A Neutralizing IL-11 Antibody Improves Renal Function and Increases Lifespan in a Mouse Model of Alport Syndrome
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
BASIC RESEARCH www.jasn.org
A Neutralizing IL-11 Antibody Improves Renal
Function and Increases Lifespan in a Mouse Model
of Alport Syndrome
Anissa A. Widjaja ,1 Shamini G. Shekeran,1 Eleonora Adami,1,2 Joyce G Wei Ting,1
Jessie Tan,3 Sivakumar Viswanathan ,1 Sze Yun Lim ,1 Puay Hoon Tan,1,4,5
Norbert H€ubner,2,6,7 Thomas Coffman,1 and Stuart A. Cook1,3,8
Due to the number of contributing authors, the affiliations are listed at the end of this article.
ABSTRACT
Background Alport syndrome is a genetic disorder characterized by a defective glomerular basement
membrane, tubulointerstitial fibrosis, inflammation, and progressive renal failure. IL-11 was recently impli-
cated in fibrotic kidney disease, but its role in Alport syndrome is unknown.
Methods We determined IL-11 expression by molecular analyses and in an Alport syndrome mouse
model. We assessed the effects of a neutralizing IL-11 antibody (3203) versus an IgG control in Col4a32/2
mice (lacking the gene encoding a type IV collagen component) on renal tubule damage, function, fibrosis,
and inflammation. Effects of 3203, the IgG control, an angiotensin-converting enzyme (ACE) inhibitor
(ramipril), or ramipril1X203 on lifespan were also studied.
Results In Col4a32/2 mice, as kidney failure advanced, renal IL-11 levels increased, and IL-11 expression
localized to tubular epithelial cells. The IL-11 receptor (IL-11RA1) is expressed in tubular epithelial cells and
podocytes and is upregulated in tubular epithelial cells of Col4a32/2 mice. Administration of 3203
reduced albuminuria, improved renal function, and preserved podocyte numbers and levels of key podo-
cyte proteins that are reduced in Col4a32/2 mice; these effects were accompanied by reduced fibrosis
and inflammation, attenuation of epithelial-to-mesenchymal transition, and increased expression of regen-
erative markers. X203 attenuated pathogenic ERK and STAT3 pathways, which were activated in
Col4a32/2 mice. The median lifespan of Col4a32/2 mice was prolonged 22% by ramipril, 44% with 3203,
and 99% with ramipril1X203.
Conclusions In an Alport syndrome mouse model, renal IL-11 is upregulated, and neutralization of IL-11
reduces epithelial-to-mesenchymal transition, fibrosis, and inflammation while improving renal function.
Anti-IL-11 combined with ACE inhibition synergistically extends lifespan. This suggests that a therapeutic
approach targeting IL-11 holds promise for progressive kidney disease in Alport syndrome.
JASN 33: –, 2022. doi: https://doi.org/10.1681/ASN.2021040577
Alport syndrome (AS) is caused by mutation in
the A3/4/5 genes that encode chains of type IV
Received April 29, 2021. Accepted January 7, 2022.
collagen.1,2 These mutations lead to abnormali-
ties in glomerular basement membrane (GBM) A.A.W. and S.G.S contributed equally to this work.
collagen composition, integrin-mediated podo- Published online ahead of print. Publication date available at
cyte dysfunction, glomerular hypertension, and www.jasn.org.
ultrafiltration.3,4 AS affects up to 60,000 people Correspondence: Dr. Anissa A. Widjaja or Dr. Stuart A. Cook,
in the United States and is associated with hear- Duke-National University of Singapore Medical School,
ing loss, ocular abnormalities, and CKD. 8 College Road 169857, Singapore. Email: anissa.widjaja@
duke-nus.edu.sg or stuart.cook@duke-nus.edu.sg
In the commonest form of disease due to
X-linked mutation of COL4A5, 90% of affected Copyright ß 2022 by the American Society of Nephrology
JASN 33: –, 2022 ISSN : 1533-3450/1046-667 1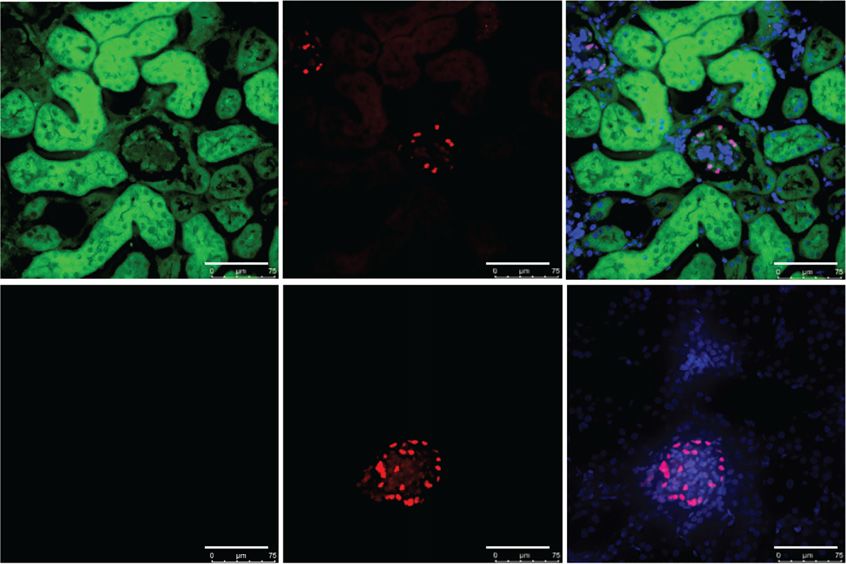
BASIC RESEARCH www.jasn.org
men develop ESKD by the age of 40.5 Early disease can
Significance Statement
manifest as hematuria, microalbuminuria, or proteinuria,
and although there are no specific therapies, affected chil- Alport syndrome (AS), a genetic disorder of the glomerular
dren are commonly treated with an angiotensin converting basement membrane, frequently leads to end stage renal failure.
In an animal model of AS—mice lacking the Col4a3 gene,
enzyme inhibitor (ACEi), on the basis in part of extrapola-
—angiotensin-converting enzyme inhibition is protective. The
tion of studies conducted in Col4a32/2 mice6 and sup- authors show that IL-11 is upregulated in the renal tubular epi-
ported by more recent data from clinical trials.7,8 thelia of Col4a32/2 mice; the IL-11 receptor (IL11RA1),
The Col4a32/2 mouse strain is widely viewed as one of expressed on podocytes and tubule cells, is upregulated in the
the best animal models of progressive AS. In seminal stud- diseased kidneys of Col4a32/2 mice. Giving 6-week-old
Col4a32/2 mice a neutralizing IL-11 antibody (X203) reduced
ies, treatment of 4-week-old Col4a32/2 mice with an ACEi
pathologic ERK and STAT3 activation and limited epithelial-to-
(ramipril), before the onset of proteinuria and tubulointer- mesenchymal transition; reduced kidney fibrosis, inflammation,
stitial fibrosis, attenuated kidney dysfunction and pro- and tubule damage; and improved kidney function. The median
longed lifespan.6 However, if ramipril treatment of lifespan of Col4a32/2 mice was prolonged 22% by ramipril
Col4a32/2 mice was delayed until 7 weeks of age, after pro- alone, 44% with X203 alone, and 99% with ramipril1X203.
These data suggest that anti-IL-11 therapies hold promise for
teinuria was established, there was limited beneficial
treating kidney disease in AS.
effect.6,9 There are no specific or second-line medical thera-
pies for AS, and renal transplantation is the preferred treat-
ment for progressive CKD in AS.10 (13110; Cell Signaling Technology), a-smooth muscle actin
Kidney dysfunction in AS is initiated in the glomerulus, (19245; Cell Signaling Technology, Western blot [WB]),
related to altered GBM mechanics and podocyte dysfunc- SNAI1 (3879; Cell Signaling Technology, WB), p-STAT3
tion. However, as in other primary glomerular diseases, a (4113; Cell Signaling Technology), STAT3 (4904; Cell
major determinant of progressive kidney failure is in the Signaling Technology), TGF-b (3711; Cell Signaling Tech-
associated tubulointerstitial disease.4 Indeed, similar to nology), Wilms’ tumor 1 (WT1; ab89901; Abcam, immuno-
other forms of CKD, kidney function in AS patients corre- fluorescence [IF] and immunohistochemistry [IHC]), WT1
lates most strongly with the degree of tubulointerstitial (ab267377; Abcam, WB), anti-goat Alexa Fluor 488
fibrosis rather than glomerular pathology.11 Disease patho- (ab150129; Abcam), anti-rabbit Alexa Fluor 647 (ab150067;
genesis in AS is complex, involving the renin-angiotensin Abcam), anti-rabbit horseradish peroxidase (HRP; 7074;
system (RAS) and TGF-b activation, inflammation, partial Cell Signaling Technology), and anti-mouse HRP (7076;
epithelial-to-mesenchymal transition (pEMT) of tubular Cell Signaling Technology).
epithelial cells (TECs)/podocytes and fibrosis, among other
factors.9,12,13 Increasingly the role of pEMT, a failed-repair
proximal tubule cell (FR-PTC) state,14 is viewed as an initi- Ethics Statements
ating factor for renal fibrosis, inflammation, and failure, Animal studies were carried out in compliance with the
particularly because it prevents TEC proliferation and renal recommendations in the Guidelines on the Care and Use of
repair.15–20 Animals for Scientific Purposes of the National Advisory
Here, we investigated whether (1) IL-11, recently impli- Committee for Laboratory Animal Research. All experi-
cated as important for tubulointerstitial fibrosis and renal mental procedures were approved (SHS/2019/1482) and
dysfunction,21 is involved in the kidney pathology of AS conducted in accordance with the SingHealth Institutional
and (2) a neutralizing IL-11 antibody given to Col4a32/2 Animal Care and Use Committee.
mice with established renal disease and proteinuria could
improve molecular pathology, renal structure, and function,
and delay onset of death due to kidney failure. Mouse Model of Alport
Col4a32/2 (Col4a3tm1Dec) mice were purchased from the
Jackson Laboratory (https://www.jax.org/strain/002908). Mice
METHODS were housed at temperatures of 21 C–24 C with 40%–70%
humidity on a 12-hour/12-hour light/dark cycle and provided
Antibodies with food and water ad libitum. For the treatment study,
Cyclin D1 (55506; Cell Signaling Technology, Danvers, Col4a32/2 were administered 20 mg/kg of anti-I-L11 (3203)
MA), E-cadherin (3195; Cell Signaling Technology), p- or IgG isotype control (11E10) by intraperitoneal injection,
ERK1/2 (4370; Cell Signaling Technology), ERK1/2 (4695; starting from 6 weeks of age twice a week for 2.5 weeks;
Cell Signaling Technology), GAPDH (2118; Cell Signaling wild-type (WT) littermates were used as controls. Mice were
T), green fluorescent protein (GFP; ab6673; Abcam, Cam- euthanized for blood and kidney collection when they were 8.
bridge, UK), IgG (11E10; Aldevron, Fargo, ND), neutraliz- 5 weeks old. For the lifespan study, mice were intraperitone-
ing anti-IL-11 (X203; Aldevron), anti-IL-11RA (X209; ally administered either 3203 (twice a week, 20 mg/kg) or
Aldevron), NHPS2/podocin (ab181143; Abcam), PCNA 11E10 (twice a week, 20 mg/kg) alone or in combination
2 JASN JASN 33: –, 2022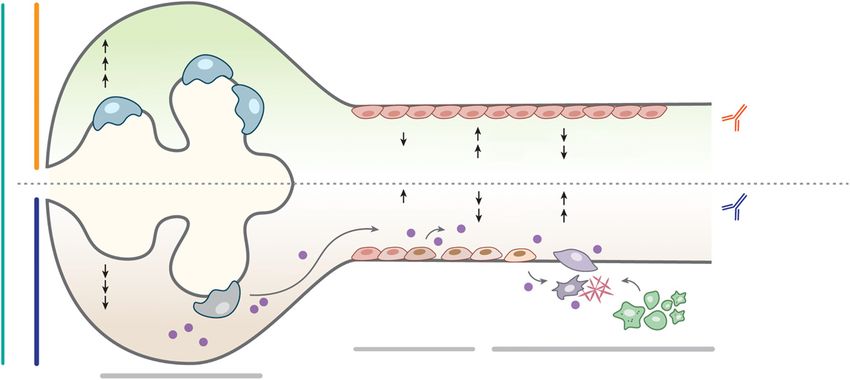
www.jasn.org BASIC RESEARCH
with ramipril (10 mg/kg; 6 days/week) starting from 6 weeks Histology
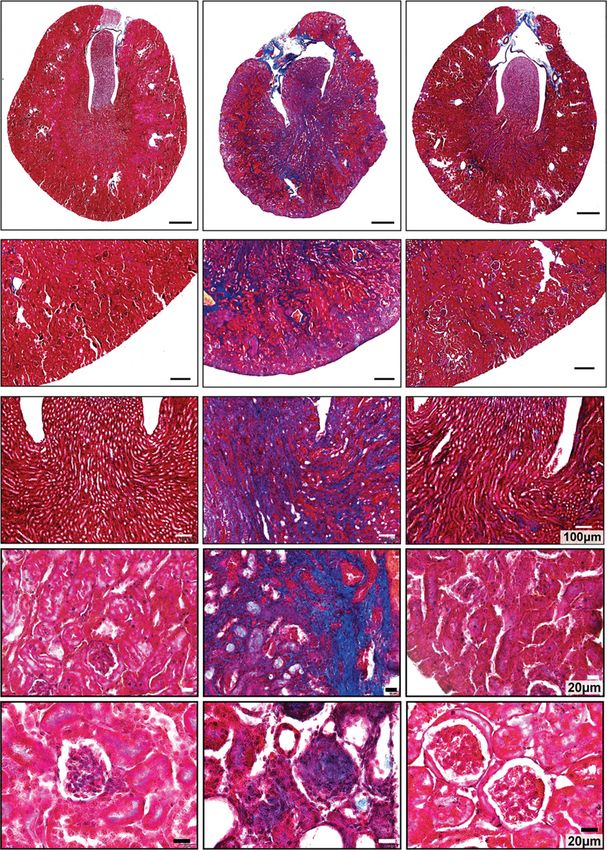
of age until death ensued. Kidney tissues were fixed for 48 hours at room temperature
in 10% neutral-buffered formalin, dehydrated, embedded in
paraffin, and sectioned at 7 mm. Transverse kidney sections
Col4a32/2-Il-11:EGFP were then stained with periodic acid–Schiff (PAS) and Mas-
Col4a32/2 mice were crossed to transgenic mice with
son’s trichrome according to standard protocol. Images of
EGFP constitutively knocked in to the Il-11 gene22 to gen-
the sections were captured by light microscopy, and blue-
erate hybrid cross of Col4a32/2-Il-11:EGFP1/2. Age-
stained fibrotic areas were semi-quantitatively determined
matched Col4a31/1-Il-11:EGFP1/2 littermates were used as
with ImageJ software (color deconvolution-Masson’s Tri-
controls. Mice were euthanized at 7.5 weeks of age; kidneys
chrome; National Institutes of Health, Bethesda, MD)
were excised and OCT embedded for IF staining.
from the whole kidney area (3100 field, n54 kidneys/
group). Kidney sections (n54–7/group) were indepen-
Western Blot (WB) dently scored for tubulointerstitial fibrosis (from Masson’s
WB was carried out on total protein extracts from mouse trichrome–stained kidney sections) and for glomeruloscle-
kidney tissues. Kidneys were lysed in radioimmunoprecipi- rosis and tubular atrophy (from PAS-stained kidney sec-
tation assay buffer containing protease and phosphatase tions) by a renal pathologist in a blinded fashion with the
inhibitors (Thermo Fisher Scientific, Waltham, MA), fol- following criteria:
lowed by centrifugation to clear the lysate. Protein concen- Interstitium (0, no fibrosis; 1, ,25% fibrosis; 2, 25%–50%
trations were determined by Bradford assay (Bio-Rad, fibrosis; 3, .50% fibrosis).
Hercules, CA). Protein lysates were separated by SDS- Glomeruli (0, no sclerosis; 1, ,25% sclerosis; 2, 25%–50%
PAGE, transferred to a polyvinylidene fluoride membrane, sclerosis; 3, .50% sclerosis).
and subjected to immunoblot analysis for various antibod- Tubules (0, no atrophy; 1, ,25% atrophy; 2, 25%–50%
ies (1:1000 in Tris-buffered saline with Tween 20) as out- atrophy; 3, .50% atrophy).
lined in the main text, figures, and/or figure legends. Total score is the sum of interstitial fibrosis score, glomer-
Proteins were visualized using the enhanced chemilumines- uli score, and tubule atrophy score.
cence detection system (Pierce, Rockford, IL) with the Treatment and genotypes were not disclosed to investi-
appropriate secondary antibodies: anti-rabbit HRP or anti- gators performing the histology and generating semi-
mouse HRP (1:2000 in Tris-buffered saline with Tween 20). quantitative readouts.
IHC
Quantitative PCR
Kidneys were fixed in 10% neutral-buffered formalin, paraf-
Total RNA was extracted from snap-frozen kidney tissues
finized, cut into 7-mm sections, incubated with primary
using Trizol (Invitrogen, Carlsbad, CA) followed by RNeasy
antibodies overnight, and visualized using the appropriate
column (Qiagen, Hilden, Germany) purification. cDNAs
ImmPRESS HRP IgG polymer detection kit: anti-rabbit
were synthesized with iScript cDNA synthesis kit (Bio-Rad)
(MP-7401; Vector Laboratories, Burlingame, CA), anti-
according to the manufacturer’s instructions. Gene expres-
mouse (MP-7402; Vector Laboratories) with ImmPACT
sion analysis was performed on duplicate samples with
DAB Peroxidase Substrate (SK-4105; Vector Laboratories).
either TaqMan (Applied Biosystems, Foster City, CA) or
Quantification of WT1ve cells were performed in a blinded
fast SYBR green (Qiagen) technology using StepOnePlus
fashion from four images (3200 field)/kidney (n53–4 kid-
(Applied Biosystem) over 40 cycles. Expression data were
neys/group).
normalized to GAPDH mRNA expression, and fold change
was calculated using the 2–DDCt method. The sequences of
specific TaqMan probes and SYBR green primers are avail- IF
able upon request. Kidneys were rinsed in cold PBS, patted dry with lint-free
paper, and cryo-molded in OCT compound (4583; Tissue-
Tek). After the OCT compound was frozen, kidney speci-
Colorimetric Assays mens were wrapped in aluminum foil and stored in –80 C.
The levels of BUN and creatinine (Cr) in mouse serum Cryo-embedded kidneys were cryosectioned (–20 C) at a
were measured using a Urea Assay Kit (ab83362; Abcam) thickness of 7 mm and allowed to dry on the slides for 1
and Creatinine Assay Kit (ab65340; Abcam), respectively. hour at room temperature. Kidney sections were fixed in
Urine albumin and Cr levels were measured using a Mouse cold acetone for 15 minutes before brief PBS washes, per-
Albumin ELISA kit (ab108792; Abcam) and Creatinine meabilized with 0.1% TritonX-100 (T8787; Sigma–Aldrich,
Assay Kit (ab204537; Abcam), respectively. All ELISA and St. Louis, MO), and blocked with 2.5% normal horse serum
colorimetric assays were performed according to the manu- (S-2012; Vector Laboratories) for 1 hour at room tempera-
facturer’s protocol. ture. Kidney sections were incubated with GFP and WT1
JASN 33: –, 2022 IL-11 Antibody and Alport Syndrome 3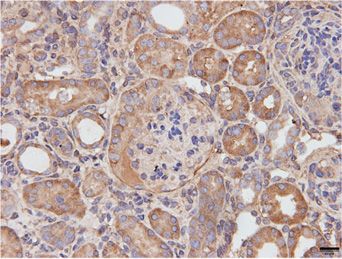
BASIC RESEARCH www.jasn.org
(1:500 in PBS containing 0.1% Tween 20) primary antibod- source of IL-11 better in the kidneys of Col4a32/2 mice,
ies overnight at 4 C, followed by incubation with the we crossed this strain with mice with EGFP knocked into
appropriate Alexa Fluor 488/647 secondary antibodies the Il-11 locus—the Il-11:EGFP mouse.22 Kidneys of
(1:250) for 1 hour at room temperature. DAPI was used to Col4a32/2-Il-11:EGFP1/2 mice exhibited large upregula-
stain the nuclei before imaging by fluorescence microscope tion of EGFP in the renal tubules and also showed EGFP
(Leica, Wetzlar, Germany). expression in the glomeruli, which co-localized with WT1
in podocytes (Figure 1G).
Statistical Analyses
Statistical analyses were performed using GraphPad Prism Antibody Neutralization of IL-11 Reduces Molecular
v8 (GraphPad Software, La Jolla, CA). Statistical signifi- Pathology in Col4a32/2 Mice
cance between control and experimental groups was ana- Over recent years, we have developed and characterized
lyzed by two-sided Student’s t tests or by one-way ANOVA antibodies that inhibit IL-11 signaling in murine and
as indicated in the figure legends. P values were corrected human cells.23,26 We administered one of these neutralizing
for multiple testing according to Tukey when several condi- IL-11 antibodies (X20326,27) or an IgG control antibody to
tions were compared with each other within one experi- 6-week-old Col4a32/2 mice—a time point when initiation
ment. Comparison analysis for two parameters from two of ramipril likely has limited effect.6 We examined renal
different groups was performed by two-way ANOVA. The pathologies in these mice 2.5 weeks after starting treatment,
criterion for statistical significance was P,0.05. when they are close to death due to renal failure, and com-
pared data with WT controls (Figure 2A).
At the end of the study period, total body weight loss, mea-
RESULTS sured as a percentage of their starting body weight, was signifi-
cantly attenuated in Col4a32/2 mice receiving 3203,
IL-11 Is Upregulated in the Kidneys of Col4a32/2 Mice compared with those administered IgG (IgG: 30%; X203:17%;
IL-11 is not expressed in normal healthy tissues, but its P50.0003; Figure 2B). As compared with Col4a32/2 mice
induction is commonly seen in fibroinflammatory dis- receiving IgG, X203-treated mice exhibited preserved kidney
eases.23 We profiled the Il-11 mRNA expression in kidneys mass (Figure 2C) and had significantly less kidney fibrosis by
of Col4a32/2 mice and found it to be upregulated both biochemical and histologic assessments (Figure 2, D–F).
(17.8-fold; P,0.001) compared with WT littermate controls Gene expression analyses showed renal levels of extracel-
(Figure 1A). We then assessed renal IL-11 expression at the lular matrix genes (Col1a1, Col1a2, Col3a1, and Fn), the
protein level in Col4a32/2 mice. IL-11 was not detectable myofibroblast marker Acta2, and pro-fibrotic factors (Il-11
at 4 weeks of age (or in 8.5-week-old WT mice), but it was and Tgfb1) were all reduced by 3203 compared with IgG
upregulated in the kidneys of 6-week-old mice (2.4-fold; (Figure 3A). The effect seen on transcript expression was
P50.04) and highly expressed by 8.5 weeks of age (5.9-fold; confirmed at the protein level for a-smooth muscle actin
P,0.001; Figure 1, B and C). and fibronectin (Figure 3B).
We sought to determine the target cells in the kidney At the signaling level, IL-11 is known to activate ERK
that express the IL-11 receptor (IL-11RA1) by both IHC across cell types, and this pathway has been mechanistically
and mining publicly available single-cell RNA sequencing linked with IL-11-driven fibrosis.26,28,29 IL-11 inhibition
data.24 In WT mice, IL-11RA1 expression was easily seen in vivo can also be associated with reduced STAT3 activa-
in tubules and in the glomerulus, whereas no staining was tion, which is thought to be largely a secondary phenome-
seen in sections from Il-11ra12/2 mice, confirming specific- non reflecting lesser stromal-driven inflammation.26,30
ity of detection (Figure 1D). In single-cell RNA sequencing Compared with WT mice, kidneys from Col4a32/2 mice
data from WT mice,24 we observed that Il-11ra1 and its treated with IgG exhibited elevated ERK and STAT3 activa-
partner receptor (gp130) were most highly expressed in tion. In contrast, ERK and STAT3 phosphorylation was
podocytes, collecting ducts with lesser expression in tubule largely diminished in the kidneys of 3203-treated
cells across the nephron, and fibroblasts (Figure 1E). Col4a32/2 mice (Figure 3, B and C). These data are consis-
IL-11RA1 expression, like IL-11, may be dynamically regu- tent with 3203 target engagement in the kidney, reduced
lated in disease, and we determined its expression in kid- ERK activation, and diminished inflammation.
neys from both WT and Col4a32/2 mice and found it to In many kidney diseases, it is thought that damaged
be upregulated in diseased kidneys, notably in the tubules TECs transition to a pEMT/FR-PCT state, which is central
(Figure 1F). to the subsequent development of tubulointerstitial fibrosis
IL-11 is secreted by epithelial cells exposed to infective and CKD.15–18 TEC pEMT/FR-PCT14 is characterized by
and toxic factors and in response to cytokines, chemokines, increased SNAI1 expression and reciprocal downregulation
and other signaling molecules.23,25 IL-11 is also secreted of E-cadherin that is regulated, in part, by TGF-b.15,16
from activated stromal cells across tissues. To define the Compared with WT controls, Col4a32/2 mice receiving
4 JASN JASN 33: –, 2022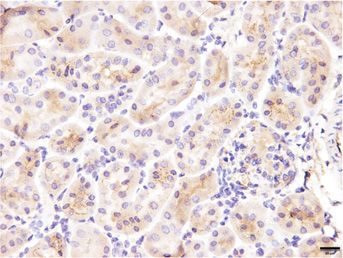
www.jasn.org BASIC RESEARCH
A II11/Gapdh mRNA (FC) B C
8BASIC RESEARCH www.jasn.org
www.jasn.org BASIC RESEARCH
IgG exhibited a strong molecular signature of EMT with in both podocytes and TECs.9,18 IHC analysis of the podo-
increased SNAI1 and decreased E-cadherin expression (Fig- cyte marker WT1 revealed greater staining in WT mice and
ure 3, B and C). In contrast, SNAI1 and E-cadherin levels 3203-treated Col4a32/2 mice compared with IgG-treated
in Col4a32/2 mice receiving 3203 were similar to those Col4a32/2 mice (Figure 4A). Quantification of the number
seen in WT mice. Thus, anti-IL-11 reduces TEC pEMT in of WT1-positive cells (podocytes) was carried out in a
the kidneys of Col4a32/2 mice. blinded fashion and confirmed significant (P,0.001) pres-
A specific feature of injured TECs that enter a pEMT/FR- ervation of podocyte integrity in Col4a32/2 mice receiving
PCT state is their inability to replicate—a process that relates to 3203 compared with Col4a32/2 mice receiving IgG (Figure
SNAI1 repression of cyclins D1/2 that blocks G1/S transi- 4B). Preservation of podocytes in 3203-treated Col4a32/2
tions.19,20,31 We profiled levels of cyclin D1 and those of mice was further ascertained by immunoblotting, and find-
PCNA, a marker of G1/S, in the kidneys of Col4a32/2 mice ings were extended to podocin, a second podocyte marker
treated with either IgG or 3203 and also in WT controls. (Figure 4, C and D).
Col4a32/2 mice receiving 3203 showed marked upregulation TGF-b upregulation in podocytes and tubular cells,
of cyclin D1 and PCNA compared with WT mice and which coincides with the onset of proteinuria in the
Col4a32/2 mice receiving IgG (Figure 3, B and C). This sug- Col4a32/2 mouse,9,32 is thought to be of importance for
gests anti-IL-11 inhibits pEMT/FR-PCT transitions in the kid- disease pathogenesis in AS. We thus examined TGF-b lev-
neys of Col4a32/2 mice and releases TECs to re-enter G1/S, to els and observed that 3203, but not IgG, significantly
replicate, and to repair damaged tubules. reduced the degree of TGF-b upregulation in the kidneys
of Col4a32/2 mice (Figure 4, C and D). Apoptosis of podo-
Podocyte Preservation and Lesser Renal Inflammation cytes and tubule cells is implicated in AS, and caspase
Is Associated with Inhibition of IL-11 Signaling in activity is reduced in Col4a32/2 mice given Olmesartan.32
Col4a32/2 Mice We observed caspase 3 activation in the IgG-treated
AS affects GBM composition, leading to podocyte dysfunc- Col4a32/2 mice that was reduced by 3203 administration
tion/loss that relates to TGF-b activity and pEMT processes (Figure 4, C and D).
A
0.0288 ns 0.0048
Rel. RNA expr. /Gapdh (FC)
Rel. RNA expr. /Gapdh (FC)
ns ns
80 25BASIC RESEARCH www.jasn.org
A Col4a3 –/– B
WT Control lgG X203
ns
150
0.0002
200X
WT1+ve cells
100
50
0
WT lgG X203
600X Col4a3 –/–
Wilms’ Tumor 1 staining
C Col4a3 –/– D
WT lgG X203
kDa
15 0.0060 ns
TGFE
Cl. CASP3/CASP3 (FC)
8
Podocin/GAPDH (FC)
20 1.5 0.0373 1.5
TGFE/GAPDH (FC)www.jasn.org BASIC RESEARCH
A B C D
2.0 2.0 2.0 4 ns
Glomerulosclerosis score
Interstitial fibrosis score
Tubular atrophy score
0.0167BASIC RESEARCH www.jasn.org
A B C
100
Probability of Survival (%)
No Treatment
lgG
death
0.0005www.jasn.org BASIC RESEARCH
Podocytes
Podocyn
WT1
Alport syndrome (Col4a3 –/–)
Anti IL11
Anti IL11
Proliferation Fibrosis
pEMT - Preserved renal function
Repair Immune cells
- Life extension
pEMT Proliferation Fibroblast activation
Control lgG
Repair Immune cell recruitment Control lgG
- Renal dysfunction / failure
- Early death
WT1 IL11
pSTAT3
Podocyn
Podocytes
Tubule Stress Stromal/ immune cell activation
Podocyte dysfunction
Figure 7. Proposed mechanism for IL-11-induced renal failure in AS. Glomerular basement membrane disruption due to Col4A3
mutation causes podocyte dysfunction and consequent tubular stress. Injured TECs, and podocytes, upregulate IL-11, leading to
autocrine pEMT (SNAI1 upregulation and E-cadherin downregulation) of TECs and podocytes and paracrine activation of stromal
myofibroblasts, which themselves secrete IL-11 that amplifies the fibrotic response and stimulates the recruitment/activation of
immune cells. Neutralizing antibodies against IL-11 reduce renal dysfunction and extend life in Col4a3-deleted mice.
weeks of age was more effective than ramipril alone, DISCLOSURES
increasing lifespan by 29 days (44%). Most notably, the
combination of anti-IL-11 and ramipril prolonged sur- S.A. Cook is a co-inventor of the patent applications WO/2017/103108
vival of Col4a32/2 mice by 62 days (99%), which is sub- (Treatment of Fibrosis), WO/2018/109174 (IL-11 Antibodies), and WO/
2018/109170 (IL-11RA Antibodies). S.A. Cook and A.A. Widjaja are
stantially longer than ACEi alone—the current standard co-inventors of the patent application US US2020/0270340A1 (Treatment
of care in AS patients. of Kidney Injury) and GB2009292.0 (Treatment and Prevention of Disease
Except for limited and incompletely penetrant develop- Caused by Type IV Collagen Dysfunction). S.A. Cook is a co-founder and
mental defects of teeth and skull sutures, humans with loss- shareholder of Enleofen Bio PTE Ltd., a company that made anti-IL-11
therapeutics, which were acquired for further development by Boehringer
of-function of IL-11RA appear well, with normal immune
Ingelheim in 2019. T. Coffman reports being on the editorial boards of
function. A similar phenotype is seen in Il-11ra1 null mice. Cell Metabolism and JCI and on the board of directors for Singapore
Interestingly, two recently and separately developed Il-11 Health Services, the Singapore Eye Research Institute, and the Kidney
null mice appear normal, with no obvious bony deficits,41,42 Research Institute University of Washington. P.H. Tan reports honoraria
suggesting that inhibiting IL-11 might have advantages in for delivering a talk on prostate cancer from AstraZeneca. A.A. Widjaja
reports patents and inventions with Boehringer Ingelheim. S.A. Cook
side-effect profile over targeting IL-11RA. Taken together, reports research funding from Boehringer Ingelheim. All remaining
the mild phenotypes of humans and mice lacking IL-11RA authors have nothing to disclose.
or IL-11, along with absence of untoward effects with
lengthy anti-IL-11RA and anti-IL-11 treatment in mice,26
provide an encouraging safety signal for long-term FUNDING
inhibition of IL-11 signaling in chronic diseases such as
AS.23,25 This research was supported by the National Medical Research Council
(NMRC), Singapore STaR awards (NMRC/STaR/0029/2017), NMRC Cen-
We end by suggesting that inhibition of IL-11 signaling
tre Grant to the NHCS, MOH-CIRG18nov-0002, MRC-LMS (UK), Tanoto
may be considered as a novel therapeutic approach for Foundation (to S.A. Cook). A.A. Widjaja is supported by the NMRC
patients with AS and perhaps other progressive forms of (NMRC/OFYIRG/0053/2017). N. H€ ubner and S.A. Cook are supported by
CKD. Anti-IL-11 therapy combined with RAS blockade a grant from the Leducq Foundation (16CVD03). N. H€ ubner is a recipient
may be of particular interest, given the near ubiquitous use of an ERC advanced grant under the European Union Horizon 2020
Research and Innovation Program (AdG788970).
of ACEi/ARB in CKD and the synergistic interaction
between anti-IL-11 and ACEi therapy shown here. With
anti-IL-11/anti-IL-11RA drugs nearing the clinic, it will be AUTHOR CONTRIBUTIONS
interesting to see if a therapeutic approach for AS discov-
ered in the Col4a32/2 mouse translates to patients for a E. Adami was responsible for visualization; T. Coffman, S.A. Cook, S.G.
second time.6,7 Shekeran, and A.A. Widjaja were responsible for the methodology;
JASN 33: –, 2022 IL-11 Antibody and Alport Syndrome 11BASIC RESEARCH www.jasn.org
T. Coffman, S.A. Cook, and A.A. Widjaja reviewed and edited the manu- mesenchymal transition drives renal fibrosis in mice and can be
script; S.A. Cook and N. H€ ubner were responsible for funding acquisition; targeted to reverse established disease. Nat Med 21: 989–997,
S.A. Cook, P.H. Tan, and A.A. Widjaja conducted the formal analysis; S.A. 2015
Cook and A.A. Widjaja conceptualized the study, were responsible for 16. Lovisa S, LeBleu VS, Tampe B, Sugimoto H, Vadnagara K, Carstens
supervision, and wrote the original draft of the manuscript; J.W.T. Goh, JL, et al: Epithelial-to-mesenchymal transition induces cell cycle
S.Y. Lim, S.G. Shekeran, J. Tan, P.H. Tan, S. Viswanathan, and A.A. Wid- arrest and parenchymal damage in renal fibrosis. Nat Med 21: 998–
jaja were responsible for the investigation; and S.G. Shekeran was respon- 1009, 2015
sible for validation. 17. Ying Q, Wu G: Molecular mechanisms involved in podocyte EMT
and concomitant diabetic kidney diseases: An update. Ren Fail 39:
474–483, 2017
REFERENCES 18. Liu Y: New insights into epithelial-mesenchymal transition in kidney
fibrosis. J Am Soc Nephrol 21: 212–222, 2010
1. Hudson BG, Tryggvason K, Sundaramoorthy M, Neilson EG: Alport’s 19. Chang-Panesso M, Humphreys BD: Cellular plasticity in kidney injury
syndrome, Goodpasture’s syndrome, and type IV collagen. N Engl and repair. Nat Rev Nephrol 13: 39–46, 2017
J Med 348: 2543–2556, 2003 20. Vega S, Morales AV, Oca~ na OH, Vald es F, Fabregat I, Nieto MA:
2. Hudson BG, Reeders ST, Tryggvason K: Type IV collagen: structure, Snail blocks the cell cycle and confers resistance to cell death.
gene organization, and role in human diseases. Molecular basis of Genes Dev 18: 1131–1143, 2004
Goodpasture and Alport syndromes and diffuse leiomyomatosis. 21. Schafer S, Viswanathan S, Widjaja AA, Lim W-W, Moreno-Moral A,
J Biol Chem 268: 26033–26036, 1993 DeLaughter DM, et al: IL-11 is a crucial determinant of cardiovascu-
3. Savige J: Alport syndrome: its effects on the glomerular filtration lar fibrosis. Nature 552: 110–115, 2017
barrier and implications for future treatment. J Physiol 592: 4013– 22. Widjaja AA, Dong J, Adami E, Viswanathan S, Ng B, Pakkiri LS, et al:
4023, 2014 Redefining IL11 as a regeneration-limiting hepatotoxin and thera-
4. Funk SD, Lin M-H, Miner JH: Alport syndrome and Pierson syn- peutic target in acetaminophen-induced liver injury. Sci Transl Med
drome: Diseases of the glomerular basement membrane. Matrix Biol 13: eaba8146, 2021
71–72: 250–261, 2018 23. Cook SA, Schafer S: Hiding in plain sight: Interleukin-11 emerges as
5. Jais JP, Knebelmann B, Giatras I, Marchi M, Rizzoni G, Renieri A, a master regulator of fibrosis, tissue integrity, and stromal inflamma-
et al: X-linked Alport syndrome: Natural history in 195 families and tion. Annu Rev Med 71: 263–276, 2020
genotype–phenotype correlations in males. J Am Soc Nephrol 11: 24. Park J, Shrestha R, Qiu C, Kondo A, Huang S, Werth M, et al: Sin-
649–657, 2000 gle-cell transcriptomics of the mouse kidney reveals potential cellu-
6. Gross O, Beirowski B, Koepke M-L, Kuck J, Reiner M, Addicks K, lar targets of kidney disease. Science 360: 758–763, 2018
et al: Preemptive ramipril therapy delays renal failure and reduces 25. Widjaja AA, Chothani SP, Cook SA: Different roles of interleukin 6
renal fibrosis in COL4A3-knockout mice with Alport syndrome. Kid- and interleukin 11 in the liver: Implications for therapy. Hum Vaccin
ney Int 63: 438–446, 2003 Immunother 16: 2357–2362, 2020
7. Gross O, T€ onshoff B, Weber LT, Pape L, Latta K, Fehrenbach H, et al; 26. Widjaja AA, Singh BK, Adami E, Viswanathan S, Dong J, D’Agostino
German Pediatric Nephrology (GPN) Study Group and EARLY PRO- GA, et al: Inhibiting interleukin 11 signaling reduces hepatocyte
TECT Alport Investigators: A multicenter, randomized, placebo- death and liver fibrosis, inflammation, and steatosis in mouse mod-
controlled, double-blind phase 3 trial with open-arm comparison els of nonalcoholic steatohepatitis. Gastroenterology 157: 777–
indicates safety and efficacy of nephroprotective therapy with rami- 792.e14, 2019
pril in children with Alport’s syndrome. Kidney Int 97: 1275–1286, 27. Ng B, Dong J, D’Agostino G, Viswanathan S, Widjaja AA, Lim W-W,
2020 et al: Interleukin-11 is a therapeutic target in idiopathic pulmonary
8. Gross O, Licht C, Anders HJ, Hoppe B, Beck B, T€ onshoff B, et al; fibrosis. Sci Transl Med 11: eaaw1237, 2019
Study Group Members of the Gesellschaft f€ ur P€
adiatrische Nephro- 28. Adami E, Viswanathan S, Widjaja AA, Ng B, Chothani S, Zhihao N,
logie: Early angiotensin-converting enzyme inhibition in Alport syn- et al: IL11 is elevated in systemic sclerosis and IL11-dependent ERK
drome delays renal failure and improves life expectancy. Kidney Int signalling underlies TGFb-mediated activation of dermal fibroblasts.
81: 494–501, 2012 Rheumatology (Oxford) 60: 5820–5826, 2021
9. Sayers R, Kalluri R, Rodgers KD, Shield CF, Meehan DT, Cosgrove 29. Widjaja AA, Viswanathan S, Jinrui D, Singh BK, Tan J, Wei Ting JG,
D: Role for transforming growth factor-beta1 in Alport renal disease et al: Molecular dissection of pro-fibrotic IL11 signaling in cardiac
progression. Kidney Int 56: 1662–1673, 1999 and pulmonary fibroblasts. Front Mol Biosci 8: 740650, 2021
10. Kashtan CE: Renal transplantation in patients with Alport syndrome: 30. Dong J, Viswanathan S, Adami E, Singh BK, Chothani SP, Ng B,
Patient selection, outcomes, and donor evaluation. Int J Nephrol et al: Hepatocyte-specific IL11 cis-signaling drives lipotoxicity and
Renovasc Dis 11: 267–270, 2018 underlies the transition from NAFLD to NASH. Nat Commun 12:
11. Hood JC, Dowling J, Bertram JF, Young RJ, Huxtable C, Robinson 66, 2021
W, et al: Correlation of histopathological features and renal impair- 31. Cano A, P erez-Moreno MA, Rodrigo I, Locascio A, Blanco MJ, del
ment in autosomal dominant Alport syndrome in Bull terriers. Neph- Barrio MG, et al: The transcription factor snail controls epithelial-
rol Dial Transplant 17: 1897–1908, 2002 mesenchymal transitions by repressing E-cadherin expression. Nat
12. Qi R, Yang C: Renal tubular epithelial cells: The neglected media- Cell Biol 2: 76–83, 2000
tor of tubulointerstitial fibrosis after injury. Cell Death Dis 9: 1126, 32. Suh SH, Choi HS, Kim CS, Kim IJ, Ma SK, Scholey JW, et al: Olme-
2018 sartan attenuates kidney fibrosis in a murine model of Alport syn-
13. Ryu M, Mulay SR, Miosge N, Gross O, Anders H-J: Tumour necrosis drome by suppressing tubular expression of TGFb. Int J Mol Sci 20:
factor-a drives Alport glomerulosclerosis in mice by promoting E3843, 2019
podocyte apoptosis. J Pathol 226: 120–131, 2012 33. Ninichuk V, Gross O, Reichel C, Khandoga A, Pawar RD, Ciubar R,
14. Kirita Y, Wu H, Uchimura K, Wilson PC, Humphreys BD: Cell profiling et al: Delayed chemokine receptor 1 blockade prolongs survival in
of mouse acute kidney injury reveals conserved cellular responses to collagen 4A3-deficient mice with Alport disease. J Am Soc Nephrol
injury. Proc Natl Acad Sci U S A 117: 15874–15883, 2020 16: 977–985, 2005
15. Grande MT, S anchez-Laorden B, L opez-Blau C, De Frutos CA, Bou- 34. Lim W-W, Corden B, Ng B, Vanezis K, D’Agostino G, Widjaja AA,
tet A, Ar evalo M, et al: Snail1-induced partial epithelial-to- et al: Interleukin-11 is important for vascular smooth muscle
12 JASN JASN 33: –, 2022www.jasn.org BASIC RESEARCH
phenotypic switching and aortic inflammation, fibrosis and remodel- 39. Su H, Lei C-T, Zhang C: Interleukin-6 signaling pathway and its role
ing in mouse models. Sci Rep 10: 17853, 2020 in kidney disease: An update. Front Immunol 8: 405, 2017
35. Simon-Tillaux N, Hertig A: Snail and kidney fibrosis. Nephrol Dial 40. Chien J-W, Chen W-L, Tsui Y-G, Lee M-C, Lin A-Y, Lin C-Y: Daily uri-
Transplant 32: 224–233, 2017 nary interleukin-11 excretion correlated with proteinuria in IgA
36. Shull MM, Ormsby I, Kier AB, Pawlowski S, Diebold RJ, Yin M, et al: nephropathy and lupus nephritis. Pediatr Nephrol 21: 490–496,
Targeted disruption of the mouse transforming growth factor-beta 1 2006
gene results in multifocal inflammatory disease. Nature 359: 693– 41. Nishina T, Deguchi Y, Ohshima D, Takeda W, Ohtsuka M, Shichino
699, 1992 S, et al: Interleukin-11-expressing fibroblasts have a unique gene
37. Strikoudis A, Cieslak A, Loffredo L, Chen Y-W, Patel N, Saqi A, et al: signature correlated with poor prognosis of colorectal cancer. Nat
Modeling of fibrotic lung disease using 3D organoids derived from Commun 12: 2281, 2021
human pluripotent stem cells. Cell Rep 27: 3709–3723.e5, 2019 42. Ng B, Widjaja AA, Viswanathan S, Dong J, Chothani SP, Lim S, et al:
38. Su J, Morgani SM, David CJ, Wang Q, Er EE, Huang Y-H, et al: Similarities and differences between IL11 and IL11RA1 knockout
TGF-b orchestrates fibrogenic and developmental EMTs via the RAS mice for lung fibro-inflammation, fertility and craniosynostosis. Sci
effector RREB1. Nature 577: 566–571, 2020 Rep 11: 14088, 2021
AFFILIATIONS
1
Cardiovascular and Metabolic Disorders Program, Duke-National University of Singapore Medical School, Singapore
2
Cardiovascular and Metabolic Sciences, Max Delbr€ uck Center for Molecular Medicine in the Helmholtz Association (MDC), Berlin, Germany
3
National Heart Research Institute Singapore, National Heart Centre Singapore, Singapore
4
Department of Anatomical Pathology, Singapore General Hospital, Singapore, Singapore
5
Department of Anatomy, Yong Loo Lin School of Medicine, National University of Singapore, Singapore, Singapore
6
DZHK (German Centre for Cardiovascular Research), Partner Site Berlin, Berlin, Germany
7
Charit
e-Universit€
atsmedizin, Berlin, Germany
8
MRC-London Institute of Medical Sciences, Hammersmith Hospital Campus, London, UK
JASN 33: –, 2022 IL-11 Antibody and Alport Syndrome 13You can also read

















































