Diverse DNA modification in marine prokaryotic and viral communities
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Nucleic Acids Research, 2022 1
https://doi.org/10.1093/nar/gkab1292
Diverse DNA modification in marine prokaryotic and
viral communities
Satoshi Hiraoka1,* , Tomomi Sumida1 , Miho Hirai2 , Atsushi Toyoda3 , Shinsuke Kawagucci2,4 ,
Taichi Yokokawa2 and Takuro Nunoura1
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
1
Research Center for Bioscience and Nanoscience (CeBN), Research Institute for Marine Resources Utilization,
Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Kanagawa 237–0061, Japan,
2
Institute for Extra-cutting-edge Science and Technology Avant-garde Research (X-star), Japan Agency for
Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Kanagawa 237–0061, Japan, 3 Advanced Genomics
Center, National Institute of Genetics, Mishima, Shizuoka 411-8540, Japan and 4 Marine Biodiversity and
Environmental Assessment Research Center (BioEnv), Research Institute for Global Change (RIGC), Japan Agency
for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Kanagawa 237–0061, Japan
Received July 01, 2021; Revised November 30, 2021; Editorial Decision December 11, 2021; Accepted December 17, 2021
ABSTRACT INTRODUCTION
DNA chemical modifications, including methylation, DNA chemical modifications are found in diverse prokary-
are widespread and play important roles in prokary- otes and viruses as well as eukaryotes. DNA methyla-
otes and viruses. However, current knowledge of tion is a representative DNA modification that is cat-
these modification systems is severely biased to- alyzed by DNA methyltransferases (MTases), wherein S-
adenosylmethionine (SAM) provides the methyl group
wards a limited number of culturable prokaryotes,
(1). In prokaryotes, three types of methylation (i.e., N6-
despite the fact that a vast majority of microor- methyladenine [m6A], C5-methylcytosine [m5C] and N4-
ganisms have not yet been cultured. Here, using methylcytosine [m4C]) have been investigated in detail (2).
single-molecule real-time sequencing, we conducted DNA methylation plays a role in regulating gene expression
culture-independent ‘metaepigenomic’ analyses (an and DNA mismatch repair (3–5). These systems have var-
integrated analysis of metagenomics and epige- ious physiological functions, including asymmetric cell di-
nomics) of marine microbial communities. A total vision (6,7), ultraviolet (UV) tolerance (8), motility (9) and
of 233 and 163 metagenomic-assembled genomes virulence of pathogens (10–12). DNA methylation also fa-
(MAGs) were constructed from diverse prokaryotes cilitates cell protection from extracellular DNA (e.g., viral
and viruses, respectively, and 220 modified motifs infection and plasmid transfection), known as restriction-
and 276 DNA methyltransferases (MTases) were iden- modification (RM) systems (13,14). The RM systems are
classified into four types based on subunit composition and
tified. Most of the MTase genes were not geneti-
cofactor requirements. Type I, II and III are composed
cally linked with the endonuclease genes predicted of both MTase and restriction endonuclease (REase) and
to be involved in defense mechanisms against ex- specify non-methylated DNA, while Type IV consists of
tracellular DNA. The MTase-motif correspondence only MTase and specifies modified DNA substrates (15).
found in the MAGs revealed 10 novel pairs, 5 of Some viruses may possess MTases and modify their ge-
which showed novel specificities and experimentally nomic DNA to escape the host Type I, II and III RM
confirmed the catalytic specificities of the MTases. systems. In contrast, Type IV systems have evolved to
We revealed novel alternative specificities in MTases counter the viral anti-RM system, which results in a ‘co-
that are highly conserved in Alphaproteobacteria, evolutionary phage-host arms race’ (2,3). Moreover, evi-
which may enhance our understanding of the co- dence for gene duplication and loss, horizontal gene trans-
evolutionary history of the methylation systems and fer within and between domains, and changes in MTase
the genomes. Our findings highlight diverse unex- sequence specificity, has been frequently noted in the evo-
lution of prokaryotes (16,17). In addition to methylation,
plored DNA modifications that potentially affect the other epigenetic modifications, such as phosphorothioat-
ecology and evolution of prokaryotes and viruses in ion, have recently been reported to have significant effects
nature. on cells, including the maintenance of cellular redox home-
* To whom correspondence should be addressed. Tel: +81 46 867 9397; Email: hiraokas@jamstec.go.jp
C The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial License
(http://creativecommons.org/licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work
is properly cited. For commercial re-use, please contact journals.permissions@oup.com
2 Nucleic Acids Research, 2022
ostasis and epigenetic regulation (18). There has been a were approximately 180 and 140 km offshore from the main
growing interest in exploring the various epigenomic sys- island of Japan and 60 km from each other. Each 50–320 L
tems amongst diverse prokaryotes and viruses owing to of seawater was collected from 5 and 200 m below sea level
their importance in microbial physiology, genetics, evo- (mbsl) at station CM1 (34.2607 N 142.0203 E) and 90 and
lution and disease pathogenicity (19–21). However, most 300 mbsl at station Ct9H (34.3317 N 141.4143 E) (referred
studies rely on a small number of culturable prokaryotic to as CM1 5m, CM1 200m, Ct9H 90m, and Ct9H 300m,
strains, whereas the majority of microbes have yet to be cul- respectively). Sampling permits for expeditions in Japan’s
tured. This limited sample size skews our knowledge of mi- exclusive economic zone were not required, as our work was
crobial epigenomics, particularly in terms of diversity, dis- centered in domestic areas and did not involve endangered
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
tribution, and impact upon ecology and evolution. or protected species. Seawater from 5 mbsl was directly sam-
Recent technological advances have led to the devel- pled using a built-in pumping system from the bottom of
opment of single-molecule real-time (SMRT) sequencing the ship via an intake pipe of approximately 5 m, which
technology as a useful method for detecting DNA mod- was designed for continuous monitoring of sea surface hy-
ifications. Its implementation in PacBio sequencing plat- drography. The valve of the pumping system was opened for
forms has yielded an array of DNA modifications amongst at least 30 min before the start of sampling to thoroughly
prokaryotic (22–27) and viral strains (28,29). The ability of flush the internal water and rinse the pipe. Seawater from 90,
this technology to generate long reads with few context- 200 and 300 mbsl was sampled using 12-L Niskin-X bottles
specific biases (e.g., GC bias) (30) allows the circler con- (General Oceanic, Miami, Florida, USA) in a CTD rosette
sensus sequencing (CCS) method to generate accurate high- system. Vertical profiles of temperature, salinity, and pres-
fidelity (HiFi) reads; a process facilitated by error cor- sure data were obtained using the SBE9plus CTD system
rection within multiple ‘subread’ sequences in each sin- (Sea-Bird Scientific, Bellevue, Washington, USA). The ver-
gle read (31). Based on the innovative SMRT sequenc- tical profiles of dissolved oxygen (DO) concentrations were
ing technique, we conducted culture-independent shotgun obtained using an in situ DO sensor RINKO III (JFE Ad-
metagenomic and epigenomic analyses of freshwater micro- vantech, Hyogo, Japan) connected to the CTD. The vertical
bial communities. This allowed us to determine their natu- profiles of chlorophyll a concentrations were obtained using
ral DNA modification systems and metaepigenomics (32). an in situ Fluorometer RINKO profiler (JFE Advantech).
Apart from PacBio, nanopore sequencing platforms, pro- The seawater samples in the Niskin-X bottles were trans-
duced by Oxford Nanopore Technologies (ONT), can also ferred to sterilized 20 L plastic bags and immediately stored
achieve longer reads that potentially improve metagenomic at 4◦ C until further filtration. Filtration was performed with
assembly with high diversity (33). Accordingly, a hybrid ap- 0.22 m Durapore membrane filters (Merck KGaA, Darm-
proach with HiFi and ONT reads is an ideal way to im- stadt, Germany) after pre-filtration with 5 m Durapore
prove metaepigenomic analysis by enhancing the accuracy membrane filters (Merck KGaA) onboard. The filters were
of identifying organismal modification in highly diverse mi- then immediately stored at temperatures below −30◦ C.
crobial communities.
Here, we conducted metaepigenomic analysis of pelagic
Flow cytometric assessments of prokaryotic cell and viral-like
microbial communities, using the SMRT sequencing tech-
particle abundances
nology, to reveal the epigenomic characteristics of diverse
marine prokaryotes and viruses whose epigenomic status Seawater samples were obtained for flow cytometric assess-
remains largely unknown. The diverse DNA modifications ment of prokaryotic cells and viral-like particle (VLP) abun-
were successfully characterized in numerous metagenomic- dances. The samples were collected every 10–50 m at sta-
assembled genomes (MAGs) from both prokaryotes and tion CM1 and 10–100 m at station Ct9H, fixed with 0.5%
viruses, which were obtained using a combination of PacBio (w/v) glutaraldehyde (final concentration) in 2 ml cryo-
Sequel, ONT GridION and Illumina MiSeq sequencing vials on board, and stored at −80◦ C until further analy-
platforms. Our computational prediction and experimental sis. To assess prokaryotic cell abundance, 200 l of each
assays determined several MTases responsible for the de- sample was stained with SYBR Green I Nucleic Acid Gel
tected methylated motifs, including the novel ones. In par- Stain (Thermo Fisher Scientific, Waltham, Massachusetts,
ticular, a highly conserved methylation system with varied USA) (5 × of manufacturer’s stock, final concentration) at
specificity was identified in Alphaproteobacteria, suggest- room temperature for >10 min. To assess VLP abundance,
ing co-evolution between the methylation systems and the 20 l of each fixed sample was diluted 10 times with TE
genomes. buffer and stained with SYBR Green I (0.5 × of manufac-
turer’s stock, final concentration) for 10 min at 80◦ C. To-
MATERIALS AND METHODS tal prokaryotic cells and VLP abundance in 100 l samples
were determined using an Attune NxT Acoustic Focusing
Seawater sampling
Flow Cytometer (Thermo Fisher Scientific) via the green
Seawater samples were collected at two close pelagic sta- fluorescence versus side scatter plot (34,35).
tions of the Japan Agency for Marine-Earth Science and
Technology (JAMSTEC). Work at these stations, located
DNA extraction and shotgun sequencing
in the northwest Pacific Ocean, yielded these samples dur-
ing the JAMSTEC KM19-07 cruises of the Research Ves- Microbial DNA was retrieved using the DNeasy Power-
sel (R/V) Kaimei in September 2019 (Supplementary Fig- Soil Pro Kit (QIAGEN, Hilden, Germany) according to
ure S1 and Supplementary Table S1). The sampling stations the supplier’s protocol. The filters were cut into 3 mm frag-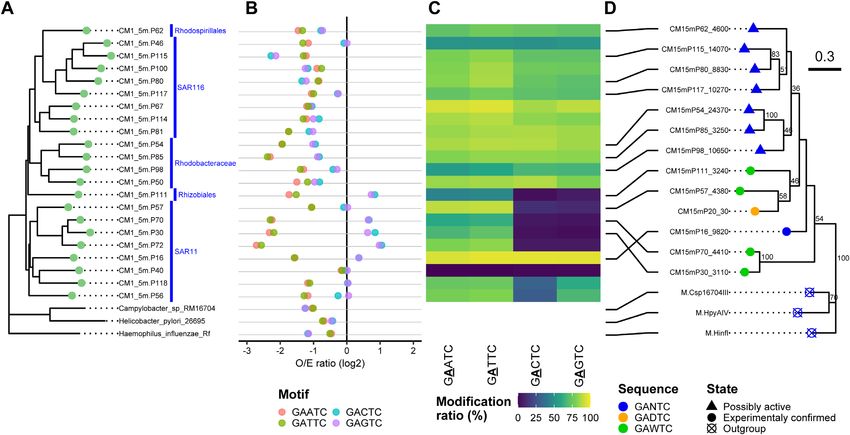
Nucleic Acids Research, 2022 3
ments and directly suspended in a cell lysis solution pro- reads were polished using both HiFi and Illumina short
vided with the kit. SMRT sequencing was conducted us- reads and HyPo (48). For the polishing, HiFi and Illu-
ing a PacBio Sequel system (Pacific Biosciences of Califor- mina reads were mapped on the pre-polished contigs us-
nia, Menlo Park, California, USA) at the National Insti- ing pbmm2, an official wrapper software for minimap2 (49)
tute of Genetics (NIG), Japan. SMRT libraries for HiFi with CCS reads settings, and Bowtie2 (50) with ‘-N 1’ set-
read via CCS mode were prepared with a 5 kb insertion ting, respectively.
length. Briefly, 4–6 kb DNA fragments from each genomic The assembled contigs were binned using MetaBAT (51)
DNA sample were extracted using the BluePippin DNA based on genome coverage and tetra-nucleotide frequencies,
size selection system (Sage Science, Beverly, Massachusetts, as genomic signatures. The genome coverage was calculated
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
USA). The SMRT sequencing library of CM1 5m and the with Illumina reads using Bowtie2 with ‘-N 1’ setting. The
other three samples were prepared using the SMRTbell quality of bins was assessed using CheckM (52), which esti-
Template Prep Kit 1.0-SPv3 and SMRTbell Express Tem- mates completeness and contamination based on the taxo-
plate Prep Kit 2.0, respectively, according to the manufac- nomic collocation of prokaryotic marker genes with default
turer’s protocol (Pacific Biosciences of California). The final settings. Bins with 99% average base-call ity assessment of the retrieved contigs and removal of flank-
accuracy were retained as HiFi reads using the standard ing host regions from integrated proviruses was performed
PacBio SMRT software package with default settings. using CheckV (59). Contigs assigned to either ‘Complete’
Metagenomic HiFi read coverage was estimated using Non- or ‘High-quality’ or ‘Medium-quality’ were defined as viral
pareil3 with default settings (37). For taxonomic assign- MAGs (V-MAGs) and used for further analysis. Taxonomy
ment of HiFi reads, Kaiju (38) in Greedy-5 mode (‘-a gradations lower than the kingdom level were estimated us-
greedy -e 5’ setting) with NCBI nr (39) and GORG-Tropics ing CAT (54). CDSs were predicted using Prodigal (45) in
databases (40) were used. HiFi reads potentially encoding an anonymous mode (‘-p meta’ setting). Functional anno-
16S ribosomal RNA (rRNA) genes were extracted using tations were achieved in the same way as for the P-MAGs.
SortMeRNA (41) with default settings, and full-length 16S
rRNA gene sequences were then predicted using RNAm-
Bioinformatic analysis of modification systems
mer (42) with default settings. The 16S rRNA gene se-
quences were taxonomically assigned using BLASTN (43) DNA modification detection and motif analysis were per-
against the SILVA database release 128 (44), wherein the formed in each MAG independently according to the of-
top-hit sequences with e-values ≤1E-15 were retrieved. ficially provided tool SMRT Link v8.0. Briefly, subreads
Coding sequences (CDSs) with >33 aa length in HiFi reads were mapped to the assembled contigs using pbmm2, and
were predicted using Prodigal (45) in anonymous mode (‘-p the interpulse duration ratios were calculated. Candidate
meta’ setting). For Illumina read data, both ends of reads motifs with scores higher than the default threshold val-
that contained low-quality bases (Phred quality score < 20) ues were retrieved as modified motifs. Those with infre-
and adapter sequences were trimmed using TrimGalore quent occurrences (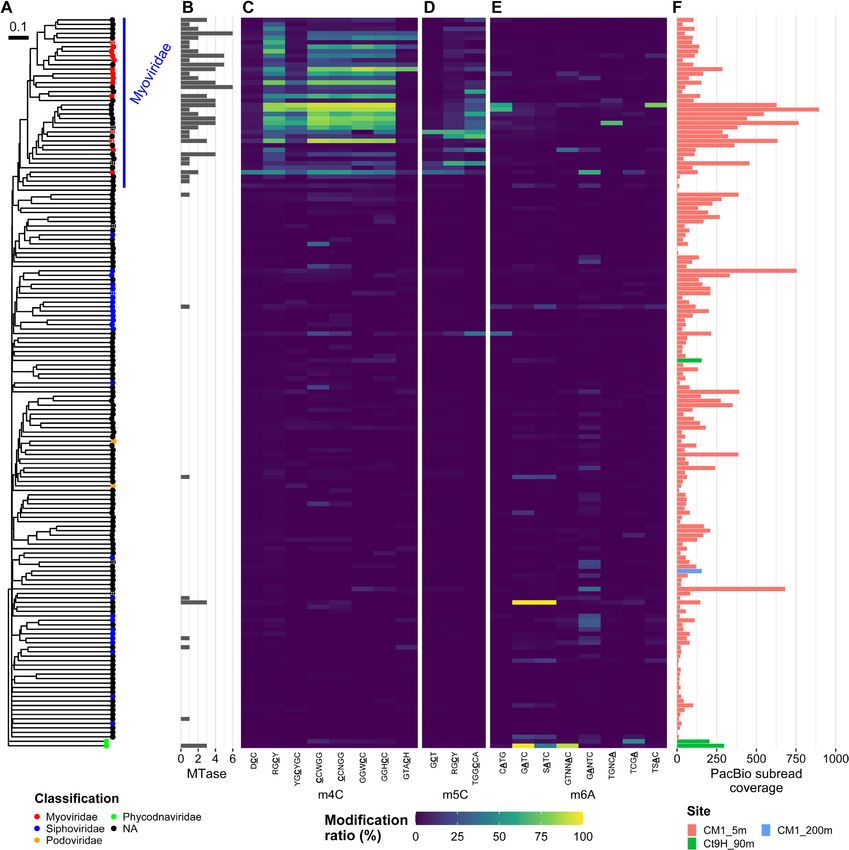
4 Nucleic Acids Research, 2022
dromic GGWCC. Further, the spurious partial sequences of Rickettsia felis URRWXCal2, Rhodospirillum rubrum
of former HBNNNNNNV and latter NH were likely due ATCC11170, Rickettsia bellii RML369-C, and Acidiphilium
to incomplete detection of the motif. Notably, we frequently cryptum JF-5 were retrieved from the NCBI database and
found candidate motifs showing this type of ambiguity in V- used as outgroups. Phylogenetic trees were constructed us-
MAGs. This possibly results from the method’s weak motif ing PhyloPhlan3 with ‘–force nucleotides –diversity low –
estimation power for small genomes. It may also reflect the accurate’ settings. Subclades of the SAR11 P-MAGs were
possibility that low presence of motifs in the genomes neg- inferred based on the topology of the phylogenetic tree, in
atively affected the motif-finding algorithm implemented in accordance with a previous definition (71–73).
MotifMaker, a tool based on progressive testing for seeking
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
longer motif sequences using a branch-and-bound search.
Genes encoding DNA methyltransferases (MTases), re- Experimental verification of MTase activities
striction endonucleases (REases), and DNA sequence- To verify MTase specificity, selected MTase genes were ar-
recognition proteins (S subunits) were searched using tificially synthesized with codon optimization by Thermo
BLASTP (43). They were compared against an experimen- Fisher Scientific. The genes were cloned into the pCold
tally confirmed gold-standard dataset from REBASE (60) III expression vector (Takara Bio, Shiga, Japan) using the
(downloaded on 9 February 2021), with a cutoff e-value In-Fusion HD Cloning Kit (Takara Bio). Additional spe-
of ≤1E-5. The sequence specificity information for each cific sequences were inserted downstream of the termi-
MTase and REase gene was retrieved from REBASE. Pairs nation codon for the methylation assay if an appropri-
of MTase and REase genes in the same genome were exam- ate sequence was absent from the plasmid vector. The
ined to determine whether they possessed the same speci- constructs were transformed into Escherichia coli HST04
ficity and constituted potential RM systems. The BREX dam− /dcm− (Takara Bio), which lacks the dam and dcm
(61) and DISARM (62) systems were sought based on Pfam MTase genes. In addition, constructs of Ct9H90mP5 10800
domains. and Ct9H90mP30 5500 were alternatively induced into the
For accurate analysis of methylome diversity, P-MAGs pET-47b(+) expression vector (Merck KGaA) using the
with >20% completeness were used for the phylogenetic In-Fusion HD Cloning Kit, and transformed into E. coli
analysis. A maximum-likelihood (ML) tree of the MAGs BL21 Star (DE3) (Thermo Fisher Scientific), owing to se-
was constructed using PhyloPhlan3 (63) on the basis of a vere insolubilization of the expressed protein in the former
set of 400 conserved prokaryotic marker genes (64) with manner. The soluble protein concentrations were measured
‘–force nucleotides –diversity high –accurate’ settings. The via SDS-PAGE as needed. E. coli strains were cultured
proteomic tree of V-MAGs was estimated using ViPTree- in LB broth supplemented with ampicillin or kanamycin.
Gen (65) with default settings. MTase expression was induced according to the supplier’s
To construct a robust phylogenetic tree of Alphapro- protocol for the expression vector. Plasmid DNA was iso-
teobacteria P-MAGs, those with higher quality (>25% lated using the FastGene Plasmid Mini Kit (Nippon Ge-
completeness) were retrieved and used for ML tree recon- netics, Tokyo, Japan) or NucleoSpin Plasmid EasyPure Kit
struction using PhyloPhlan3 with ‘–force nucleotides (Takara Bio). REase NdeI was employed for linearization
–diversity low –accurate’ settings. To calculate the of plasmid DNA. Methylation status was assayed simul-
expected/observed (E/O) ratio of each motif sequence, taneously with linearizing digestion using the appropriate
the expected and observed counts of its presentation on REases. All REases were purchased from New England Bi-
the genome were computed using R’MES (66) and SeqKit oLabs (NEB) (Ipswich, Massachusetts, USA). All digestion
(67), respectively. An ML tree of MTases was constructed reactions were performed at 37◦ C for 1 h, except for the si-
using MEGA X (68) with LG substitution model with a multaneous digestion of HinfI and TfiI at 37◦ C for 30 min,
gamma distribution (LG+G), which was selected based followed by 65◦ C for 30 min. DNA fragments were sepa-
on the Bayesian information criterion (BIC) and 100 rated via capillary electrophoresis using 5300 Fragment An-
bootstrap replicates. Three pairs of the Proteobacteria alyzer System (Agilent Technologies, Santa Clara, Califor-
genome and MTase homolog genes were retrieved from nia, USA) and the HS Genomic DNA Kit (Agilent Tech-
the NCBI database and REBASE, respectively, and used nologies).
for the following outgroups: pairs of Campylobacter sp. We further verified MTases with novel motif speci-
RM16704 and M.Csp16704III, Haemophilus influenzae Rd ficities (i.e., Ct9H300mP26 1870, Ct9H90mP5 10800,
KW20 and M.HinfI, and Helicobacter pylori 26695 and CM1200mP2 32760, CM15mP129 7780,
M.HpyAIV. Multiple sequence alignment was performed CM1200mP10 13750 and CM15mP20 30) via SMRT
using the MTase sequences, in addition to M.CcrMI from sequencing. The chromosomal DNA of E. coli HST04
Caulobacter crescentus CB15, using Clustal Omega (69). dam− /dcm− strains in which target MTases were trans-
For phylogenetic tree analysis of Alphaproteobacteria formed was extracted using the DNeasy UltraClean
and SAR11 genomes, a total of 112 and 195 deposited Microbial Kit (QIAGEN), according to the supplier’s
genomes, described by Muñoz-Gómez et al. (70) and protocol, after induction of gene expression. Multiplex
Haro-Moreno et al. (71), were retrieved from the NCBI SMRT sequencing was conducted using PacBio Sequel
database, respectively. For the analysis of Alphaproteobac- II (Pacific Biosciences of California) according to the
teria, four Betaproteobacteria and four Gammaproteobac- manufacturer’s standard protocols. Briefly, 12–50 kb DNA
teria genomes were retrieved from the NCBI database and fragments from each genomic DNA sample were extracted
used as outgroups. For the analysis of SAR11, genomes using the BluePippin size selection system (Sage Science)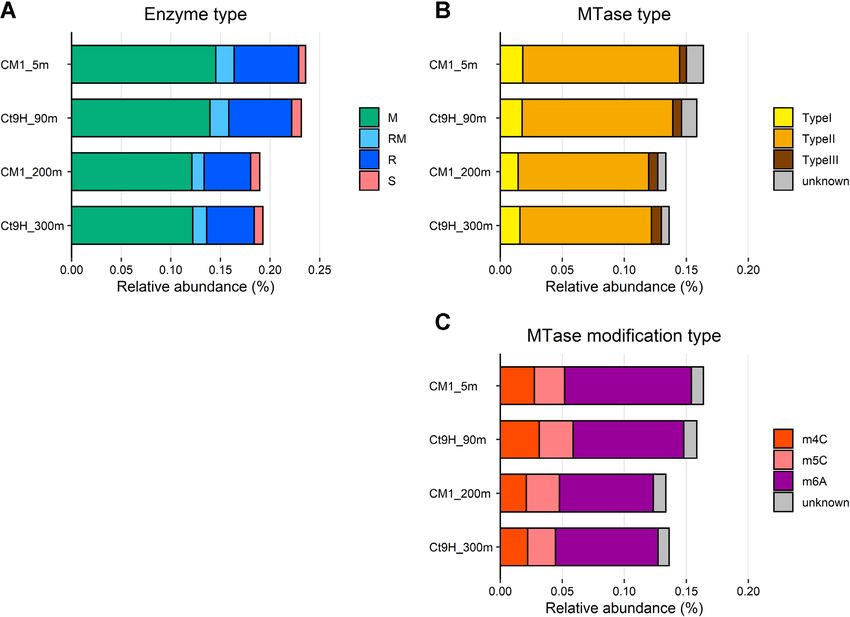
Nucleic Acids Research, 2022 5
for continuous long read (CLR) sequencing. SMRT se- was varied from 5 to 40◦ C. To investigate star activity,
quencing libraries were prepared using SMRTbell Express MTase and glycerol concentrations were varied between 1
Template Prep Kit 2.0 and Barcoded Overhang Adapter and 15 M and 0–10% v/v, respectively, and the reaction
Kit 8A according to the manufacturer’s protocol (Pacific time was extended to 3 h. The reactions were initiated with
Biosciences of California). All final SMRT libraries were 160 M SAM (NEB) in solution and terminated by adding
sequenced using Sequel II SMRT Cell 8M. Methylated mo- guanidinium thiocyanate solution buffer NTI (Takara
tifs were detected using SMRT Link v9.0 against the E. coli Bio). After the methylation reaction and following DNA
K-12 MG1655 reference genome (RefSeq NC 000913.2). purification, the methylation status was assayed using
For the in vitro assay of CM15mP111 3240 MTase and HinfI digestion at 37◦ C for 30 min.
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
its point mutant, recombinant proteins were purified. The
N-terminal 6 × His-tag fusion MTase and D49G mutant
were constructed using PCR and cloned into the pCold III RESULTS
expression vector. E. coli cells (HST04 dam− /dcm− ), trans-
formed with the constructs, were grown at 37◦ C for 16 h Shotgun sequencing and HiFi read analysis
in 20 ml of medium A (LB medium containing 50 g/ml Four seawater samples were collected from the epipelagic (5
ampicillin) via shaking. The culture was inoculated into 2 L and 90 mbsl) and mesopelagic (200 and 300 mbsl) (referred
of medium A in a 5 L flask, incubated at 37◦ C for 2–3 h with to as CM1 5m, Ct9H 90m, CM1 200m and Ct9H 300m,
constant shaking, and allowed to grow until the optical den- respectively) layers of two closely positioned stations in
sity at 600 nm reached 0.6. Then, MTase expression was in- the Pacific Ocean (Supplementary Note S1, Supplemen-
duced with 0.1 mM isopropyl -D-1-thiogalactopyranoside tary Figure S1, Supplementary Table S1). For each sample,
(IPTG) at 15◦ C, and the cultures were subsequently incu- PacBio Sequel produced 0.66–1.1 million (3.2–4.8 Gb) HiFi
bated for 16 h according to the manufacturer’s standard reads with >99% accuracy with the average length range
protocol of pCold. E. coli cells were lysed via sonication 4311–4926 bp (Supplementary Figure S2A–D, Supplemen-
in Buffer A [20 mM HEPES-Na (pH 7.5), 150 mM NaCl, tary Table S2). The HiFi reads were estimated to cover 42–
5% glycerol, 1 mM DTT, and 50 mM imidazole]. The cell 63% of the community diversity per sample (Supplementary
lysate was centrifuged at 12,000 rpm and 4 ◦ C for 30 min Figure S3). In addition to SMRT sequencing, we conducted
and then passed through a GD/X syringe filter with a 0.45 shotgun sequencing of CM1 5m using GridION and ob-
m pore size (Cytiva, Marlborough, Massachusetts, USA). tained 25 million (67 Gb) ONT reads (Supplementary Ta-
The supernatant was subjected to two-column chromatog- ble S2). Notably, SMRT and ONT sequencing each requires
raphy using the ÄKTA prime chromatography system (Cy- >10 g of DNA as initial input for library preparation,
tiva). The presence of the desired protein was confirmed via which limited our ONT sequencing to only a sample from
SDS-PAGE. Thereafter, the sample was loaded onto a 5- the surface layer (CM1 5m). The average ONT read length
ml HisTrap HP column (Cytiva) at a flow rate of 2 ml/min. (2734 ± 2013 bp) was shorter than the HiFi reads with high
The column was then washed with Buffer A. The His- deviation, likely because of the methods used for DNA ex-
tagged protein was eluted with Buffer B [20 mM HEPES- traction based on bead-beating technique, although N50
Na (pH 7.5), 150 mM NaCl, 5% glycerol, 1 mM DTT and reached 3.5 kb, and the longest read achieved was 200 kb in
300 mM imidazole]. The eluted fractions were pooled and length (Supplementary Figure S2E). Illumina MiSeq reads
diluted 5-fold with Buffer C [20 mM HEPES-Na [pH 7.5], were also obtained for each sample (Supplementary Table
150 mM NaCl and 1 mM DTT]. The diluted solution was S2). The taxonomic assignment of the HiFi reads was con-
concentrated to approximately 5 ml using a 30 kDa molec- sistent with those of previous studies, suggesting that the
ular weight cutoff Amicon Ultra centrifugal filters (Merck HiFi reads reflected general pelagic microbial communities
KGaA). It was then passed through a Millex-GP syringe with small sequencing biases (Supplementary Note S2, Sup-
filter with 0.22 m pore size (Merck KGaA), and loaded plementary Figure S4).
onto a HiLoad 16/600 Superdex 200 pg column (Cytiva) The number of genes related to DNA methylation and
pre-equilibrated with Buffer C. The protein was collected RM systems in the samples was determined by system-
as a single peak and concentrated to 2.5 mg/ml (∼50 M atic annotation of MTase and REase genes on the HiFi
in monomer concentration). It was then aliquoted, flash- reads, using the REBASE Gold Standard database (60).
frozen in liquid nitrogen, and stored at −80◦ C, or In general, genes assigned to MTase (M), REase (R), pro-
preserved with 50% glycerol at −30◦ C until further tein fused with the MTase and REase domains (RM), and
use. DNA sequence-recognition protein used in the Type I RM
The purified MTases were used for enzymatic methy- systems (S) showed similar compositions among the mi-
lation. The substrate unmethylated DNA was pro- crobial communities (Figure 1A). Within the MTase pro-
duced via PCR, with the pCold III vector transferred teins (i.e., M and RM), Type II was predominant, account-
CM15mP111 3240 gene as a template to match with the ing for 76.5–78.6% of each sample (Figure 1B). The rela-
in vivo assay of the MTase. Methylation reactions were tive abundances of Type I (11.2–12.1%) and III (3.4–5.9%)
carried out in a reaction buffer [20 mM HEPES-Na pH were approximately 2–3 times lower than those identified
7.5, 100 mM NaCl and 100 g/ml BSA] containing 5 nM in the genomes of prokaryotic isolates, reported as 27%
substrate DNA and 1 M purified MTase, at 20◦ C for 1 and 8%, respectively (23). Among the detected MTases, the
h, unless specified otherwise. To investigate salt sensitivity, most abundant predicted modification type was m6A (56.9–
NaCl concentration was varied from 0 to 400 mM. To 62.9%), followed by m4C (15.6–19.6%) and m5C (14.6–
investigate thermal sensitivity, the reaction temperature 19.6%) (Figure 1C).6 Nucleic Acids Research, 2022
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
Figure 1. Relative abundance of genes encoding DNA restriction and modification enzymes in marine pelagic metagenomes. CDSs predicted from HiFi
reads were used in this analysis. (A) Distribution of enzyme types: DNA methyltransferase (MTase; M), Restriction endonuclease (REase; R), protein
fused with M and R domains (RM), and DNA sequence-recognition protein (S). (B) Distribution of MTase types. (C) Distribution of modification types.
Diverse DNA modifications in metagenomic assembled with 30% on average), similar to previous observations of
genomes phosphorothioated motifs in E. coli (12%) and Thaumar-
chaeota (20%) strains (25,26).
After HiFi and ONT read assembly and binning analyses,
Among the P-MAGs with methylated motifs, GATC
we obtained 233 and 163 prokaryotic MAGs (P-MAGs)
was detected most frequently (41 P-MAGs), followed by
and viral MAGs (V-MAGs), respectively (Supplementary
GANTC (28 P-MAGs), CGCG (19 P-MAGs) and BAAAA
Note S3, Supplementary Figure S5, Supplementary Ta-
(9 P-MAGs), where B = C/G/T and N = A/C/G/T, and
ble S3, Supplementary Data S1). From the reconstructed
the underlined characters indicate modified bases. Among
MAGs, a total of 178 and 42 candidate modified motifs
the V-MAGs, RGCY (9 V-MAGs) was the most abundant
were detected in 108 (46%) P-MAGs and 15 (9%) V-MAGs,
motif, followed by CCNGG (4 V-MAGs), GGWCC (3 V-
respectively (Supplementary Data S2). Mapped subread
MAGs) and GGHCC (3 V-MAGs), where R = A/G, Y =
coverages of the modified motifs were compatible with P-
C/T, W = A/T and H = A/C/T. Notably, even consider-
MAGs and V-MAGs that ranged from 30.6 to 508.9 × and
ing some vague motifs, at least 15 motifs (i.e., BAAAA,
88.3 to 568.8 ×, respectively.
ACAAA, CAAAT, CTAG, GATGG, GATCC, GTNAC,
The detected motifs were composed of 59 unique mo-
GTWAC, SATC, TGNCA, TSAC, CTCC [m4C], GCGC
tifs, including 32 motifs with palindromic sequences that al-
[m4C], GGWCC [m4C] and TGGCCA [m5C], where S =
low double-strand modification. Among the said motifs, 27
C/G) did not match the known MTase motifs in the RE-
and 23 were classified as m6A and m4C methylation types,
BASE repository. In addition, methylated motifs likely cat-
respectively. Although current SMRT sequencing technol-
alyzed by Type I MTases, which are generally characterized
ogy does not support detection of the m5C motif, we found
as bipartite sequences with a gap of unspecified nucleotides
four candidate m5C motifs with high subread mean cov-
(e.g., ATGNNNNNTAC), were undetected in all P-MAGs
erage (259 × on average). Among the methylated motifs
and V-MAGs. This result indicates that Type I RM sys-
from P-MAGs, 57 (35%) showedNucleic Acids Research, 2022 7
Prediction of MTases and corresponding methylated motifs was found in 30 (13%) P-MAGs, but no methylated mo-
tifs were detected. Among the 30 P-MAGs, 42 candidate
To identify MTases that catalyze methylation of the de-
MTase genes, comprising 1 m4C-type, 6 m5C-type, 27 m6A-
tected motifs, systematic annotation of MTase genes was
type, and 8 type-unknown MTase genes were found. We an-
performed. Sequence similarity searches against known
ticipate that either the MTase genes were inactive, or the
genes stored in REBASE (60) identified 171, 43, and 7
corresponding methylated motif went undetected owing to
of M, R, and RM genes, respectively, from 112 (48%)
the low sensitivity of SMRT sequencing, especially for m5C
P-MAGs (sequence identity in the range 20–92%) (Sup-
modification (22,23).
plementary Figure S6, Supplementary Data S3). M genes
Among the viral genomes, 82, 13 and 16 of the M, R and
tended to be more frequently detected than R genes in each
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
RM genes were identified in 49 (30%) V-MAGs (sequence
P-MAG (Supplementary Figure S7A). Only three S genes
identity in the range 23–73%) (Supplementary Figure S6,
were found in the P-MAGs, and the small number was
Supplementary Data S3). Similar to the case of P-MAGs, M
concordant with the results of HiFi read analysis (Figure
genes tended to be more frequently detected than R genes
1A). Among the M and RM genes from P-MAGs, m6A
in each V-MAG. Type II MTases were the most abundant
(64%) was the most abundant modification type, followed
(79%), followed by Type I (7%), with no Type III detected
by m4C (14%) and m5C (10%), as found in the HiFi read
at all (Supplementary Figure S6, S7B). In contrast to that
analysis (Figure 1C). Among the MTase types, Type II
in P-MAGs, m4C (62%) was the most abundant modifica-
MTases were the most abundant (82%), and 9% and 6%
tion type in V-MAGs, followed by m6A (30%) and m5C
of genes showed the highest sequence similarity to Type
(1%). All MTases and methylated motifs were unmatched
I and III MTases, respectively. This trend was consistent
in V-MAGs, except for three pairs (GATC in CM1 5m.V34,
with the HiFi read analysis result in which Type I and III
GATC, and GTNNAC in Ct9H 90m.V1). This may reflect
MTases were scarcely detected in the communities (Figure
the very low number of viral MTases stored in the REBASE
1B). Most of the MTases were orphan, and only four pairs
Gold Standard database, where 16 viral MTase genes were
of Type II MTase and REase genes were predicted to pos-
found out of a total of 1938 MTase genes.
sess the same motif sequence specificity and be adjusted
on the genome, which may constitute intact Type II RM
systems. Other known antiviral defense systems associated Exploration and experimental verification of MTases with
with DNA modification, BREX (61) and DISARM (62), new specificity
were surveyed. However, no MTase genes likely associated Among the detected MTase genes, 132 (74%) and 94 (96%)
with these systems were found in the P-MAGs. Moreover, MTases from P-MAGs and V-MAGs, respectively, showed
neither the number of modified motifs nor MTase genes inconsistency between the recognition motifs of their clos-
showed a clear association with the number of CRISPR est relatives and the methylated motifs identified in our
arrays in the P-MAGs (Supplementary Data S1). Overall, metaepigenomic analysis (Supplementary Data S2 and S3).
these analyses highlight the previously unknown diverse This result suggested that the homology-based estimation
MTases in epipelagic and mesopelagic prokaryotic commu- of MTase specificity was not sufficient, as in our previous
nities and suggest that methylation systems play unexplored metaepigenomic study of the freshwater microbiome (32).
roles apart from their known role in the defense mechanisms To reveal the catalytic specificity of these MTases, we se-
against exogenous DNA. lected potential pairs of MTase and methylated motifs as
A total of 58 (20%) MTase genes in P-MAGs showed the follows: (i) MTase and methylated motifs were present in the
best sequence similarity to MTases, whose specificity was same genome, and novel correspondence was predicted, (ii)
exactly matched to the motif identified in our metaepige- modification types (i.e., m4C, m5C and m6A) of MTase and
nomic analysis (Supplementary Data S2 and S3). For exam- methylated motifs were concordant and (iii) the complete
ple, CM1 200m.P15 contained one MTase that showed the sequence of the MTase gene was retrieved. Subsequently,
best sequence similarity to those that recognized CCSGG, the methylation specificities of selected MTases were ex-
and this finding was perfectly congruent with the motif de- perimentally verified by heterologous expression in E. coli.
tected in the P-MAG. For CM1 200m.P39, two MTases Briefly, plasmids with one artificially synthesized MTase
similar to those that recognize either TTAA or CGCG were gene were constructed and transformed into E. coli cells,
identified, and these motifs were congruently detected in the and the methylation status of the isolated plasmid DNA was
genome. In Ct9H 300m.P17, five MTases were predicted, subsequently observed using REase digestion after heterol-
two of which were similar to the known MTases that recog- ogous expression. The artificially synthesized sequences are
nize either AGCT or GATC. All of the detected methylated summarized in Supplementary Data S4.
motifs in the genome completely matched with them, sug- In Actinobacteria, Ct9H 300m.P26, one orphan m6A
gesting that the two MTases were active. MTase gene, and two m6A and m4C motifs were detected.
At least one methylated motif was detected in 44 (19%) However, none of the MTase and motif matched with each
P-MAGs, whereas no MTase gene was found. We assumed other. Thus, we predicted that Ct9H300mP26 1870, whose
that the corresponding MTase genes were missed because closest homolog encoded an MTase that exhibits CTCGAG
of insufficient completeness of genomes (including chro- methylation activity, would encode an MTase that recog-
mosome, plasmids, or multi-partite genomes such as chro- nizes BAAAA, whereas the motif sequence was not regis-
mid and megaplasmid) in the binning process. Alternatively, tered in REBASE and no MTase is currently reported to
these MTase genes may have diverged considerably from recognize the motif. The REase digestion assay result was
known MTase genes. In contrast, at least one MTase gene consistent with the hypothesis that ScaI (AGTACT speci-8 Nucleic Acids Research, 2022
ficity) did not cleave the BAAAAGTACT sequence, which that possesses previously unreported ACAAA specificity
overlapped with BAAAA and AGTACT sequences on the (Table 1).
plasmids, only when MTase was expressed in the cells (Fig- In Candidatus (Ca.) Marinimicrobia CM1 200m.P10,
ure 2A). We named this protein M.AspCt9H300mP26I, a one orphan MTase gene, and one methylated motif were de-
novel MTase that possesses BAAAA specificity (Table 1). tected. The reported recognition motif of the closest MTase
In Actinobacteria, Ct9H 90m.P5, two orphan MTase is GAAGA (the modified base is the second position of the
genes, and three methylated motifs were detected. While complementary sequence TCTTC), while the detected mo-
a pair of MTase and motif was concordantly matched, tif was CTCC. Thus, we hypothesized that the recognition
the other MTases did not match any motifs. The lat- motif of CM1200mP10 13750 MTase would be CTCC, a
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
ter MTase gene Ct9H90mP5 10800 showed moderate se- previously unreported methylated motif. The REase diges-
quence similarity (32%) to M.AspCt9H300mP26I using tion assay showed that ScaI was inhibited from cleaving the
BLASTP search with a low e-value (1E-70), and either of GGAGTACTCC sequence site, where the ScaI-targeting
the remaining motifs was m6A and m4C. Thus, we pre- site was complementally sandwiched by CTCC (Figure 2E).
dicted that Ct9H90mP5 10800 MTase, whose closest ho- We named this protein M.MspCM1200mP10I (Table 1).
molog is an m6A MTase that exhibits ATTAAT methy- Furthermore, we conducted a re-sequencing analysis to
lation, would have BAAAA specificity. As expected, the examine the methylation status of the chromosomal DNA
REase digestion assay showed that ScaI resisted cleav- of E. coli in which each novel MTase gene was transformed
ing the BAAAAGTACT sequence only when the pro- and expressed. As a result, all five methylated motifs were
tein was expressed (Figure 2B). We named this pro- successfully recalled in each of the E. coli genomes (Supple-
tein M.AspCt9H90mP5I, a novel MTase that possesses mentary Table S4).
BAAAA specificity (Table 1). Notably, another candi-
date orphan MTase gene, Ct9H90mP30 5500, was de-
tected in Actinobacteria Ct9H 90m.P3. It was predicted
Phylogenetic distribution of modified motifs
to possess the same BAAAA specificity and showed
moderate (33%) and high (87%) sequence similarity to To investigate the phylogenetic distribution of the DNA
M.AspCt9H300mP26I and M.AspCt9H90mP5I, respec- modification system in the MAGs, we used 117 P-MAGs
tively. However, this protein was insoluble in E. coli, result- (>20% completeness) and all 163 V-MAGs for robust phy-
ing in no clear cleavage inhibition in our experiment. logenetic tree reconstruction, and visualized the modifica-
A Planctomycetes CM1 200m.P2 had three orphan tion ratios of the detected motifs in each genome (Figure
MTase genes and two methylated motifs. One of the 3). Within the P-MAGs, modified motifs were sporadically
MTases showed the highest sequence similarity to those distributed across the phyla, whereas some showed great
recognizing TTAA with high similarity (64%). The other concordance with the phylogenetic clades. For example,
CM1200mP2 32760 and CM1200mP2 5150 MTases within the phylum Actinobacteria, CGCG and BAAAA
showed the highest sequence similarity to those catalyz- were spread in the genomes of all organisms from the class
ing m6A modification and recognizing GTTAAC and Acidimicrobiia but were not detected in organisms from
ATTAAT, respectively, with low similarity (37% and 25%, the class Actinobacteria. By contrast, AATT was found in
respectively). The two detected motifs were GCGC (m4C) three P-MAGs belonging to a subclade in Acidimicrobiia.
and CAAAT (m6A), the latter of which was not found in TTAA was detected in four P-MAGs in Chloroflexi. GATC
REBASE. Thus, we expected that either or both MTases was detected with moderate to high modification ratios (19–
would recognize and methylate the novel CAAAT motif. 99%) through archaeal P-MAGs, with two exceptions; no
The construct CM1200mP2 32760 was not successfully significant GATC signature was detected in Euryarchaeota
prepared in our experiment, likely because the protein Ct9H 90m.P24 (7%) and CM1 5m.P82 (0.4%) possibly due
was toxic to E. coli. In contrast, CM1200mP2 5150 to the methylation activity being weak or absent in these
MTase showed that MluCI (AATT specificity) did not organisms. AGCT was observed in both the Thaumar-
cleave all CAAATT sequences when only MTase was chaeota P-MAGs with high modification ratios (82–91%).
expressed. This clearly indicated that MTase recognizes CGCG was found in members from three phyla across the
CAAAT (Figure 2C). Accordingly, we named the pro- domain: Actinobacteria, Chloroflexi and Euryarchaeota.
tein M.PspCM1200mP2I, a novel MTase that possesses GANTC/GAWTC appeared in all 26 Alphaproteobacteria
previously unknown CAAAT specificity (Table 1). P-MAGs, with only one exception. In addition to methyla-
Chloroflexi CM1 5m.P129 had one orphan MTase gene, tion, AGCT modified motif showed weak modification ra-
which showed the highest sequence similarity to those rec- tios (2–19%) in the class Ca. Poseidoniia P-MAGs. How-
ognizing TCTAGA (whose modification type and position ever, this motif was detected only in Ct9H 300m.P10 in
were not reported). However, the only methylated motif the motif prediction analysis. This result demonstrates that
detected in the genome was ACAAA, which no MTase phylogeny-based modification ratio analysis is efficient for
was currently reported to recognize. Thus, we hypothesized analyzing infrequently modified motifs.
that CM15mP129 7780 MTase should recognize and mod- By sharp contrast, many motifs showed no clear as-
ify this novel motif. The REase digestion assay result was sociation with the phylogenetic topology. For example,
consistent with the hypothesis that BceAI (ACGGC speci- GCWGC was solitary, with a high modification ratio in
ficity) did not cleave the ACAAACGCG sequence when Chloroflexi CM1 200m.P6. Similarly, CTAG in Ca. Marin-
only MTase was expressed (Figure 2D). Accordingly, we imicrobia Ct9H 300m.P2, CTCC in Ca. Marinimicrobia
named this protein M.CspCM15mP129I, a novel MTase CM1 200m.P10, GTAC in Euryarchaeota CM1 5m.P3,Nucleic Acids Research, 2022 9
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
Figure 2. REase digestion assays of MTases with novel specificity. (A) Assay of the Ct9H300mP26 1870 gene. ScaI was used, where the plasmid contained
two AGTACT target sites. Within the two sites, one of the target sites was BAAAAGTACT, where overlapped BAAAA and AGTACT were recognized by
the MTase and REase. (B) Assay of the Ct9H90mP5 10800 gene. ScaI was used, where the plasmid contained one AGTACT target site in the BAAAAG-
TACT site. (C) Assay of the CM1200mP2 32760 gene. MluCI were used, where the plasmid contained 23 AATT target sites. Within them, the six target
sites were CAAATT, where overlapped CAAAT and AATT were recognized by the MTase and REase, respectively. (D) Assay of the CM15mP129 7780
gene. BceI were used, where the plasmid contained six ACGGC target sites. Within them, one of the target sites was ACAAACGGC, where overlapped
ACAAA and ACGGC were recognized by the MTase and REase, respectively. (E) Assay of the CM1200mP10 13750 gene. ScaI were used, where the
plasmid contained two TCATGA target sites. Within them, one of the target sites was GGAGTACTCC, where a pair of CTCC and GGAG (comprehen-
sive sequence of CTCC) and TCATGA were recognized by the MTase and REase, respectively. The pCold III (A, C–E) and pET-47b(+) (E) were used as
expression vectors. The schematic representation and plasmid map are presented on the right side. The underlined characters indicate modified bases. The
orange arrow represents the transferred gene, and the red framed digestion sites represent the location of the overlapped sequence. The band sizes were
expected to emerge (red triangles) and disappear (blue triangles) when the induced MTase caused methylation. All plasmid DNAs were linearized using
NdeI.Table 1. Novel MTases whose specificities were experimentally confirmed. The underlined characters indicate modified bases.
Recognition
Top-hit motif of the Confirmed
10 Nucleic Acids Research, 2022
protein in Identity RM closest-match Modification Modification RM recognition Novel
Gene ID Genome ID Lineage REBASE (%) type MTase position type system MTase name motif specificity
CM15mP129 7780 CM1 5m.P129 Bacteria; Chloroflexi; NA; NA; NA; NA; M.Spn6BI 35.5 II TCTAGA (unknown) (unknown) No M.CspCM15mP129I ACAAA Yes
NA
Ct9H90mP5 10800 Ct9H 90m.P5 Bacteria; Actinobacteria; Acidimicrobiia; M.Sgl8271II 26.8 II ATTAAT 5 m6A No M.AspCt9H90mP5I BAAAA Yes
Acidimicrobiales; NA; NA; NA
CM1200mP2 5150 CM1 200m.P2 Bacteria; Planctomycetes; Planctomycetia; M.Sgl8271II 24.5 II ATTAAT 5 m6A No M.PspCM1200mP2I CAAAT Yes
Planctomycetales; Planctomycetaceae; NA;
Planctomycetaceae bacterium
Bacteria; Candidatus Marinimicrobia; NA; M2.HpyAII 44.2 II GAAGA -2 m4C No CTCC Yes
CM1200mP10 13750 CM1 200m.P10 NA; NA; NA; Candidatus Marinimicrobia M.MspCM1200mP10I
bacterium
Bacteria; Actinobacteria; Acidimicrobiia; M.TliI 25.0 II CTCGAG 5 m6A No BAAAA Yes
Ct9H300mP26 1870 Ct9H 300m.P26 Acidimicrobiales; NA; NA; NA M.AspCt9H300mP26I
CM15mP16 9820 CM1 5m.P16 Bacteria; Proteobacteria; M.CspNS6I 62.9 II GANTC 2 m6A No M.PspCM15mP16I GANTC No
Alphaproteobacteria; Pelagibacterales;
NA; NA; NA
CM15mP20 30 CM1 5m.P20 Bacteria; Proteobacteria; M.SmeI 59.9 ? GANTC 2 m6A No M.AspCM15mP20I GADTC Yes
Alphaproteobacteria; NA; NA; NA;
Alphaproteobacteria bacterium
CM15mP30 3110 CM1 5m.P30 Bacteria; Proteobacteria; 49.9 II GANTC 2 m6A No M.PspCM15mP30I GAWTC No
Alphaproteobacteria; Pelagibacterales; M.Bba35685I
Pelagibacteraceae; NA; Pelagibacteraceae
bacterium
CM15mP57 4380 CM1 5m.P57 Bacteria; Proteobacteria; M.SmeI 57.0 ? GANTC 2 m6A No M.AspCM15mP57I GAWTC No
Alphaproteobacteria; NA; NA; NA; NA
CM15mP70 4410 CM1 5m.P70 Bacteria; Proteobacteria; M.Bsp460I 48.0 II GANTC 2 m6A No M.PspCM15mP70I GAWTC No
Alphaproteobacteria; Pelagibacterales;
Pelagibacteraceae; NA;
Alphaproteobacterium
CM15mP111 3240 CM1 5m.P111 Bacteria; Proteobacteria; M.SstE37II 55.6 II GANTC 2 m6A No M.RspCM15mP111I GAWTC No
Alphaproteobacteria; Rhizobiales; NA;
NA; NA
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022Nucleic Acids Research, 2022 11
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
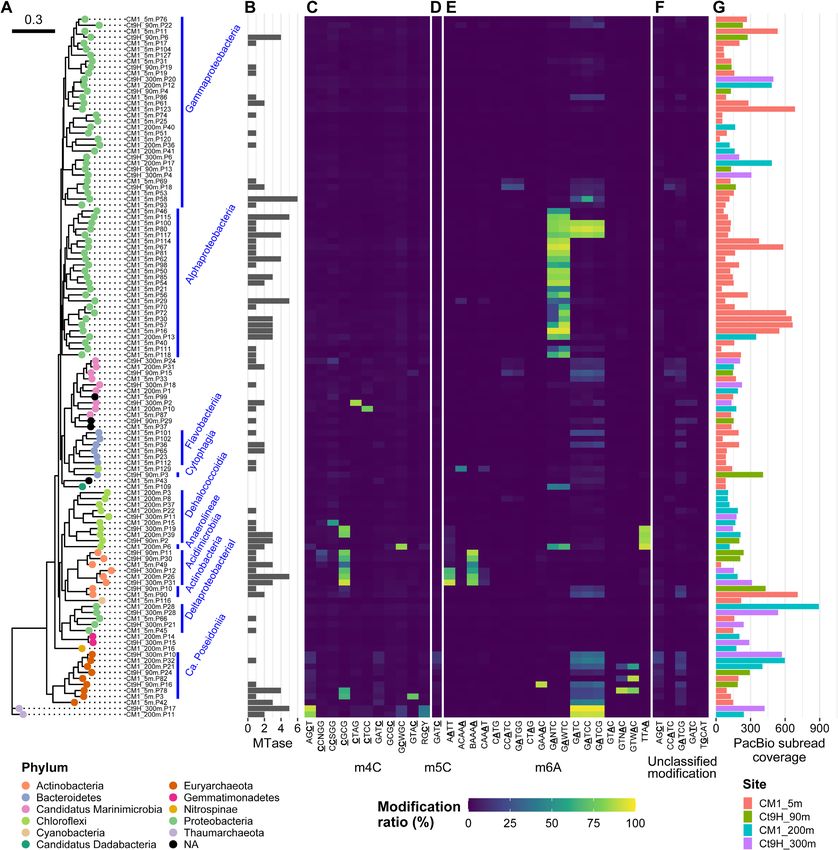
Figure 3. Methylomes of P-MAGs. (A) A phylogenetic tree was constructed based on a set of up to 400 conserved bacterial marker genes using the
maximum-likelihood method. Node color indicates taxonomy at the phylum level. Nodes were grouped at class to family levels, if estimated (blue bars and
texts). (B) Numbers of MTase genes identified in each genome. (C–F) Modification ratios of detected motifs per genome. (C) m4C, (D) m5C, (E) m6A and
(F) unclassified modifications are shown individually. Motifs detected from P-MAGs without spurious sequence were used. The color range from blue over
green to yellow represents modification ratios of motifs on each genome. The underlined characters indicate modified bases. Notably, modification ratios
were affected by overlapped motif sequences; for example, GATCC was completely overlapped by GATC, and both motifs showed similar modification
rates in their genomes except in Gammaproteobacteria CM1 5m.P58 where GATCC was detected on the genome as per the metaepigenomic analysis and
concordantly the modification ratio of GATCC was higher than that of GATC. (G) Coverages of subread on each genome. The bar color represents the
source of the genome.12 Nucleic Acids Research, 2022
ACAAA in Chloroflexi CM1 5mP129, and GAAAC in Eu- MTases that recognize GADTC/GAWTC motifs in marine
ryarchaeota Ct9H 90m.P16 were found. Alphaproteobacteria
Within all the P-MAGs in this study, no methylated motif
M.CcrMI is also known as ‘cell cycle-regulated MTase’
was detected in 125 (54%) P-MAGs with high subread cov-
(CcrM) from Caulobacter crescentus. Along with GANTC
erage (ranging from 31.4–3305.7 × and 207.6 × on average);
specificity, it is one of the model protein of prokary-
thus, this was not addressed by insufficient coverage depth
otic MTase that is well conserved in Alphaproteobacteria
for modification detection. The 125 P-MAGs were found
(23,75,76). Indeed, GANTC was previously identified in
to be dispersed across diverse phyla, such as Proteobacte-
diverse lineages of Alphaproteobacteria isolates (23,77,78)
ria, Bacteroidetes, Ca. Marinimicrobia, Chloroflexi, Gem-
and one MAG (32) using the modern SMRT sequencing
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
matimonadetes, Cyanobacteria and Verrucomicrobia. In-
technique, and no alternative motifs have been reported. In
terestingly, neither methylated motifs nor MTase genes were
our metaepigenomic analysis, GANTC was concordantly
detected in P-MAGs belonging to the following lineages:
detected in 26 of 40 Alphaproteobacteria P-MAGs (Supple-
both members of Gemmatimonadetes and Nitrospinae, and
mentary Data S3). In addition, we detected similar but dif-
all five members of Verrucomicrobia (Supplementary Data
ferent motifs GAWTC, GADTC, and GAHTC from seven,
S1). Methylated motifs were also absent from all three
four and one Alphaproteobacteria P-MAGs, respectively
Deltaproteobacteria P-MAGs, although two of them pos-
(where D = A/G/T) (Figure S8). This result strongly sug-
sessed the MTase gene. Within the Gammaproteobacteria
gests the presence of unknown variations in the methyla-
P-MAGs, 31 of the 52 genomes lacked both methylated
tion system in the lineage. We should note that because
motifs and MTase genes. Taken together, these observa-
all the detected modified bases on both DNA strands were
tions suggest the absence of a DNA methylation system in
used for motif prediction, the detection of non-palindromic
several clades. This finding contradicts that of a previous
GADTC and GAHTC motifs was not explained by the
study, which reported the pervasiveness of DNA methyla-
differences in the proportion of GACTC and GAGTC se-
tion among bacteria and archaea (23).
quences in the genomic data (represented by one side of the
Methylated motifs were occasionally detected with low
DNA strand in the fasta file). It should also be noted that
modification ratios in most V-MAGs, except for Phycod-
modifications other than m6A were rarely found on either
naviridae and Myoviridae (Figure 4). Among the Phy-
of the strands in the motif. This suggests that the motifs did
codnaviridae V-MAGs, Ct9H 90m.V1 showed GATC and
not result from the wrong prediction due to the modifica-
GTNNAC having a high modification ratio, whereas
tions other than m6A. With regard to the Alphaproteobac-
Ct9H 90m.V2 harbored TCGA. These results were con-
teria P-MAGs, we predicted 13 complete gene sequences of
sistent with previous findings, which reported that m6A is
MTase that were assumed to recognize either of the motifs.
frequently found in Phycodnaviridae genomes (74). In 14
However, all of them showed the highest sequence similar-
Myoviridae V-MAGs, 0–5 methylated motifs were detected.
ity (47–80%) to those known to recognize GANTC (Sup-
However, the proteomic tree showed numbers of V-MAGs
plementary Data S3).
that, though not taxonomically assigned, were closely re-
Considering the correspondence of the methylated mo-
lated to the Myoviridae family (referred to as ‘Myoviridae-
tifs and MTases, it was expected that four and one MTases
like’). The Myoviridae-like V-MAGs frequently appeared
would recognize GAWTC and GADTC, respectively,
to share several m4C motifs (e.g., RGCY, CCWGG,
rather than GANTC (Supplementary Data S2 and S3).
GGWCC) with different combinations. Sometimes, they
The REase digestion assay of the former four MTases
also harbored additional motifs, while a few numbers of
(CM15mP30 3110, CM15mP57 4380, CM15mP70 4410
modified motifs were detected in the motif prediction anal-
and CM15mP111 3240) showed that TfiI (GAWTC
ysis (0.95 motifs per genome on average). This indicates
specificity) cleavage was completely blocked only when
that the taxonomic assignment of the viral genome was fre-
MTase was expressed in the cells, whereas HinfI (GANTC
quently missed due to the lack of viral genomes in the refer-
specificity) partly cleaved the plasmids (Supplementary
ence database and the severe underestimation of modified
Figure S9A–D). Despite exhibiting off-target effects under
motifs in V-MAGs, likely due to their small genome size
high concentrations of the enzyme, known as ‘star activity’
(see Materials and Methods). In contrast to the Phycod-
(79,80), assays of purified CM15mP111 3240 MTase pro-
naviridae and Myoviridae-like V-MAGs, methylation was
tein suggested that it showed canonical specificity toward
scarcely detected in the other V-MAGs, including those of
GAWTC (Supplementary Note S4, Figure S10A–C).
Siphoviridae and Podoviridae. Notably, the methylated mo-
The digestion pattern in the assay of CM15mP20 30 was
tifs detected in the V-MAGs were rarely shared with those
also congruent with the hypothesized GADTC methyla-
in the P-MAGs, and no modified motifs other than methy-
tion and re-sequencing analysis successfully recalled the
lation were found. Five Myoviridae-like V-MAGs were pre-
methylated motif, thus indicating its low efficiency with
dicted to be proviruses. However, no clear difference was
regard to GACTC methylation (Supplementary Note
observed in the modification ratio compared to the case for
S5, Supplementary Figure S9E, Supplementary Table
the other non-provirus V-MAGs. In 39 Myoviridae-like V-
S4). By contrast, as expected, robust GANTC speci-
MAGs, several MTases were found to be encoded in their
ficity was confirmed in the assay of CM15mP16 9820,
genomes (ranging from 0 to 6 and 2.2 MTase genes per
which completely inhibited both TfiI and HinfI cleavage
genome on average) (Figure 4B and Supplementary Data
(Supplementary Figure S9F). Accordingly, we named
S3). Yet, they were scarcely detected in the other V-MAGs
the four (M.PspCM15mP30I, M.AspCM15mP57I,
(ranging from 0 to 3 and 0.1 on average).
M.PspCM15mP70I, and M.RspCM15mP111I) and oneNucleic Acids Research, 2022 13
Downloaded from https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkab1292/6509096 by guest on 19 February 2022
Figure 4. Methylomes of V-MAGs. (A) A proteomic tree was generated based on the global genomic similarities between viral genomes. Proviruses are
indicated by circle cross. Node color indicates taxonomy at the family level. (B) Numbers of MTase genes identified in each genome. (C–E) Modification
ratios of (C) m4C, (D) m5C and (E) m6A motifs. (F) Coverages of subread on each genome. Please also see Figure 3.
(M.AspCM15mP20I) proteins as novel MTases that Based on the sequence alignment of the 13 MTases with
preferentially recognize GAWTC and GADTC, respec- M.CcrMI and its homologs, a glycine residue (correspond-
tively, and the last protein (M.PspCM15mP16I), as one ing to Gly40 in M.CcrMI) was roughly conserved in all
that recognizes GANTC (Table 1). Interestingly, we MTases with GANTC specificity. By contrast, it was re-
found that the temperature-activity profile of the purified placed with lysine or aspartic acid in all MTases with
M.RspCM15mP111I MTase (Supplementary Figure S10B) GAWTC specificity (Supplementary Figure S11). It has
was concordant with the marine water temperature at the been reported that the M.CcrMI protein contains a sub-
sampling sites (Supplementary Figure S1B), suggesting structure that forms a pocket to accommodate the third
that MTase was thermally optimized in an epipelagic position of the recognized motif (i.e., nucleotide ‘N’ in
environment. GANTC); two hydrophobic residues Leu38–Leu42 stacks,You can also read

















































