Tukey-
equilibrate for 5 min before the introduction of flies. A replicate con- Kramer test (SAS Institute 2009).
sisted of two trials, one with the odor on the right arm, and the other
with the odor on the left in a randomized sequence. Between each
trial, the olfactometer was cleaned using an unscented soap solution, Results
a DI water rinse, an acetone rinse, and air dried with a space heater Male Response to ME
to ensure no odor from the previous cohort remained in the Y-tube. Attraction to ME, where attraction is defined as a significantly
For each trial, a cohort of 15 male B. dorsalis, representing half the greater number of flies in the ME trap or arm than the control
flies in a replicate, were released into the base tube, and the location trap or arm, by male B. dorsalis not previously fed ME (con-
of each individual was recorded every minute for 15 min. The final trol flies) was observed in RC and SC but not from Y-tube bio-
spatial distribution of flies in the Y-tube after this period was used assays (Fig 2; Supp Table 1 [online only]). Both RC and SC assays
in the analysis. were effective at detecting male attraction to ME regardless of
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
ME feeding and colony source treatment with the distribution
Effect of Colony Type and ME Feeding on B. dorsalis of flies between ME-baited trap versus the control trap signifi-
Male Response to ME cantly skewed towards the ME-baited trap (Fig. 2). By contrast,
Three to 5 d prior to ME feeding treatment, cohorts of genetic sexing the Y-tube bioassay did not detect male attraction to ME. For
strain and wild-type males were separated into cages of 100, 50, and ME-fed wild-type and genetic sexing strain males, there were sig-
30 for behavior testing in the RC enclosure, SC arena, and Y-tube, re- nificantly more males in the control arm of the Y-tube than ME
spectively (Fig. 1) with a total of nine replicates per treatment/colony treated arm (Fig. 2)
combination. Males in the ME feeding treatment were offered ME
as described by Shelly et al. (2005). Briefly, two hours prior to be- Effect of ME Feeding and Colony of Origin on ME
havioral testing, sexually mature males (13–16 days post eclosion) Response
were moved into a laboratory hood in a separate room and offered In RC and SC bioassays, ME-feeding resulted in 75% and 65% de-
ME via an ME-loaded cotton wick for 1 h. The flies to be tested in crease (RC: F1,24 = 283.47, P < 0.0001; SC: F1,24 = 127.90, P < 0.0001)
the Y-tube and SC were exposed to a cotton wick loaded with 100 μl in number of total B. dorsalis males that made a choice (i.e., trapped
ME while the flies to be tested in the RC were exposed to a wick in either ME baited trap or control trap) with 95.6% and 67.9% of
loaded with 200 μl ME to accommodate the greater number of flies those males, respectively, in ME baited traps. However, the same ef-
in the RC replicates. Males had access to the ME treatment for 1 h as fect was not observed from Y-tube assays (F1,24 = 0.82, P = 0.3736).
90% of mature males were expected to feed on ME within 1 h under Among the males that made a choice in all three bioassays, ME
these conditions. Control males were held under the same conditions feeding decreased the total proportion of flies attracted to ME by
in a separate fume hood for the same period of time, but without 3, 27, and 38% compared to flies that were ME-naive in RC, SC
ME exposure. All males always had access to water (via a separate and Y-tube bioassays, respectively (RC: F1,24 = 5.02, P = 0.346; SC:
covered container with dental wick) and adult diet ad libitum prior F1,22 = 55.97, P < 0.0001; Y-tube: F1,23 = 16.46, P = 0.0005). A gen-
to the experiment. eralized linear model using treatment as a fixed effect showed that
the RC and SC bioassays revealed a significant decrease in attrac-
Statistical Analyses tion to ME after ME feeding. In the RC and SC bioassays, a 24.4%
For choice tests in RC, SC, and Y-tube olfactory bioassays, B. dor- (F1,24 = 283.47, P < 0.0001) and 15.7% (F1,24 = 127.9, P < 0.0001)
salis male preference between ME treatment versus control was ana- decrease in attraction response was observed as a result of ME
lyzed using χ 2 test for a deviation from an even distribution of flies feeding (Fig. 3 RC: Treatment, SC: Treatment). The effect of ME
between two choices using base R functions and tidyverse packages feeding on attraction response was not observed in the Y-tube bio-
in R (Wickham et al. 2019, R Development Core Team 2020). Only assay (F1,24 = 0.91, P = 0.3736) (Fig. 3 Y: Treatment). The results of
males that made a choice were used for the χ 2 test; in other words, the these statistical analyses show that ME feeding lowers the prob-
individuals that were caught in traps (RC), beakers (SC), or moved ability of capture in the ME trap, but not the probability of capture
upwind into the terminus of the arms (Y). The effect of different ME in the blank trap in the RC. By contrast, the SC and Y-tube bioassays
feeding and source colony treatments on number flies that made a showed a slight but significant increase of flies in the blank trap rela-
choice in RC, SC, and Y assays was analyzed using generalized linear tive to the ME trap.
models in a randomized block design with the number of flies that In the SC bioassay, there was a significant effect of colony origin
made a choice as the dependent variable, ME fed status (ME-naïve vs on the number of males that made a choice with 25% more wild-type
ME fed), and colony source (wild-type vs genetic sexing strain) treat- males making a choice than genetic sexing strain males (F1,24 = 6.04,
ments as two fixed independent variables, and replicate as a blocking P = 0.0215), while there were no significant differences observed
random variable with Poisson distribution and log link function from RC and Y-tube bioassays (RC: F1,24 = 1.65, P = 0.2115; Y-tube:
and maximum likelihood estimation (Proc Glimmix; SAS Institute F1,24 = 1.22, P = 0.2804). Among the males that made a choice in all
2009). The means were compared using the Tukey-Kramer test three bioassays, there was no significant effect of colony origin on
(SAS Institute 2009). The effect of different ME feeding and source the proportion of flies attracted to ME, indicating that, once acti-
colony treatments on male preference for ME over control was ana- vated, genetic sexing strain and wild-type males responded similarly
lyzed using generalized linear model in a randomized block design to ME (for all tests, 0.3609 < P < 0.9091).
with the number of males that chose ME over the total number of For the small-cage bioassay, analysis using colony of origin as a
males that made a choice as the dependent variable, ME feeding, and fixed effect showed that the genetic sexing strain colony had a lower560 Journal of Economic Entomology, 2022, Vol. 115, No. 2
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
Fig. 2. Horizontal bar plots with positive values representing the proportion of flies responding to the methyl eugenol (ME) trap or lure arm emanating from the
center at zero in each bioassay type (rotating-carousel = RC [N = 9, 100 flies per replicate], small-cage = SC [N = 9, 50 flies per replicate], and Y-tube = Y [N = 9,
30 flies per replicate]) and wild-type colony or genetic sexing strain (GSS) and ME treatment or control treatment combination across all replicates. Error bars
represent the standard error, ** denotes P < 0.01, *** denotes P < 0.001, and **** denotes P < 0.0001.
attraction response to ME than the wild-type colony. The SC bio- Effect of Colony and Treatment Interaction
assay revealed a 3.3% decrease (F1,24 = 6.04, P < 0.05) in attraction Analysis investigating the effect of the interaction between
(Fig. 3 SC: Colony). This effect of colony was not observed in the RC colony and treatment showed that only the rotating carousel
(Fig. 3 RC: Colony) or the Y-tube (Fig. 3 Y: Colony). revealed a significant effect of the interaction. In the rotatingJournal of Economic Entomology, 2022, Vol. 115, No. 2 561
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
Fig. 3. Box plots representing numbers of flies attracted to methyl eugenol (ME) for each colony [genetic sexing strain (GSS) or wild-type] or treatment (control
or ME) (RC: Colony, RC: Treatment, SC: Colony, SC: Treatment, Y: Colony, and Y: Treatment) and colony treatment interaction (RC: Interaction).
carousel bioassay, the wild-type flies demonstrated a 34.7% de- Discussion
crease in attraction response as a result of ME feeding while
The results of this study showed that the RC and SC behavioral bio-
genetic sexing flies demonstrated only a 22.9% decrease in at-
assays both yielded qualitatively similar results to field studies in
traction response (F 1,24 = 4.9, df = 24, P < 0.0001) (Fig. 3 RC:
attraction to ME by B. dorsalis, but this was not apparent in the
Interaction).562 Journal of Economic Entomology, 2022, Vol. 115, No. 2
Y-tube assay with no significant differences observed in the number described. A recent study by Fezza and Shelly (2020) performed this
of ME-naïve males choosing between blank and ME-treated arms. comparison using a rotating-carousel bioassay to measure response
The inefficiency of the Y-tube assay was also observed when the ef- to ME. Surprisingly, the difference in response was not detected in
fect of ME feeding on reduced male attraction to ME was tested the RC. Possible explanations for this discrepancy with the Fezza and
using different bioassay formats. Sexually mature B. dorsalis males Shelly (2020) study include the use of wild flies reared from fruit in
fed ME prior to testing showed a decreased attraction to ME in RC contrast to the wild-type colony flies and a difference in amount of ME
and SC, but again this was not observed in the Y-tube assay. used in the lures in this experiment: twice the amount used in Fezza
Of the three methods tested, the Y-tube olfactometer bioassay and Shelly (2020). This suggests that genetic sexing strain B. dorsalis
was the outlier, and we consider that it failed in this study. This might be a good choice for release for the sterile insect technique
was not expected, because though this method is notoriously diffi- concurrently where the male annihilation technique is being admin-
cult to execute correctly, this exact apparatus had been successfully istered as the decrease sensitivity of male genetic sexing strain flies to
applied previously to assay attraction to ME in our laboratory. ME may give them a survival advantage over wild males.
A number of factors may have contributed to the failure, including Behavioral bioassays should be systematically compared for par-
experimental mistakes or colony differences. However, we consider ticular species/lure combinations to ensure optimum sensitivity and
that colder temperatures indoors where the Y assay was conducted throughput. From this study, we favor the use of either RC or SC as
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
(21.0 ± 2.4°C) compared with outdoors (23.1 ± 1.8°C) where RC behavioral bioassays on the attraction of male B. dorsalis to ME.
and SC were housed is the most likely cause. Previous studies with Due to the size and availability of the SC bioassays, the simplicity of
this setup indicate an average of 24.5°C for Y assays (Manoukis et al. execution, minimal equipment requirements, and the high number
2015, 2018), and it is likely that warmer temperature than what we of treatment combinations that can be assayed concurrently, poten-
used in this study is required for the assay to function. tially indoors, this method may be the best choice in most instances.
Beyond the above-mentioned failure, the Y assay has other dis- The larger sample size in the RC bioassay enabled the detection of
advantages. It requires more sophisticated laboratory equipment a small but significant colony and treatment interaction. This, com-
compared with RC and SC, it has limited throughput, and is time bined with the potential for testing more than two attractants at
consuming. One of the reasons it may be technically difficult to once, argues for the utility of a set of RC bioassays where the climate
execute Y is that the insect must travel upwind and leave the entry supports it.
chamber and move toward the lure. Further, flies in the control arm
of the Y-tube may not represent flies repelled by the lure as they must
walk towards the source of the odor plume to escape it. This must Supplementary Data
be taken into account in studies using the Y-tube to test for the repel- Supplementary data are available at Journal of Economic
lency of a chemical compound (Wu et al. 2013, Uniyal et al. 2016). Entomology online.
An important limitation of this experiment is that SC and RC
assays were conducted outdoors, as there are favorable conditions
in Hilo, Hawaii for B. dorsalis year-round. The SC assays should be Acknowledgments
equally effective indoors but note that care must be taken to ensure We would like to acknowledge Keoua Dayton, Lehua Patnaude, Lori
adequate ventilation. SC in a small indoor space could lead to inter- Carvalho, Gwang Hyun Roh, and Tom Fezza for their contributions
ference (Elkinton and Cardé 1988), making it difficult for the males to administering the bioassays, Angela Kauwe and Stephanie Gayle
to find the ME source. This would impact the efficacy of the SC assay. for providing genetic sexing strain and wild-type colony flies, and
It is common for researchers initiating experiments that include a Boyd Mori and Bruce Mackey for their contributions to the statis-
behavioral response to include a laboratory component, and it may tical analyses performed in this study. This work was conducted, in
require significant time and effort to establish an assay that reliably part, to support the Coordinated Research Project number for IAEA
measures the phenotype of interest with a usable level of sensitivity support #23187/R0 sponsored by the International Atomic Energy
(Little et al. 2021). The ideal behavioral bioassay for testing the at- Agency and directed by Rui Cardoso Pereira and Carlos Caceres.
traction of insects to a lure is robust, consistent, time-efficient, able to The US Department of Agriculture, Agricultural Research Service
accept variable throughput, and would accurately capture responses is an equal opportunity/affirmative action employer and all agency
as they occur under natural conditions. We found the SC and the RC services are available without discrimination. The authors declare
to rank highly on these characteristics. Both were robust to small dif- no conflicts of interest, no disputes over the ownership of the data
ferences between days (at least under the stable climatic conditions at presented in this paper, and all contributions have been attributed
our research site), were simple and time-effective, and displayed good appropriately via co-authorship or acknowledgment as appropriate
consistency in capturing expected lure response differences between to the situation. This research used resources provided by the SCINet
ME fed and control males of both colonies. The larger RC cages have project of the USDA Agricultural Research Service, and was funded
a slight edge in being able to support a higher number of maximum by ARS project number 2040-22430-027-00-D.
insects and might be considered slightly more consistent (lower vari-
ance), but they were limited in the number of replicates by their large
size: It is logistically more practical to set up a large number of small Data Availability
cages to increase counts per replicate. Measurement of attraction be-
The raw data is included as Supp Table 2 (online only).
havior in SC assays can also be conducted indoors in locations with
less moderate climate than Hawaii, though from our experience with
the Y-tube assay we suggest warmer temperatures than 21°C. References Cited
The difference in attraction between control flies and ME fed flies Cha, D. H., T. Adams, H. Rogg, and P. J. Landolt. 2012. Identification and
is an effect that is congruent with previous field studies (Shelly 1994, field evaluation of fermentation volatiles from wine and vinegar that me-
2020), but the difference in response to ME between the wild-type diate attraction of spotted wing drosophila, Drosophila suzukii. J. Chem.
colony and genetic sexing strain colony flies had not been previously Ecol. 38: 1419–1431.Journal of Economic Entomology, 2022, Vol. 115, No. 2 563
Cha, D. H., G. M. Loeb, C. E. Linn, Jr., S. P. Hesler, and P. J. Landolt. 2018. technique against Bactrocera dorsalis (Diptera: Tephritidae) at varying ap-
A multiple-choice bioassay approach for rapid screening of key attractant plication densities. PLoS One. 14: e0213337.
volatiles. Environ. Entomol. 47: 946–950. McCombs, S. D., and S. H. Saul. 1992. Linkage analysis of five new genetic
Dohino, T., G. J. Hallman, T. G. Grout, A. R. Clarke, P. A. Follett, markers of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae).
D. R. Cugala, D. Minh Tu, W. Murdita, E. Hernandez, R. Pereira, et al. J. Hered. 83: 199–203.
2016. Phytosanitary treatments against Bactrocera dorsalis (Diptera: McCombs, S. D., and S. H. Saul. 1995. Translocation-based genetic sexing
Tephritidae): current situation and future prospects. J. Econ. Entomol. system for the oriental fruit-fly (Diptera: Tephritidae) based on pupal color
110: 67–79. dimorphism. Ann. Entomol. Soc. Am. 88: 695–698.
Elkinton, J. S., and R. T. Cardé. 1988. Effects of intertrap distance and wind Metcalf, R. L., W. C. Mitchell, T. R. Fukuto, and E. R. Metcalf. 1975
direction on the interaction of gypsy moth (Lepidoptera: Lymantriidae) Attraction of the oriental fruit fly, Dacus dorsalis, to methyl eugenol and
pheromone-baited traps. Environ. Entomol. 17: 764–769. related olfactory stimulants. Proc. Natl. Acad. Sci. USA. 72: 2501
Fezza, T. J., and T. E. Shelly. 2020. Comparative dispersal and survival of Mumford, J. D. 2005. Application of benefit/cost analysis to insect pest control
male oriental fruit flies (Diptera: Tephritidae) from wild and genetic sexing using the sterile insect technique, pp. 481–498. In V. A. Dyck, J. Hendrichs
strains. Int. J. Trop. Insect Sci. 41: 751–757. and A. S. Robinson (eds.), Sterile insect technique: principles and practice in
Follett, P. A., and J. W. Armstrong. 2004. Revised irradiation doses to con- area-wide integrated pest management. Springer Netherlands, Dordrecht.
trol melon fly, Mediterranean fruit fly, and oriental fruit fly (Diptera: Nishida, R., K. H. Tan, M. Serit, N. H. Lajis, A. M. Sukari, S. Takahashi, and
Tephritidae) and a generic dose for tephritid fruit flies. J. Econ. Entomol. H. Fukami. 1988. Accumulation of phenylpropanoids in the rectal glands
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022
97: 1254–1262. of males of the Oriental fruit fly, Dacus dorsalis. Experientia. 44: 534–536.
Franz, G. 2005. Genetic sexing strains in Mediterranean fruit fly, an example Orankanok, W., S. Chinvinijkul, S. Thanaphum, P. Sitilob, and W. R. Enkerlin.
for other species amenable to large-scale rearing for the sterile insect tech- 2007. Area-wide integrated control of oriental fruit fly Bactrocera dorsalis
nique, pp. 427–451. In V. A. Dyck, J. Hendrichs and A. S. Robinson (eds.), and guava fruit fly Bactrocera correcta in Thailand. In V. Mjb, A. S. Robinson
Sterile insect technique. Springer, Dirdrecht, The Netherlands. and J. Hendrichs (eds.), Area-wide control of insect pests: from research to
Gilbert, A. J., R. R. Bingham, M. A. Nicolas, and R. A. Clark. 2013. Insect field implementation. Springer, Dordrecht, The Netherlands.
trapping guide, Thirteenth ed., CDFA, Sacramento, CA, USA. R Development Core Team. 2020. R: a language and environment for stat-
Gothilf, S., and R. Galun. 1982. Olfactometer and trap for evaluating attract- istical computing computer program, version By R Development Core
ants for the Mediterranean fruit fly, Ceratitis capitata. Phytoparasitica. 10: Team, Vienna, Austria.
79–84. Roh, G. H., P. E. Kendra, and D. H. Cha. 2021. Preferential attraction of
Habu, N., M. Iga, and K. I. Numazawa. 1984. An eradication program of oviposition-ready oriental fruit flies to host fruit odor over protein food
the oriental fruit fly, Dacus dorsalis Hendel (Diptera: Tephritidae), in the odor. Insects 12: 909–921.
Ogasawara (Bonin) Islands: I. eradication field test using a sterile fly re- SAS Institute. 2009. SAS/STAT 9.2 User’s Guide, 2nd ed. SAS Institute Inc,
lease method on small islets. Appl. Entomol. Zool. 19: 1–7. Cary, NC, USA.
Heather, N. W., and G. J. Hallman. 2008. Pest management and phytosanitary Shelly, T. E. 1994. Consumption of methyl eugenol by male Bactrocera dorsalis
trade barriers. CAB International, Wallingford, United Kingdom. (Diptera: Tephritidae) - incidence of repeat feeding. Fla. Entomol. 77: 201–208.
Howlett, F. M. 1915. Chemical reactions of fruit-flies. Bull. Entomol. Res. 6: Shelly, T. E. 2020. Evaluation of a genetic sexing strain of the oriental fruit fly
297–305. as a candidate for simultaneous application of male annihilation and sterile
Hsu, Y.-L., S.-C. Chen, K.-W. Lin, C.-C. Shiesh, C.-H. Lin, and W.-B. Yeh. insect techniques (Diptera: Tephritidae). J. Econ. Entomol. 113: 1913–1921.
2018. Quarantine vapor heat treatment of papaya fruit for Bactrocera dor- Shelly, T. E., J. Edu, and E. Pahio. 2005. Influence of diet and methyl eugenol
salis and Bactrocera cucurbitae (Diptera: Tephritidae). J. Econ. Entomol. on the mating success of males of the oriental fruit fly, Bactrocera dorsalis
111: 2101–2109. (Diptera: Tephritidae). Fla. Entomol. 88: 307–313.
Jang, E. B., A. Ramsey, and L. A. Carvalho. 2013. Performance of methyl Sim, S. B., A. N. Kauwe, R. E. Y. Ruano, P. Rendon, and S. M. Geib. 2019. The
eugenol plus matrix plus toxicant combinations under field conditions ABCs of CRISPR in Tephritidae: developing methods for inducing herit-
in Hawaii and California for trapping Bactrocera dorsalis (Diptera: able mutations in the genera Anastrepha, Bactrocera and Ceratitis. Insect
Tephritidae). J. Econ. Entomol. 106: 727–734. Mol. Biol. 28: 277–289.
Knipling, E. F. 1955. Possibilities of insect control or eradication through the Steiner, L. F., and R. K. S. Lee. 1955. Large-area tests of a male-annihilation
use of sexually sterile males. J. Econ. Entomol. 48: 4. method for oriental fruit fly control. J. Econ. Entomol. 48: 311–317.
Knudsen, G. K., M. Bengtsson, S. Kobro, G. Jaastad, T. Hofsvang, and P. Witzgall. Steiner, L. F., W. G. Hart, E. J. Harris, R. T. Cunningham, K. Ohinata, and
2008. Discrepancy in laboratory and field attraction of apple fruit moth D. C. Kamakahi. 1970. Eradication of the oriental fruit fly from the
Argyresthia conjugella to host plant volatiles. Physiol. Entomol. 33: 1–6. Mariana Islands by the methods of male annihilation and sterile insect
Koyama, J., H. Kakinohana, and T. Miyatake. 2004. Eradication of the melon release. J. Econ. Entomol. 63: 131–135.
fly, Bactrocera cucurbitae, in Japan: importance of behavior, ecology, gen- Sutantawong, M., W. Orankanok, W. Enkerlin, V. Wornoauporn, and
etics, and evolution. Annu. Rev. Entomol. 49: 331–349. C. Cáceres. 2002. The sterile insect technique for the control of the oriental
Koyama, J., T. Teruya, and K. Tanaka. 1984. Eradication of the oriental fruit fruit fly, Bactrocera dorsalis (Hendel), in mango orchards in Ratchaburi
fly (Diptera; Tephritidae) from the Okinawa Islands by a male annihilation Province, Thailand, pp. 223–232. In Proceedings of the 6th International
method. J. Econ. Entomol. 77: 468–472. Fruit Fly Symposium, 2002, Stellenbosch, South Africa. International
Little, C. M., P. L. Dixon, D. L. Moreau, T. W. Chapman, and N. K. Hillier. Atomic Energy Agency.
2021. Assessment of attractant lures and monitoring traps for Drosophila Uniyal, A., S. N. Tikar, M. J. Mendki, R. Singh, S. V. Shukla, O. P. Agrawal,
suzukii (Diptera: Drosophidae) using electrophysiology, laboratory choice V. Veer, and D. Sukumaran. 2016. Behavioral response of Aedes aegypti
assays, and field trials. J. Econ. Entomol. 114: 652–675. mosquito towards essential oils using olfactometer. J. Arthropod. Borne
Manoukis, N. C., D. H. Cha, R. M. Collignon, and T. E. Shelly. 2018. Dis. 10: 370–380.
Terminalia larval host fruit reduces the response of bactrocera dorsalis Vargas, R. I. 1989. Rearing: mass production of tephritid fruit flies. World
(Diptera: Tephritidae) adults to the male lure methyl eugenol. J. Econ. Crop Pests. 3B: 141–151.
Entomol. 111: 1644–1649. Vargas, R. I., J. C. Piñero, R. F. L. Mau, E. B. Jang, L. M. Klungness,
Manoukis, N. C., M. Siderhurst, and E. B. Jang. 2015. Field estimates D. O. Mcinnis, E. B. Harris, G. T. Mcquate, R. C. Bautista, and L. Wong.
of attraction of Ceratitis capitata to trimedlure and Bactrocera dor- 2010. Area-wide suppression of the Mediterranean fruit fly, Ceratitis
salis (Diptera: Tephritidae) to methyl eugenol in varying environments. capitata, and the Oriental fruit fly, Bactrocera dorsalis, in Kamuela,
Environ. Entomol. 44: 695–703. Hawaii, J. Insect Sci. 10: 135–152.
Manoukis, N. C., R. I. Vargas, L. Carvalho, T. Fezza, S. Wilson, T. Collier, Vargas, R. I., S. K. Souder, K. Hoffman, J. Mercogliano, T. R. Smith,
and T. E. Shelly. 2019 A field test on the effectiveness of male annihilation J. Hammond, B. J. Davis, M. Brodie, and J. E. Dripps. 2014 Attraction564 Journal of Economic Entomology, 2022, Vol. 115, No. 2
and mortality of Bactrocera dorsalis (Diptera: Tephritidae) to static Wong, T. T. Y., D. O. McInnis, and J. I. Nishimoto. 1989 Relationship of
spinosad me weathered under operational conditions in California and sexual maturation rate to response of oriental fruit fly strains (Diptera:
Florida: a reduced-risk male annihilation treatment. J. Econ. Entomol. Tephritidae) to methyl eugenol. J. Chem. Ecol. 15: 1399–1405.
107: 1362–1369. Wu, H., C. C. Fu, D. D. Yu, J. T. Feng, X. Zhang, and Z. Q. Ma. 2013. Repellent
Wickham, H., M. Averick, J. Bryan, W. Chang, L. D. A. McGowan, R. François, activity screening of 11 kinds of essential oils against Aedes albopictus
G. Grolemund, A. Hayes, L. Henry, and J. Hester. et al. 2019 Welcome to Skuse: microcapsule preparation of Herba Schizonepetae oil and repellent
the tidyverse. J. Open Source Softw. 4: 1686. bioassay on hand skin. Trans. R. Soc. Trop. Med. Hyg. 107: 471–479.
Downloaded from https://academic.oup.com/jee/article/115/2/556/6549821 by guest on 01 July 2022You can also read

















































