Development of Male Endangered Species Germ Cells from Induced Pluripotent Stem Cells
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Development of Male Endangered Species Germ Cells from Induced Pluripotent
Stem Cells
I. Specific Aims
More than 25% of all mammals in the world are considered “threatened” with half of all species showing a
downward trend in population and others considered extinct outside of captivity (International Union for
Conservation of Nature (IUCN) 2008 Red List of Threatened Species). This is true despite the numerous
traditional conservation strategies that have been employed including captive breeding programs. One
significant challenge in endangered species captive breeding programs is the limited number of individuals
resulting in small gene pools and unsustainable populations. This situation is further exacerbated by the early
deaths of animals, the inability to easily move individuals between zoological parks for breeding and
incompatibility between potential mates. In this study, we introduce a novel approach to preserve genetic
diversity by generating male germ cells (sperm) from skin fibroblast cells. We will use induced pluripotent stem
cells (iPSCs), cells closely resembling embryonic stem cells that can turn (or differentiate) into any cell type in
the body including germ cells (GCs), to accomplish this goal. Our long term goal is to produce iPSC derived
sperm and/or eggs capable of being easily stored and producing offspring. Once accomplished, this system
can provide an important tool in breeding programs by:
1. Providing an infinite source of undifferentiated stem cells, as iPSCs are immortal and can be continually
expanded and easily frozen, that can be banked from living and deceased animals to preserve genetics
2. Turning endangered species stem cells into sperm and oocytes for use in captive breeding programs,
including species where these cell types are not easily collected
3. Providing an opportunity to increase gene pools by non-invasively collecting cells from wild specimens
that can be turned into sperm and oocytes and used in captive breeding programs
4. Reducing the need to transfer animals between zoos for breeding, which is costly and a potential
source of disease transfer
5. Offering a systems to study germ cell development and abnormalities in endangered species
6. Allowing for easy introduction of genes conferring resistance to common diseases that have
significantly affected species in the past
Our labs (West and Stice) have recently developed a novel and efficient method of generating relatively
homogeneous (69%) populations of
germ cells (GCs) from human
embryonic stem cells (hESCs) at
levels that have not been previously
noted [1]. Using an adherent
differentiation culture system that
allows for easy isolation and
manipulation, GCs demonstrated co-
expression of the germ cell specific
marker DDX4 and the pluripotency
marker POU5F1 at the protein level
(see preliminary data). Additionally,
GC (DDX4+ POU5F1+) enriched
cultures expressed high levels of pre-
migratory, post-migratory and meiotic
GC genes and were capable of being
continually propagated. These cells
have also demonstrated meiotic
activity with the expression of the
proteins SYCP3 and MLH1 in >90% of
GCs [1]. In addition, utilizing new and Figure 1. Work flow diagram for the production of endangered species iPSCs.
innovative techniques, we have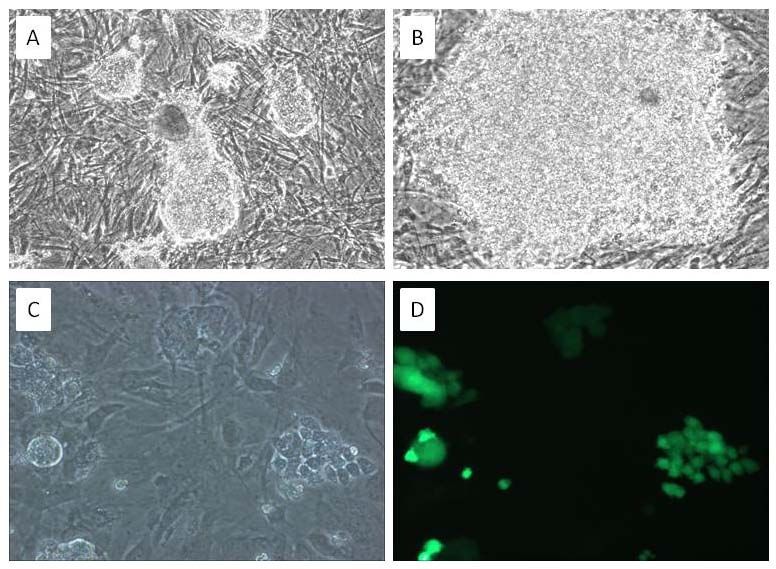
developed a system to derive iPSCs from domestic pigs (Sus scrofa domesticus). These cells possess
pluripotent properties including gene and protein expression and the ability to give rise to all 3 germ layers
(ectoderm, endoderm and mesoderm).
Here we propose to investigate the ability of fibroblast cells collected from a single male individual from six
different endangered species (Clouded Leopard, African Lion, Sumatran Tiger, Bongo, Yellow-Backed Duiker
and Giant Panda) to be turned into iPSCs and then differentiated into sperm (Fig. 1). Our central hypothesis
is that overexpression of stem cell genes (POU5F1, SOX2, NANOG, LIN28, C-MYC and KLF4) in
endangered species fibroblast cells will result in reprogramming of cells into induced pluripotent stem
cells. Induced pluripotent stem cells can then be differentiated into haploid spermatids. The following
aims will test this hypothesis:
Specific Aim 1. Determine if overexpression of POU5F1, SOX2, NANOG, LIN28,
C-MYC and KLF4 can cause formation of endangered species induced
pluripotent stem cells. The goal of this aim is to produce iPSCs capable of expressing pluripotency
markers and differentiating into all three germ layers (ecto-, endo- and mesoderm), confirming their stem cell
identity.
Specific Aim 2. Examine the ability of induced pluripotent stem cells to
differentiate into sperm cells and assess stage of development. Differentiated iPSCs
are anticipated to express the definitive germ cell marker DDX4 and the pluripotency marker POU5F1,
demonstrating a germ cell state. These cells will be analyzed to determine stage of development and their
ability to undergo meiosis, a defining event where diploid cells are reduced to a haploid state.
II. Background and Significance
The IUCN 2008 Red List of Threatened Species has
recently found that greater than 25% of all mammals
are considered “threatened” with many of these
species demonstrating declining numbers. Zoological
parks across the world are attempting to combat
mass despeciation using diverse strategies such as
breeding programs. The success of these breeding
programs is heavily dependent on the ability to
maintain genetic diversity as inbreeding has been
shown to cause fertility problems, decreased litter
sizes, and health issues that ultimately lead to
unsustainable populations [7-11]. The development
of assisted reproductive technologies has provided
important tools to help preserve genetics. For Figure 2. Sus scrofa iPSCs Resemble ESCs and Are Easily
example, the ability to cryopreserve sperm samples Transduced. iPSCs were produced from mesenchymal cells by
for extended periods allows for the conservation of lentiviral transduction of POU5F1, SOX2, NANOG, KL4, LIN28 and
C-MYC genes; resulted in cells demonstrating colonial growth
vital genetics and the coupling of this technique with patterns, high refractivity and nuclear to cytoplasmic ration
artificial insemination allows sperm samples to be resembling ESCs (A 10x, B 20x). iPSCs transduced using a
used at high efficiency. However, these samples are lentiviral GFP expression vector system demonstrated strong GFP
expression (C and D).
finite with no potential for replenishing leading to the
inevitable loss of genetic diversity.
In 2006, it was first demonstrated that adult cells could be returned to an embryonic or pluripotent- a stage
where a cell has the potential to give rise to all cells of the body- state in the mouse by the introduction of
reprogramming factors POU5F1, SOX2, C-MYC and KLF4 [12]. These mouse induced pluripotent stem cells
(miPSCs) demonstrated pluripotent gene and protein expression and the ability to form teratomas- a mass of
cells showing differentiation into all 3 germ layers. These cells were injected into blastocysts and proved that
they could function correctly and contribute to multiple cell lineages in the body and ultimately proved capable
of forming germ cells [13]. Employing the reprogramming factors POU5F1, SOX2, C-MYC, KLF4, and
combinations of additional factors NANOG and LIN28, cell reprogramming has been demonstrated in rodent,
non-human primate and human cells [2-6, 14]. In addition, reprogramming
genes have shown a high degree of conservation between species where
homologous genes have been capable of reprogramming cells of other
species [15], suggesting universal importance. We have recently found that
these factors are capable of reprogramming domestic pig (Sus scrofa)
mesenchymal cells into iPSCs (see preliminary data). As a pluripotent and
immortal cell type, iPSCs represent an endless source of cells capable of
developing into any cell type of the body including germ cells.
The ability to differentiate embryonic stem cells (ESCs)- cells believed to be
almost identical to iPSCs- into germ cells has proven to be challenging with
low percentages (69%) of germ cells (see preliminary data [1]). ESC derived germ
cells have shown faithful development with re-setting of epigenetic
programming, expression of genetic markers and advanced stages of
differentiation including meiosis and morphological features such as flagella [1,
16, 18, 20]. In addition, germ cell differentiation cultures have demonstrated
the ability to give rise to supporting cells such as theca and sertoli cells,
which are essential for recreation of the germ cell niche [17, 21]. These cells
have been found to produce androgens important for normal germ cell
development. ESC derived germ cells have even proven capable of
producing male germ cells that give rise to live offspring [22], an important proof
of principle.
The development of iPSCs and a culture system capable of differentiating
stem cells into germ cells provides a unique and useful opportunity for
endangered species conservation. One of the significant challenges of
captive breeding programs is the small number of individuals per species
capable of breeding. Combining these techniques, we can preserve
important genetics and create a genetically viable population by generating
Figure 3. Sus scrofa iPSCs Express
pluripotent and immortal cells that can be infinitely expanded, cryogenically ESC Markers. iPSCs express the ESC
preserved (banked) and used to form germ cells from living or deceased markers POU5F1, SOX2, SSEA4 and
TRA1-81 after continual expansion for
animals. iPSC derived germ cells allow for the expansion of the current 20 passages demonstrating a
captive gene pools as cells can be collected from wild specimens and turned pluripotent state. 40x.
into germ cells for the use in captive breeding programs. This can be done
without removing animals from their natural habitat and inflicting further damage to delicate ecosystems. In
addition, derived germ cells can be used to study fertility problems and potently cure afflicted individuals or
used to add specific genes which would confer resistance to many diseases. The development of endangered
species iPSCs and germ cells not only provides a useful tool to increase captive gene pools, but provides a
significant advantage for the overall breeding program.
III. Preliminary Data
Reprogramming of Sus scrofa Mesenchymal Cells into Induced Pluripotent Stem Cells
Sus scrofa iPSCs were produced from mesenchymal cells by lentiviral transduction of POU5F1, SOX2,
NANOG, KL4, LIN28 and C-MYC genes; resulting in cells that closely resemble ESCs. These cells grow in
colonies with defined borders, are highly refractive and posses a high nuclear to cytoplasm ratio (Fig. 2A and
B) similar to ESCs. Cells have been cultured for >20 passages, while maintaining stem cell morphology and
expression of POU5F1, SOX2, SSEA4, TRA1-81 (Fig. 3) stem cell markers. These cells are still being
maintained to show their ability to be kept in a state of continual self renewal. To capitalize on the utility of
iPSCs, it is of importance to be able to genetically manipulate these cells. Using a lentiviral green fluorescent
protein (GFP) expression vector, we produced iPSCs that strongly express the GFP protein and show no
indication of silencing (Fig. 2C and D). In addition these cells can be maintained in feeder free conditions (data
not shown), eliminating cross species contamination and providing a robust culture system for expansion.
Differentiation of Germ Cells from Human Embryonic Stem Cells
Starting populations of BGO1 (XY) hESCs were grown on
mitotically inactivated mouse embryonic fibroblasts feeders in
20% KSR medium. BGO1 cells were differentiated into germ
cells by every other day media changes and discontinuing
passaging for 10 days to encouraging germ cell signaling. In
most cases, hESCs demonstrated 4,6-diamidino-2-phenylindole
(DAPI) (Fig. 4A) nuclear staining with co-localization of the
pluripotency marker POU5F1(Fig. 4C) and absence of the germ
cell marker DDX4 (Fig. 4E, 4G, merge). After 10 days of
differentiation, the pluripotency marker POU5F1 (Fig. 3D) and
germ cell marker DDX4 (Fig. 4F) showed nuclear co-localization
with DAPI (Fig.4B, 4H, merge). DDX4+ POU5F1+ cells were
found to be in large clusters, suggesting germ cell signaling
events.
Meiosis is a key step in germ cell development and an indicator
of germ cell formation in vitro. To further confirm germ cell
identity and meiotic activity, hESCs were differentiated for 10, 16,
and 30 days and immunostained for MLH1, a protein essential for
meiotic chiasmata formation [25, 26], and SYCP3, a protein
involved in the formation of synaptonemal complexes in meiosis
[27-29]. Immunostaining showed that >90% of day 16 cells were
positive for MLH1 (Fig. 5E and 4F) and SYCP3 (Fig. 5G and 5H)
proteins, whereas no expression of either marker was found in
hESCs (Fig. 5A–D), day 10 (data not shown), or day 30 cells.
Staining was localized to the nucleus, which correlates with their Figure 4. Expression of DDX4/POU5F1 Protein
known role in chromosome segregation during meiosis [25, 28-30]. is Up Regulated Under Differentiation
Conditions. hESCs were cultured on feeders in
20% KSR media with or without bFGF (4 ng/ml)
IV. Project Design for 3, 10 (shown) or 30 days. DAPI (A) nuclear
staining of hESCs (control, top row) exhibited
Hypothesis: We hypothesize that overexpression of stem cell colocalization with the pluripotency marker
genes (POU5F1, SOX2, NANOG, LIN28, C-MYC and KLF4) in POU5F1 (C) and absence of the germ cell marker
endangered species fibroblast cells will result in DDX4 (E, G merge). After 10 days of
differentiation, the pluripotency marker POU5F1
reprogramming of cells into induced pluripotent stem cells. (D) and germ cell marker DDX4 (F) displayed
Induced pluripotent stem cells can then be differentiated into colocalization with DAPI (B, H merge), with similar
results seen at day 30. 40x, insets 100x.
haploid germ cells.
Specific Aim 1. Determine if overexpression of POU5F1, SOX2, NANOG, LIN28, C-MYC and KLF4 can
cause formation of endangered species induced pluripotent stem cells.
Experimental Design
Six candidate species (Feline- Clouded Leopard, African Lion, Sumatran Tiger; Bovine-Bongo and Yellow-
Backed Duiker; Giant Panda) were chosen for this project based on the abundance of information pertaining to
feline and bovine reproductive anatomy and physiology (Fig. 1). The Giant Panda was also selected as it is
one of the iconic species that is critically threatened. Fibroblast cells will be collected by skin biopsy from a
single male specimen during standard examination for each species (total of 6 samples). Biopsies will be
placed into 50 ml conical tubes containing fibroblast medium composed of Dulbecco’s Modified Eagle Medium
(DMEM)/F12 high glucose base medium supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine,
50 units/ml penicillin/50 ug/ml streptomycin and β-mercaptoethanol and transported to UGA on ice. Biopsies
will then be trypsinized and disassociated into a single cell state. Fibroblast cells will be expanded on tissue
culture treated flasks incubated at 37° C in 5% CO2 with every other day media changes.
Fibroblast cells for each species will be transduced using lentiviral particles expressing elongation factor-1 (EF-
1) alpha promoter-driven constructs for POU5F1, SOX2, NANOG, LIN28, C-MYC and KLF4 at a MOI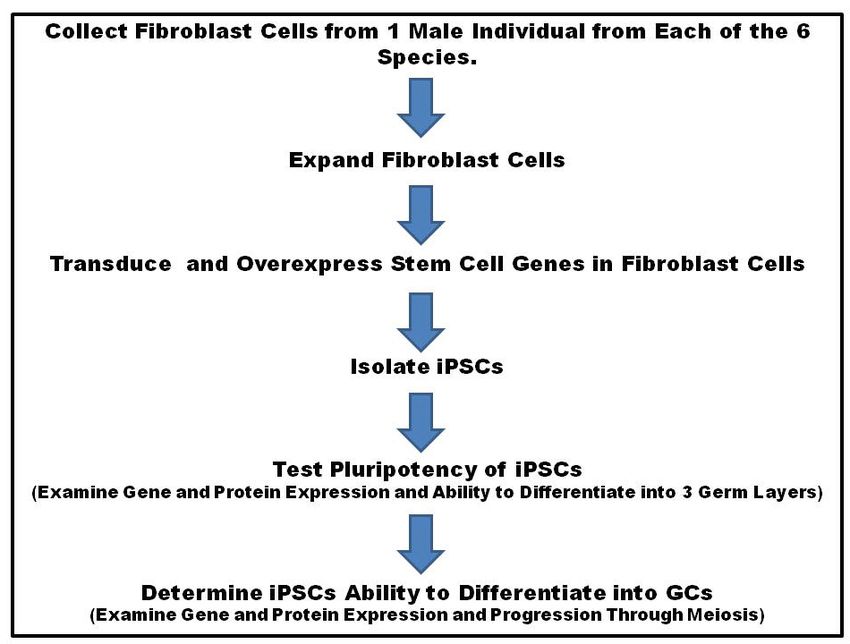
(multiplicity of infection) of 20 per factor and GeneJammer- a reagent proven to increase transduction
efficiency [31, 32]. Transduced cells will then be placed on feeder for expansion. iPSC colonies will be
collected based on stem cell morphology from day 15 to 20 and expanded under feeder free condition. All
derived lines will be tested for pluripotency by examining protein expression of SSEA4, SSEA1, TRA-1-81,
TRA-1-60 by immunocytochemistry and flow cytometry. Additionally, these cells will be tested for their ability to
differentiate into all 3 germ-layers (ecto-, endo- and mesoderm) by embryoid body formation and testing for
gene expression indicative of all 3 lineages.
Specific Aim 2. Examine the ability of induced pluripotent stem cells to differentiate into germ cells and
assess stage of development.
Experimental Design
Endangered species iPSCs lines derived in
Aim 1 will be differentiated into germ cells in
20% KSR media on polyornithine and
laminin coated plates for 0 (control), 10 and
16 days. Cells will not be passaged and
media will be changed every other day to
stimulate endogenous germ cell signaling.
Figure 5. Expression of Meiotic Markers in Differentiated Cultures. hESCs
Day 0, 10 and 16 cells will be tested for germ were differentiated on feeders for 10, 16 and 30 days in 20% KSR
cell character by determining the protein medium with bFGF (4 ng/ml). Undifferentiated hESCs (control, top row) did not
express the meiotic markers MLH1 (A, merge (B) with 4,6-diamidino-2-
expression of the germ cell markers DDX4 phenylindole [DAPI]) or SYCP3 (C, merge (D) with DAPI). After 16 days of
and POU5F1 by immunocytochemistry and differentiation, the meiotic markers MLH1 (E, merge (F) with DAPI) and SYCP3
flow cytometry. Further characterization of (G, merge (H) with DAPI) displayed co-localization with DAPI. This was not
observed at day 10 or day 30. 40x.
developmental stage will be determined by
examining the expression of additional germ cell markers using reverse transcription polymerase chain
reaction (RT-PCR). Ultimately, the meiotic ability of iPSC derived germ cells will be tested be examining the
cells for SYCP3 and MLH1 expression through immunocytochemistry and flow cytometry. As germ cells
complete normal meiosis, they are expected to move from a diploid (2n) state to a haploid (1n) state, we will
determine if these cells demonstrate an appropriate reduction in DNA content by DAPI DNA staining and
analysis by flow cytometry.
Anticipated Results
We expect that the overexpression of the pluripotent genes POU5F1, SOX2, NANOG, LIN28, C-MYC and
KLF4 will cause reprogramming of fibroblast cells of all six species to a pluripotent state. We believe that
iPSCs will demonstrate morphology (i.e. high nuclear to cytoplasm ratio, continual proliferation),
immunoreactivity (i.e. SSEA4 and TRA-1-81 expression) and ability to differentiate into cells of all 3 germ
layers confirming reprogramming of fibroblast into a stem cell state. After germ cell differentiation, we
anticipate that a sub-set of cells will be DDX4+ POU5F1+ at day 10 and SYCP3+ MLH1+ at day 16. We expect
that a portion of SCYP3 and MLH1 positive cells may be capable of completing meiosis and exist in a haploid
state.
V. Conclusion
The future of many endangered species has reached a critical stage where help from humans is essential for
continued survival. Captive breeding programs and the assisted reproductive technologies utilized in these
efforts have become important conservation tools. However, these programs are still challenged by small and
unsustainable genetic populations, fertility issues faced by many species and loss of valuable genetics
because of death or other means. In this proposal, we describe a novel conservation strategy to develop
endangered species sperm cells from fibroblast derived iPSCs. This technology is minimally invasive requiring
only a small skin sample, however the implications of this potential breakthrough is considerable. These cells
will allow conservation of important genetics through cell banking, essentially increasing captive gene pools
from hundreds of individuals to thousands, providing a way to study infertility and may even be used to provide
genetic protection from disease. Endangered species iPSC derived germ cells demonstrate significant promise
and represent a critical advance in the field of conservation.Bibliography
1. West, F.D., et al., Enrichment and differentiation of human germ-like cells mediated by feeder cells and
basic fibroblast growth factor signaling. Stem Cells, 2008. 26(11): p. 2768-76.
2. Liao, J., et al., Generation of induced pluripotent stem cell lines from adult rat cells. Cell Stem Cell,
2009. 4(1): p. 11-5.
3. Wernig, M., et al., In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature,
2007. 448(7151): p. 318-24.
4. Liu, H., et al., Generation of induced pluripotent stem cells from adult rhesus monkey fibroblasts. Cell
Stem Cell, 2008. 3(6): p. 587-90.
5. Takahashi, K., et al., Induction of pluripotent stem cells from adult human fibroblasts by defined factors.
Cell, 2007. 131(5): p. 861-72.
6. Park, I.H., et al., Reprogramming of human somatic cells to pluripotency with defined factors. Nature,
2008. 451(7175): p. 141-6.
7. Roldan, E.R., et al., Inbreeding and reproduction in endangered ungulates: preservation of genetic
variation through the Organization of Genetic Resource Banks. Reprod Domest Anim, 2006. 41 Suppl
2: p. 82-92.
8. Miller, C.R. and L.P. Waits, The history of effective population size and genetic diversity in the
Yellowstone grizzly (Ursus arctos): implications for conservation. Proc Natl Acad Sci U S A, 2003.
100(7): p. 4334-9.
9. Liberg, O., et al., Severe inbreeding depression in a wild wolf (Canis lupus) population. Biol Lett, 2005.
1(1): p. 17-20.
10. Fernandez, J., M.A. Toro, and A. Caballero, Management of subdivided populations in conservation
programs: development of a novel dynamic system. Genetics, 2008. 179(1): p. 683-92.
11. Russello, M.A. and G. Amato, Ex situ population management in the absence of pedigree information.
Mol Ecol, 2004. 13(9): p. 2829-40.
12. Takahashi, K. and S. Yamanaka, Induction of pluripotent stem cells from mouse embryonic and adult
fibroblast cultures by defined factors. Cell, 2006. 126(4): p. 663-76.
13. Okita, K., T. Ichisaka, and S. Yamanaka, Generation of germline-competent induced pluripotent stem
cells. Nature, 2007. 448(7151): p. 313-7.
14. Yu, J., et al., Induced pluripotent stem cell lines derived from human somatic cells. Science, 2007.
318(5858): p. 1917-20.
15. Li, W., et al., Generation of rat and human induced pluripotent stem cells by combining genetic
reprogramming and chemical inhibitors. Cell Stem Cell, 2009. 4(1): p. 16-9.
16. Geijsen, N., et al., Derivation of embryonic germ cells and male gametes from embryonic stem cells.
Nature, 2004. 427(6970): p. 148-54.
17. Hubner, K., et al., Derivation of Oocytes from Mouse Embryonic Stem Cells. Science, 2003. 300(5623):
p. 1251-1256.
18. Toyooka, Y., et al., Embryonic stem cells can form germ cells in vitro. Proc Natl Acad Sci U S A, 2003.
100(20): p. 11457-62.
19. Clark, A.T., et al., Spontaneous differentiation of germ cells from human embryonic stem cells in vitro.
Hum Mol Genet, 2004. 13(7): p. 727-39.
20. Tilgner, K., et al., Isolation of Primordial Germ Cells from Differentiating Human Embryonic Stem Cells.
Stem Cells, 2008.
21. Bucay, N., et al., A Novel Approach for the Derivation of Putative Primordial Germ Cells and Sertoli
Cells from Human Embryonic Stem Cells. Stem Cells, 2008.
22. Nayernia, K., et al., In vitro-differentiated embryonic stem cells give rise to male gametes that can
generate offspring mice. Dev Cell, 2006. 11(1): p. 125-32.
23. Roelke-Parker, M.E., et al., A canine distemper virus epidemic in Serengeti lions (Panthera leo).
Nature, 1996. 379(6564): p. 441-5.
24. Leisewitz, A.L., et al., Canine distemper infections, with special reference to South Africa, with a review
of the literature. J S Afr Vet Assoc, 2001. 72(3): p. 127-36.
25. Edelmann, W., et al., Meiotic pachytene arrest in MLH1-deficient mice. Cell, 1996. 85(7): p. 1125-34.
26. Barlow, A.L. and M.A. Hulten, Crossing over analysis at pachytene in man. Eur J Hum Genet, 1998.
6(4): p. 350-8.27. Di Carlo, A.D., G. Travia, and M. De Felici, The meiotic specific synaptonemal complex protein SCP3 is
expressed by female and male primordial germ cells of the mouse embryo. Int J Dev Biol, 2000. 44(2):
p. 241-4.
28. Yuan, L., et al., The murine SCP3 gene is required for synaptonemal complex assembly, chromosome
synapsis, and male fertility. Mol Cell, 2000. 5(1): p. 73-83.
29. Yuan, L., et al., Female germ cell aneuploidy and embryo death in mice lacking the meiosis-specific
protein SCP3. Science, 2002. 296(5570): p. 1115-8.
30. Codina-Pascual, M., et al., Synapsis and meiotic recombination analyses: MLH1 focus in the XY pair as
an indicator. Hum. Reprod., 2005. 20(8): p. 2133-2139.
31. Bosch, P., et al., Efficient adenoviral-mediated gene delivery into porcine mesenchymal stem cells. Mol
Reprod Dev, 2006. 73(11): p. 1393-403.
32. Fouletier-Dilling, C.M., et al., Novel compound enables high-level adenovirus transduction in the
absence of an adenovirus-specific receptor. Hum Gene Ther, 2005. 16(11): p. 1287-97.You can also read

















































