Buckhorn plantain (Plantago lanceolata) resistant to 2,4-D in Pennsylvania and alternative control options
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Weed Technology Buckhorn plantain (Plantago lanceolata)
www.cambridge.org/wet
resistant to 2,4-D in Pennsylvania and
alternative control options
Travis R. Russell1 , Tim T. Lulis2, Brian A. Aynardi3, Kaiyuan T. Tang1 and
Research Article
John E. Kaminski4
Cite this article: Russell TR, Lulis TT,
Aynardi BA, Tang KT, Kaminski JE (2021) 1
Graduate Research Assistant, Department of Plant Science, The Pennsylvania State University, University Park,
Buckhorn plantain (Plantago lanceolata)
resistant to 2,4-D in Pennsylvania and PA, USA; 2Research Technician, Department of Plant Science, The Pennsylvania State University, University
alternative control options. Weed Technol. 35: Park, PA, USA; 3Northeast Research Scientist, PBI-Gordon Corp., Shawnee, KS, USA and 4Professor, Department
297–303. doi: 10.1017/wet.2020.116 of Plant Science, The Pennsylvania State University, University Park, PA, USA
Received: 30 April 2020
Abstract
Revised: 29 September 2020
Accepted: 19 October 2020 Buckhorn plantain populations purportedly resistant to 2,4-D were identified in Pennsylvania
First published online: 4 November 2020 following long-term, continual applications of the active ingredient to turfgrass. The research
Associate Editor:
objectives of this study were to 1) confirm 2,4-D resistance with dose-response experiments, 2)
Patrick E. McCullough, University of Georgia confirm field resistance of buckhorn plantain to 2,4-D in Pennsylvania, and 3) evaluate
alternative herbicides for 2,4-D-resistant buckhorn plantain. Greenhouse dose-response experi-
Nomenclature: ments evaluated the sensitivity of buckhorn plantain biotypes that were resistant or susceptible
2, 4-D; halauxifen-methyl; buckhorn plantain,
to 2,4-D, and to halauxifen-methyl, two synthetic auxin herbicides from different chemical
Plantago lanceolata L. PLALA
families. The resistant biotype was ≥11.3 times less sensitive to 2,4-D than the susceptible
Keywords: biotype and required a 2,4-D dosage ≥4.2 times greater than the standard application rate
halauxifen-methyl; herbicide resistance; to reach 50% necrosis. No cross-resistance was observed to halauxifen-methyl because both
2, 4-D-resistance; synthetic auxin resistance; resistant and susceptible populations demonstrated similar herbicide sensitivity. Field experi-
turfgrass weed control
ments confirmed previous reports of ineffectiveness (≤30% reduction) with 2,4-D and other
Author for correspondence: phenoxycarboxylic herbicides in potentially resistant buckhorn plantain biotypes. Treatments
Travis R. Russell, Graduate Research Assistant, containing halauxifen-methyl resulted in a ≥70% reduction in resistant biotypes. This is the first
Department of Plant Science, The Pennsylvania known report of synthetic auxin herbicide resistance in any weed species in Pennsylvania and
State University, 21 Tyson Building,
University Park, PA 16802
highlights emerging herbicide resistance challenges in turfgrass systems.
Email: trr33@psu.edu
Introduction
2,4-Dichlorophenoxyacetic acid (2,4-D), a synthetic auxin herbicide discovered and commer-
cially introduced in the 1940s, has been widely used for broadleaf weed control in crop and
non-crop lands since becoming available on the herbicide market (Peterson 1967, Peterson
et al. 2016). Within the home and garden sector that encompasses residential landscapes,
2,4-D is the most commonly used pesticide active ingredient (Grube et al. 2011). Across all
agricultural sectors, a total of 11,611,964 kg of 2,4-D was applied in 2011 and the rising adoption
of 2,4-D-resistant genetically engineered crops is expected to significantly increase annual
2,4-D use (USDA 2014).
Turfgrass covers more than 16 million hectares in the United States across residential lawns,
parks and school playgrounds, golf courses and sports fields, commercial and institutional land-
scapes, and cemeteries (Milesi et al. 2005). In these turf areas, weedy plants such as buckhorn
plantain, can emerge due to improper maintenance, environmental changes, and herbicide
resistance (McCarty et al. 2008). Practices such as proper mowing, fertilizing, irrigating, and
hand pulling combined with prudent herbicide applications are important components of
an integrated weed management strategy. If herbicide use is necessary, 2,4-D applied post-
emergence is a commonly used and efficacious treatment for broadleaf weed control in grassy
areas (Elmore 1996).
Plants can be tolerant or sensitive to 2,4-D, and such selectivity is influenced by translocation
of 2,4-D in plants (Pillmoor and Gaunt 1981). In a general sense, broadleaf dicots are sensitive to
2,4-D, whereas monocots such as grasses, are tolerant (Peterson et al. 2016). Sensitive species
© The Author(s), 2020. Published by Cambridge metabolize 2,4-D through direct conjugation resulting in phytotoxic metabolites. By contrast,
University Press on behalf of the Weed Science tolerant species metabolize 2,4-D through ring hydroxylation resulting in nonphytotoxic
Society of America. metabolites (Peterson et al. 2016). Additionally, more recent research suggests that synthetic
auxin herbicides applied to sensitive plants triggers an up-regulation of an abscisic acid
(ABA) biosynthesis gene in the plant that increases ABA synthesis and a down-regulation of
photosynthetic machinery to incite plant death (McCauley et al. 2020).
Weed species have developed resistance to synthetic auxin herbicides, but the risk of
resistance development is low, with most instances occurring in situations of repeated use over
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116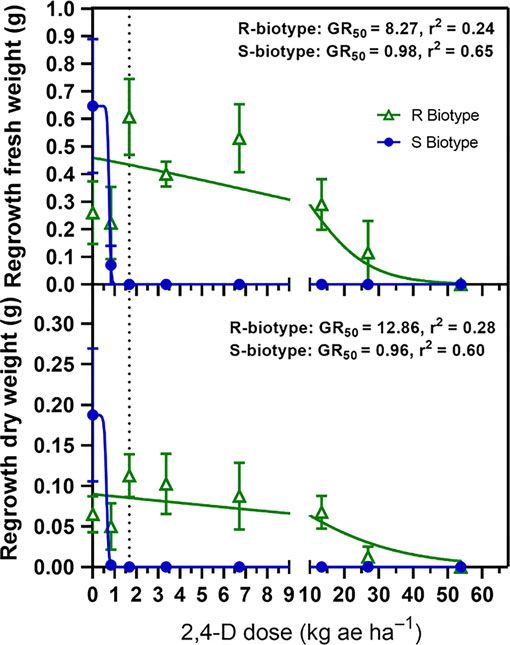
298 Russell et al.: 2,4-D-resistant Buckhorn Plantain
long periods of time (>20 yr; Beckie and Tardif 2012; Peterson et al. binding affinity at the site of action for the herbicide (Bell et al.
2016). Therefore, development of resistance to these herbicides 2014; Schmitzer et al. 2015). Further evaluation of halauxifen-
could be troublesome and would limit the options of selective her- methyl is needed in turfgrass systems to understand the role it
bicides for weed control in managed turfgrass systems (Beckie and may play in buckhorn plantain and other broadleaf weed control.
Tardif 2012; Patton et al. 2018). The resistance mechanism in Two populations of buckhorn plantain in Pennsylvania, one
plants to synthetic auxin herbicides is not completely understood, at a college campus and the other on a golf course, were suspected
but substantial progress has been made in unravelling the phe- as being resistant to 2,4-D-containing herbicide applications after
nomenon. Interaction of specific auxin-binding proteins and auxin long-term, repeated use. Thus, the objectives of this study were to
receptors have been found to be involved with auxin transport 1) confirm 2,4-D resistance in greenhouse dose-response experi-
processes and these mechanisms are thought to play a role in ments, 2) confirm field resistance of buckhorn plantain to 2,4-D
synthetic auxin herbicide resistance development (Christoffoleti in Pennsylvania, and 3) evaluate alternative commercially available
et al. 2015; Dharmasiri et al. 2005; Jugulam et al. 2011; Peterson herbicides to control buckhorn plantain in turfgrass that has
et al. 2016). become resistant to 2,4-D.
Forty weed species have been confirmed to have resistance to
synthetic auxin herbicides, with 14 of those confirming resistance
Materials and Methods
only to 2,4-D (Heap 2020). Seven of the 40 species resistant to
synthetic auxin herbicides are confirmed to have cross-resistance Site History
to multiple chemical families of herbicides in the synthetic auxin
Two sites of buckhorn plantain populations, one in Harrisburg, PA
grouping (Heap 2020). Most weed species resistant to synthetic
(40.20°N, 76.74°W) and one in Carlisle, PA (40.17°N, 77.28°W),
auxin herbicides have an annual life cycle except for spotted
were reported to not be controlled after repeated, long-term
knapweed (Centaurea stoebe L.), Canada thistle [Cirsium arvense
(>20 yr) 2,4-D applications. The soil type for the Harrisburg site
(L.) Scop.], tall buttercup (Ranunculus acris L.), and buckhorn
was a Chavies fine sandy loam (coarse-loamy, mixed, active, mesic
plantain. To our knowledge, there have been no previous reports
Ultic Hapludalfs) and the Carlsle site was a Duffield silt loam (fine-
of synthetic auxin resistance for any weed species in Pennsylvania.
loamy, mixed, active, mesic Ultic Hapludalfs). Both locations were
Buckhorn plantain is a perennial broadleaf plant that is
maintained as cool-season turfgrass landscapes mowed at 5.1 to
common in home lawns, landscapes, orchards, nursery crops
7.6 cm. At trial initiation, buckhorn plantain constituted less
and other perennial crops, and roadsides (Cavers et al. 1980;
than 10% of green plant coverage at the Harrisburg site and
Uva et al. 1997). Buckhorn plantain tolerates close mowing and
66% coverage at the Carlisle site. Greenhouse and field experiments
is commonly found on drier sites and in soils with a neutral or basic
were initiated to assess potential resistance to 2,4-D for each of
pH (Uva et al. 1997). The weed may crowd out desirable plant spe-
these buckhorn plantain populations.
cies and compete for soil nutrients, water, and light that could
impact functionality and aesthetics of the landscape. Several pre-
Greenhouse Dose-Response Experiments
vious studies have evaluated herbicide efficacy on controlling
buckhorn plantain in grassy areas (Branham 1990; McCullough Mature buckhorn plantain plants from the Harrisburg site were
et al. 2015; Patton et al. 2018; Watschke 1983). Among the collected with a hand trowel to preserve root system and trans-
synthetic auxin herbicides, active ingredients from the phen- planted into 11.4-cm-diameter and 10.2-cm-tall, round, plastic
oxycarboxylic acid chemical family including 2,4-D and others pots with potting soil growing media (Sunshine Mix #4, Sun
(dichlorprop, MCPA, and mecoprop) are generally effective in Gro Horticulture, Agawam, MA 01001) and allowed to establish
controlling buckhorn plantain, whereas the efficacy of pyridinecar- at the Pennsylvania State University (University Park, PA) green-
boxylic acids (clopyralid, fluroxypyr, triclopyr) and benzoic acids house complex. Additionally, a second buckhorn plantain collec-
(dicamba) are variable or do not effectively control the weed tion was obtained from a susceptible population in University
(Patton et al. 2019). Park, PA (40.81°N, 77.86°W). Individual potted plants were irri-
The first confirmed population of buckhorn plantain with gated and fertilized as needed to facilitate plant establishment.
resistance to 2,4-D and other phenoxycarboxylic acid herbicides Fertilization included two applications of a water-soluble fertilizer
was found in Indiana on cemetery grounds following 30 yr of (N-24 P-8 K-16; Miracle-Gro All Purpose Plant Food, Scotts
2,4-D applications to control broadleaf weeds (Patton et al. 2018). Miracle-Gro, Marysville, OH 43040) to provide 24.4 kg N ha−1.
In greenhouse dose-response experiments, the resistant buckhorn Buckhorn plantain were established in the greenhouse for 8 wk
plantain biotype was more than 6-fold less sensitive to 2,4-D com- before dose-response experiments were initiated. Greenhouse con-
pared to a susceptible biotype (Patton et al. 2018). In comparison, ditions maintained a 16/8 hr day/night cycle. Greenhouse air tem-
responses to triclopyr, a pyridinecarboxylic acid, were similar in perature was maintained above 18 C with cooling systems
both biotypes, indicating a lack of cross-resistance to other chemi- initiating when temperatures exceeded 25 C. Experiments were
cal families in the synthetic auxins. Field experiments provided conducted in October 2018 and repeated in June 2019.
more evidence that resistance was confined to the phenoxycarbox- This experiment was arranged as a randomized complete block
ylic acid chemical family because other herbicides from the pyri- design in a 2 by 8 factorial treatment arrangement within each her-
dinecarboxylic acid (clopyralid, triclopyr) and arylpicolinate bicide with four replications serving as blocks. Treatment factors
(halauxifen-methyl) chemical families were efficacious against included two buckhorn plantain biotypes (resistant or susceptible)
the resistant biotypes (Patton et al. 2018). Halauxifen-methyl is and eight doses of each herbicide. Each biotype was treated with
a new synthetic auxin herbicide that has been demonstrated to doses 0, 0.84, 1.68, 3.36, 6.72, 13.4, 26.9, and 53.8 kg ae ha−1
be an efficacious alternative to 2,4-D and dicamba for controlling of 2,4-D dimethylamine (Gordon’s Amine® 400, PBI-Gordon
horseweed [Conyza canadensis (L.) Cronq.] and redroot pigweed Corp., Shawnee, KS 66226), with 1.68 kg ae ha−1 serving as the
(Amaranthus retroflexus L.) in cropping systems (Mccauley and standard application rate. Similarly, each biotype was treated with
Young 2019; Zimmer et al. 2018). This efficacy is due to differing technical grade halauxifen-methyl (GF-2685, Corteva Agriscience,
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116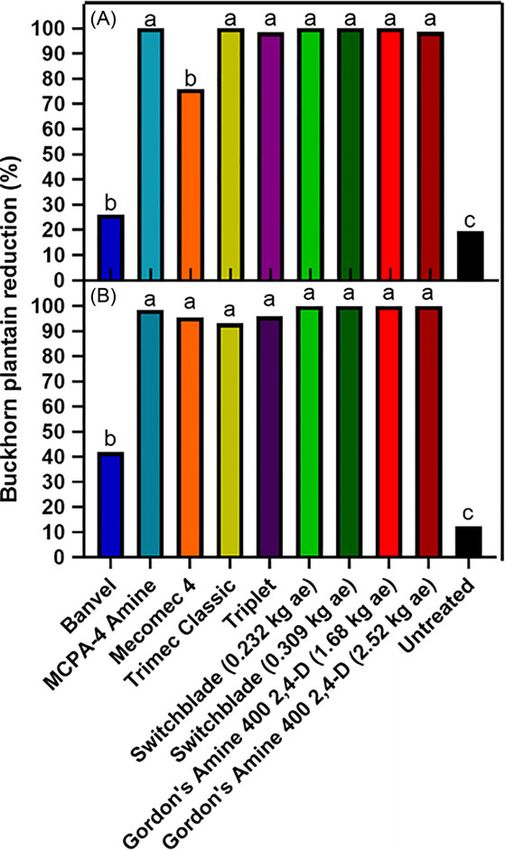
Weed Technology 299
Wilmington, DE 19805) at doses of 0, 0.003, 0.005, 0.01, 0.02, 0.04, Corp.), dicamba (Banvel®, Micro Flo Company, Memphis, TN
0.08, and 0.16 kg ae ha−1, with 0.01 kg ae ha−1 being the standard 38117), 2,4-D þ MCPP þ dicamba (Triplet®, Nufarm Americas
application rate. Treatments were applied using a CO2-pressurized Inc.; Trimec Classic®, PBI-Gordon Corp.), fluroxypyr þ
backpack sprayer equipped with a single TeeJet AI9508 EVS nozzle dicamba þ halauxifen-methyl (Switchblade®, PBI-Gordon Corp.)
(TeeJet Spraying Systems Co., Wheaton, IL 60139) calibrated to at a low and high rate, and a nontreated control (Table 1). All treat-
deliver 408 L ha−1. ments are listed by their trade name in figures and tables.
Visual weed epinasty was evaluated 3 d after treatment (DAT) Buckhorn plantain coverage of herbicide treatments was
and visual necrosis was assessed 14 and 28 DAT. Epinasty was evaluated 28 DAT. Buckhorn plantain coverage in plot areas
rated on a 0% to 100% scale, with 0% equivalent to no epinasty was visually estimated on a 0% to 100% scale, with 0% equivalent
and 100% epinasty being complete twisting, bending, and curling to no buckhorn plantain and 100% representing an entire plot area
of stems and leaf tissue. Necrosis was also rated on the same 0% to consisting of buckhorn plantain. Data were transformed to percent
100% scale, except a 100% evaluation was equivalent to completely buckhorn plantain reduction by dividing buckhorn plantain cover-
dead plant tissue. Leaf chlorophyll content index (CCI) was evalu- age at 28 DAT by initial coverage prior to treatment application.
ated 28 DAT with an MC-100 Chlorophyll Concentration Meter ANOVA was conducted using the PROC MIXED procedure
(Apogee Instruments, Logan, UT 84321) by taking two measure- in SAS. Treatment means were separated using Fisher’s protected
ments of a single, representative leaf for each plant. Plants were LSD test. A value of α = 0.05 was used to determine statistical
harvested 2 cm above the soil surface 28 DAT to measure fresh significance.
and dry plant tissue weight. Harvested plant tissue was placed in
a drying oven for 7 d at 60 C. Plants were allowed to regrow for
an additional 2 wk and harvested again at 42 DAT for fresh and Results and Discussion
dry weight measurements.
Greenhouse Dose-Response Experiments
Analysis of variance was conducted with the PROC MIXED
procedure in SAS (version 9.3; SAS Institute, Cary, NC 27513). The effect of experimental run was not significant for any
Experimental run was not a significant source of variation evaluation. Therefore, data from both experiments were pooled.
(P < 0.05), therefore data were pooled across experimental runs. ANOVA revealed a significant two-way interaction of biotype
The GR50 (the herbicide dose eliciting a 50% growth reduction by and 2,4-D dosage for all evaluations except fresh and dry weight
the plant) for each herbicide and biotype was estimated using a measurements at 28 DAT, indicating differential responses of
three-parameter log-logistic dose-response curve (Seefeldt et al. 1995): buckhorn plantain to doses of 2,4-D in susceptible (S) and resistant
(R) populations (Table 2). In contrast, the two-way interaction of
DC biotype and halauxifen-methyl dosage was only significant on one
y¼Cþ b [1] of 11 evaluations, suggesting similar responses to halauxifen-
1 þ GRx 50
methyl across S and R populations.
Epinasty differed between R and S populations in response
where y is the response variable (epinasty, necrosis, etc.), GR50 is to 2,4-D and halauxifen-methyl at 3 DAT (Table 2). Across all
the herbicide dose eliciting a 50% reduction, b is the relative slope, dosages of 2,4-D, epinasty averaged 35% in the R biotypes and
D is the upper limit, C is the lower limit, and x is the rate of the 53% in S biotypes (data not shown). For halauxifen-methyl,
herbicide being used. Log-logistic regression was analyzed using epinasty averaged 25% and 36% in R and S biotypes, respectively,
Prism 8 software (GraphPad Software, La Jolla, CA 92037). The across all dosages.
GR50 of a biotype was deemed significantly different if the 95% For necrosis evaluations at 14 and 28 DAT, a significant
confidence intervals of the GR50 estimates did not overlap. two-way interaction of biotype and 2,4-D dose was observed
Resistance factors were calculated as the ratio of resistant biotype in buckhorn plantain populations (Table 2; Figure 1). The
GR50 to susceptible biotype GR50 and were used to quantify level of two-way interaction for 2,4-D-treated plants indicates that
resistance in each population with the scale employed by Becki and S and R populations had significant response differences at
Tardif (2012). specified dosages. Conversely, only the main effects of biotype
and dosage were significant in halauxifen-methyl-treated popu-
lations (Table 2). For 2,4-D, the estimated GR50 for necrosis in
Field Herbicide Efficacy Experiments
the R population 28 DAT was 7.13 kg ae ha−1, more than 4 times
Field experiments were initiated at suspected 2,4-D-resistant sites greater than the labeled application rate of 1.68 kg ae ha−1
in Harrisburg and Carlisle and also at a susceptible site in (Figure 1). In comparison, the S population had a GR50 for
University Park (40.78°N, 77.87°W) to evaluate potential alterna- necrosis of 0.63 kg ae ha−1, a dosage lower than half of the label
tive herbicides to 2,4-D for controlling resistant populations. The rate. These results are similar to those reported by Patton et al.
University Park site averaged 30% buckhorn plantain coverage and (2018) in which a resistant buckhorn plantain biotype in Indiana
the soil type was a Hagerstown silt loam (fine, mixed, semiactive, required a 2,4-D dosage at least 6.2 times greater than the sus-
mesic Typic Hapludalf). For the Harrisburg and University Park ceptible biotype.
sites, experiments were initiated in September 2018 and repeated For halauxifen-methyl treatments, necrosis at 14 and 28
in May 2019. For the Carlisle site, two experiments were initiated in DAT was significantly affected by dose and biotype main effects
May 2019 in different, but adjacent experimental areas. (Table 2). Except when compared to the nontreated control,
Field experiments were arranged in a randomized complete necrosis at 14 DAT did not differ among doses of halauxifen-
block design with four replications serving as blocks. Herbicide treat- methyl across both populations, averaging 43% necrosis (data
ments included 2,4-D dimethylamine (Gordon’s Amine® 400) at a not shown). At 28 DAT, dosages of halauxifen-methyl at or below
low (label) and high rate, MCPA (MCPA-4 Amine®, Nufarm the standard application dose of 0.010 kg ae ha−1 resulted in
Americas Inc, Alsip, IL 60803), MCPP (Mecomec 4®, PBI-Gordon necrosis greater than the nontreated control (67% compared to
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116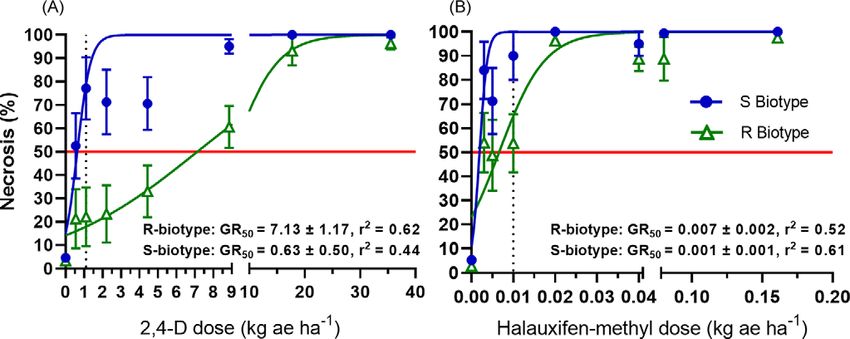
300 Russell et al.: 2,4-D-resistant Buckhorn Plantain
Table 1. Commercially available herbicides used in field efficacy experiments for control of buckhorn plantain in 2,4-D-resistant and susceptible populations.
Active ingredient Trade name Efficacya Chemical family Application rateb
———kg ae ha−1———
2,4-D dimethylamine Gordon’s Amine 400® Good Phenoxycarboxylic acid 1.68 (low)
2,4-D dimethylamine Gordon’s Amine 400® Good Phenoxycarboxylic acid 2.52 (high)
MCPA MCPA-4 Amine® Fair Phenoxycarboxylic acid 1.96
MCPP Mecomec 4® Fair Phenoxycarboxylic acid 0.30
Dicamba Banvel® Poor Benzoic acid 0.12
2,4-D dimethylamine þ MCPP þ Triplet® Excellent Phenoxycarboxylic acid þ phenoxycarboxylic 1.35 þ 0.35 þ 0.12
dicamba acid þ benzoic acid
2,4-D dimethylamine þ MCPP þ Trimec Classic® Excellent Phenoxycarboxylic acid þ phenoxycarboxylic 1.11 þ 0.29 þ 0.12
dicamba acid þ benzoic acid
Fluroxypyr þ dicamba þ Switchblade® Unknown Pyridinecarboxylic acid þ benzoic acid þ 0.139 þ 0.087 þ 0.006 (low)
halauxifen-methyl arylpicolinate
Fluroxypyr þ dicamba þ Switchblade® Unknown Pyridinecarboxylic acid þ benzoic acid þ 0.185 þ 0.116 þ 0.008 (high)
halauxifen-methyl arylpicolinate
Nontreated — — — —
a
Efficacy based on key published by Patton et al. (2019) where excellent = ≥90% control, good = 75% to 90% control, fair = 50% to 75% control, poor = F——————————————————————————
2,4-D
Biotype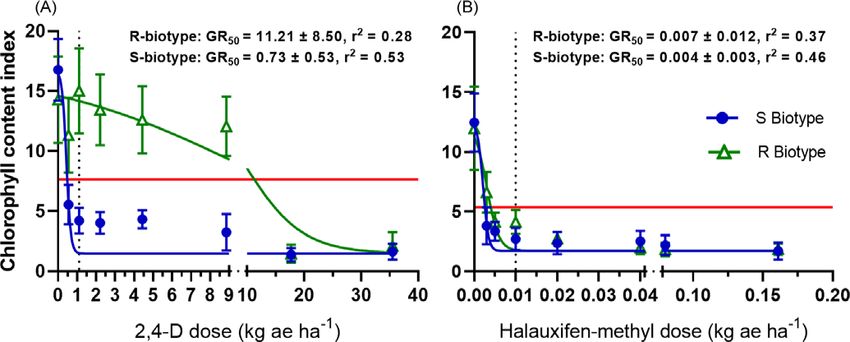
Weed Technology 301
Figure 2. Nonlinear regression of buckhorn plantain chlorophyll content index 28 d after treatment with 2,4-D (A) or halauxifen-methyl (B) in suspected-2,4-D-resistant (R)
and suspected-2,4-D-susceptible (S) biotypes in greenhouse dose-response experiments. Error bars represent the standard error of the mean. Intersection of trendline with
horizontal line at 50% chlorophyll content index is representative of the estimated GR50. Dashed vertical line indicates the labeled rate of each herbicide active ingredient.
biotypes at 28 DAT were 0.001 and 0.007 kg ae ha−1 halauxifen-
methyl, respectively, and were lower than the standard application
rate of 0.010 kg ae ha−1 (Figure 1).
CCI measurements resulted in similar trends as necrosis eval-
uations. At 28 DAT, there was a significant two-way biotype-by-
dose interaction for buckhorn plantain populations treated with
2,4-D (Table 2). Conversely, dose was the lone significant treat-
ment factor for plants treated with halauxifen-methyl, suggesting
that there were no differences among R and S biotypes in terms of
CCI when treated with this active ingredient (Table 2). Although
R2 values were lower in regression analysis and more variability
was present in CCI data than necrosis evaluations, the CCI
GR50 for the R biotype in response to 2,4-D application was
still greater (11.21 kg ae ha−1) than that of the S biotype
(0.73 kg ae ha−1) and greater than the label application 2,4-D
application rate of 1.68 kg ae ha−1 (Figure 2). The CCI GR50 values
for R and S biotypes treated with halauxifen-methyl were not dif-
ferent from each other and were less than or equivalent to the stan-
dard halauxifen-methyl application rate.
The main effect of dosage influenced fresh and dry weight
28 DAT in 2,4-D treatments, but it affected only fresh weight in
halauxifen-methyl treatments (Table 2). The loss of plant moisture
after drying could potentially explain why dosage did not affect Figure 3. Nonlinear regression of buckhorn plantain fresh (A) and dry (B) regrowth
dry weight in halauxifen-methyl treatments 28 DAT. However, weight from suspected 2,4-D-resistant (R) and suspected-2,4-D-susceptible (S)
a significant biotype-by-dose interaction was observed for 2,4-D biotypes 42 d after 2,4-D treatment following initial plant tissue harvest 28 d after
treatment in greenhouse dose-response experiments. Whole plant biomass was
treated plants regarding fresh and dry regrowth weight measure-
harvested and oven-dried at 60 C for 7 d. Error bars represent the standard error
ments taken 42 DAT after the initial harvest (Table 2). For both of the mean.
fresh and dry regrowth, the GR50 for the S biotype was less than
the label rate of 2,4-D, indicating that 2,4-D treatments are effec-
tive at controlling this biotype (Figure 3). However, R biotypes and quantify the level of 2,4-D resistance present in the resistant
needed 2,4-D dosages greater than four times the labeled rate to biotype. Although R and S biotypes had different resistance factors
exhibit a 50% reduction in fresh or dry regrowth. In both S and to halauxifen-methyl, the GR50 for each biotype was still below the
R biotypes, all rates of halauxifen-methyl resulted in complete standard application rate. These resistance factors are in the same
death of buckhorn plantain, whereas the nontreated plants grew range as those observed by Patton et al. (2018) on 2,4-D-resistant
new leaves following the initial harvest 28 DAT (data not shown). buckhorn plantain in Indiana and are consistent with studies on
This finding suggests that there is no perceived resistance to this several annual broadleaf weeds and their resistance to 2,4-D
active ingredient in the 2,4-D-resistant biotype. (Burke et al. 2009; Preston et al. 2013; Rey-Caballero et al. 2016).
Resistance factors of 11.3 for necrosis and 15.4 for CCI evalu-
ations after R and S biotypes were treated with 2,4-D indicate the
Field Herbicide Efficacy Experiments
differential responses in each population to the same herbicide.
Resistance factors greater than 10 are often considered as being Commercially available synthetic auxin herbicides were evaluated
high for synthetic auxin herbicides (Beckie and Tardif 2012) to determine buckhorn plantain control efficacy in field
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116302 Russell et al.: 2,4-D-resistant Buckhorn Plantain
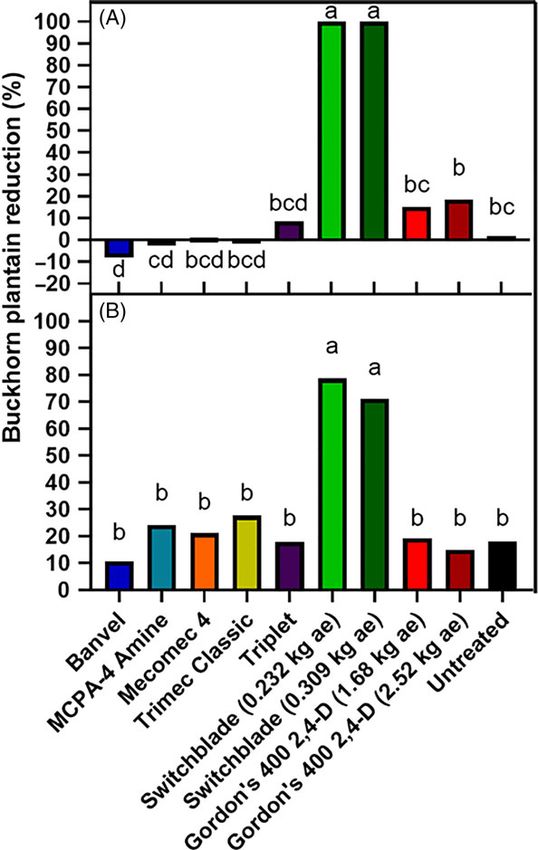
Figure 4. Buckhorn plantain cover reduction in a 2,4-D-susceptible population Figure 5. Buckhorn plantain cover reduction within 2,4-D-resistant populations in
at University Park, PA in 2018 (A) and 2019 (B) 28 d after synthetic auxin herbicide Carlisle, PA (A) and Harrisburg, PA (B) 28 d after treatment with synthetic auxin herbi-
treatment in field efficacy experiments. Within each year, means followed by the cides in field efficacy experiments. Treatment bars that do not share the same letter
same letter are not significantly different according to Fisher’s protected LSD test are significantly different according to Fisher’s protected LSD test (α = 0.05).
(α = 0.05).
treatments from the phenoxycarboxylic acid and benzoic acid
chemical families were not able to provide reduction greater than
experiments at one location with a S buckhorn plantain biotype in the nontreated, indicating resistance mechanisms to the phenox-
University Park and two locations with suspected-R biotypes in ycarboxylic acid chemical family as initially reported by Patton
Carlisle and Harrisburg. There was a significant effect of experi- et al. (2018). Alternatively, Switchblade treatments resulted in good
mental run (P = 0.0445) at the University Park site. Therefore, (70% to 90% reduction) to excellent reduction (≥90%) at both
data are presented separately by experiment. At the Harrisburg locations (Figure 5). This treatment contained active ingredients
and Carlisle sites, experimental run was not a significant effect from the pyridinecarboxylic acid (fluroxypyr) and arylpicolinate
(P = 0.5899 and 0.4300, respectively) and data were pooled across (halauxifen-methyl) chemical families in addition to a benzoic
experiments for each location. acid (dicamba) that do not have any reported cases of resistance
Initial buckhorn plantain coverage at the S biotype site averaged in buckhorn plantain. Because dicamba is not considered an
16.1% and 45.7% of plot area in 2018 and 2019, respectively effective active ingredient in controlling buckhorn plantain
(data not shown). Except for Mecomec 4 in 2018 and Banvel, all (Patton et al. 2019) and fluroxypyr has been reported to be inef-
other synthetic auxin treatments reduced buckhorn plantain fective on other 2,4-D-resistant buckhorn plantain populations
≥90% across all experiments conducted on S biotypes (Figure 4). (Patton et al. 2018), these results suggest that the effectiveness of
Banvel provided poor reduction (90%. synthetic auxins (Bell et al. 2014; Schmitzer et al. 2015) could
Across both runs of the experiment in R biotypes, initial buck- indicate why this treatment was successful in resistant popula-
horn plantain coverage averaged 10.3% at the Harrisburg location tions, as has been suggested in other studies (McCauley et al.
and 66.2% at the Carlisle location (data not shown). Gordon’s 2018). In addition to halauxifen-methyl, Patton et al. (2018) also
400 2,4-D applied at low and high rates were essentially equivalent found that other pyridinecarboxylic acid herbicides such as
to the nontreated at both resistant locations, reducing buckhorn clopyralid and triclopyr are effective in 2,4-D-resistant buckhorn
plantain cover by less than 20% (Figure 5). Additionally, all other plantain populations.
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116Weed Technology 303
In the present study, resistance in buckhorn plantain was Grube A, Donaldson D, Kiely T, Wu L (2011) Pesticides industry sales and
restricted to herbicides from the phenoxycarboxylic acid chemical usage, 2006 and 2007 Market Estimates. Washington D.C: U.S. Environmental
family of synthetic auxin herbicides and is in agreement with Protection Agency. 41 p
previous research in Indiana (Patton et al. 2018). Field evaluations Heap I (2020) The International Survey of Herbicide Resistant Weeds. http://
www.weedscience.org. Accessed: October 1, 2020
in suspected-R buckhorn plantain populations further indicated
Harrington KC, Ward AJ, Wells D. (2001) Herbicide resistance in black night-
resistance to phenoxycarboxylic acids in the synthetic auxin
shade and Onehunga weed. New Zeal Plant Prot 54:152–156
herbicides. Greenhouse and field evaluations indicate that treat- Jugulam MJ, Hall JC, Johnson WG, Kelley KB, Riechers DE (2011) Evolution
ments including halauxifen-methyl could be used to effectively of resistance to auxinic herbicides: Historical perspectives, mechanisms of
control these resistant populations. These results confirm resistance, and implications for broadleaf weed management in agronomic
resistance to 2,4-D in buckhorn plantain populations in turfgrass crops. Weed Sci 59:445–457
systems and further shed light on alternative control options in McCarty LB, Everst JW, Hall DW, Murphy TR, Yelverton F (2008) Color Atlas
R populations. Although cross-resistance to multiple chemical of Turfgrass Weeds: A Guide to Weed Identification and Control Strategies.
families within the synthetic auxins has been reported in other Hoboken, NJ: Wiley. 424 p
weed species, a majority of reports cite single chemical family McCauley CL, Johnson WG, Young BG (2018) Efficacy of halauxifen-methyl on
glyphosate-resistant horseweed (Erigeron canadensis). Weed Sci 66:758–763
resistance in synthetic auxins (Heap 2020). Specifically in turf sys-
McCauley CL, McAdam SA, Bhide K, Thimmapuram J, Banks JA, Young BG
tems, only lawn burweed has been reported to have synthetic auxin
(2020) Transcriptomics in Erigeron canadensis reveals rapid photosynthetic
resistance to the pyridinecarboxylic acid herbicides (Harrington and hormonal responses to auxin herbicide application. J Exp Bot 71:
et al. 2001). Because it is not always practical or in the research 3701–3709
objectives of a study to evaluate resistant weed species against McCauley CL, Young BG (2019) Differential response of horseweed (Conyza
all classes of an herbicide group, the disparity in cross-resistance canadensis) to halauxifen-methyl, 2,4-D, and dicamba. Weed Technol 33:
and single chemical family resistance could be artificially inflated. 673–679
There are several reports of weed species having multiple resistance McCullough PE, Johnston CR, Reed TV, Yu J (2015) Indaziflam enhances buck-
to a synthetic auxin herbicide and other herbicide modes of action horn plantain (Plantago lanceolata) control from postemergence herbicides.
(Heap 2020), but that was not evaluated in the present study. Weed Technol 29:147–153
Regardless, this is the first report of synthetic auxin resistance of Milesi C, Running SW, Elvidge CD, Dietz JB, Tuttle BT, Nemani RR (2005)
Mapping and modeling the biogeochemical cycling of turf grasses in the
any weed species in Pennsylvania. Despite substantial research
United States. Environ Manage 36:426–438
in unravelling the molecular mechanism for synthetic auxin resis- Patton AJ, Elmore M, Hoyle J, Kao-Kniffin J, Branham B, Voigt T, Christians N,
tance, additional research is needed to fully understand the factors Thoms A, Munshaw G, Hathaway A, Nikolai T, Horgan B, Miller L, Xiong X,
conferring 2,4-D resistance in buckhorn plantain. Further research Kreuser W, Gaussoin R, Gardner D, Raudenbush Z, Li D, Landschoot P,
should also focus on cultural control options and additional com- Soldat D, Koch P (2019) Turfgrass Weed Control for Professionals.
mercially available herbicide formulations for buckhorn plantain, Purdue University Extension Publication. West Lafayette, IN: Purdue
especially in resistant populations, as part of an integrated weed University. 128 p
management strategy. Patton AJ, Weisenberger DV, Schortgen GP (2018) 2,4-D-resistant buckhorn
plantain (Plantago lanceolata) in managed turf. Weed Technol 32:182–189
Acknowledgments. We thank PBI-Gordon Corp. for providing financial Peterson G (1967) The discovery and development of 2,4-D. Agric Hist 41:
funding and the facilities that allowed us to conduct the research. This work 243–254
was supported by the USDA–National Institute of Food and Agriculture, Peterson MA, McMaster SA, Riechers DE, Skelton J, Stahlman PW (2016) 2,4-D
Hatch accession 1006804 and project PEN04592. No conflicts of interest have past, present, and future: A review. Weed Technol 30:303–345
been declared. Pillmoor JB, Gaunt JK (1981) Behaviour and mode of action of the phenoxy-
acetic acids in plants. Pages 147–218 in Progress in Pesticide Biochemistry.
Chichester, UK: Wiley
References
Preston C, Dolman FC, Boutsalis P (2013) Multiple resistance to acetohydrox-
Beckie HJ, Tardif FJ (2012) Herbicide cross resistance in weeds. Crop Prot yacid synthase–inhibiting and auxinic herbicides in a population of oriental
35:15–28 mustard (Sisymbrium orientale). Weed Sci 61:185–192
Bell JL, Schmitzer PR, Robinson AE (2014) Arylex™ mode and site of action Rey-Caballero J, Menéndez J, Giné-Bordonaba J, Salas M, Alcántara R, Torra J
characterization. Proceedings of the 2014 meeting of the Weed Science (2016) Unravelling the resistance mechanisms to 2,4-D (2,4-dichlorophe-
Society of America and Canadian Weed Science Society/Société Canadienne noxyacetic acid) in corn poppy (Papaver rhoeas). Pestic Biochem Physiol
de Malherbologie. Vancouver, British Columbia, February 3–6, 2014 133:67–72
Branham BE (1990) 1989 weed control update: III. Broadleaf weed control. Schmitzer PR, Balko TW, Daeuble JF, Epp JB, Satchivi NM, Siddall TL, Weimer
Pages 22, 34–37 in Proceedings of the 60th Annual Michigan Turfgrass MR, Yerkes CN (2015) Discovery and SAR of halauxifen methyl: A novel
Conference. East Lansing, MI, January 15–17, 1990 auxin herbicide. ACS Symp Ser 1204:247–260
Burke IC, Yenish JP, Pittmann D, Gallagher RS (2009) Resistance of a Prickly Seefeldt SS, Jensen JE, Fuerst EP (1995) Log-logistic analysis of herbicide dose-
Lettuce (Lactuca serriola) Biotype to 2,4-D. Weed Technol 23:586–591 response relationships. Weed Technol 9:218–227
Cavers PB, Bassett IJ, Crompton CW (1980) The biology of Canadian Weeds: [USDA] U.S. Department of Agriculture (2014) Dow AgroScience petitions
47. Plantago lanceolata L. Canadian J Plant Sci 60:1269–1282 (09-233-01p, 09-349-01p and 11-234-01p) for determinations of non-
Christoffoleti PJ, Alves de Figueiredo MR, Pereira Peres LE, Nissen S, Gaines T regulated status for 2,4-D-resistant corn and soybean varieties. Riverdale,
(2015) Auxinic herbicides, mechanisms of action, and weed resistance: MD: Biotechnology Regulatory Services. 223 p
A look into recent. Sci Agric 72:356–362 Uva RH, Neal JC, DiTomaso JM (1997) Weeds of the Northeast. Ithaca, NY:
Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1 is an Cornell University. 397 p
auxin receptor. Nature 435:441–445 Watschke TL (1983) Broadleaf weed control in low maintenance turf. Proc
Elmore C (1996) Use of 2,4-D in orchards, vineyards, and soft fruit in the United Annu Meet Northeast Weed Sci Soc 37:377–378
States. Pages 148–164 in Biologic and Economic Assessment of Benefits from Zimmer M, Young BG, Johnson WG (2018) Weed control with halauxifen-
Use of Phenoxy Herbicides in the United States. USDA/National Pesticide methyl applied alone and in mixtures with 2,4-D, dicamba, and glyphosate.
Impact Assessment Program Report No. 1-PA-96 Weed Technol 32:597–602
Downloaded from https://www.cambridge.org/core. Pennsylvania State University, on 12 Oct 2021 at 17:15:06, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms. https://doi.org/10.1017/wet.2020.116You can also read

















































