The role of initial ecosystem state for future resilience and ecosystem outcome - Catarina Larsson
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The role of initial ecosystem state for future
resilience and ecosystem outcome
Catarina Larsson
Supervisors: Thorsten Blenckner and Susa Niiranen
Stockholm Resilience Centre, Stockholm University, Sweden
Master’s Thesis (60 ECTS)
2013-2014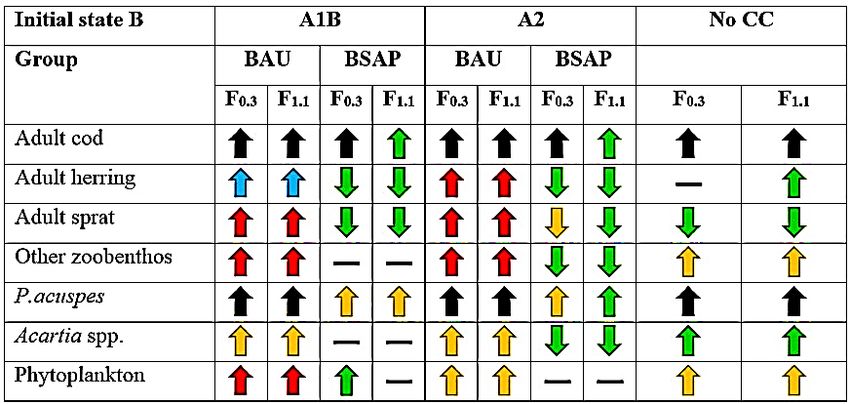
ABSTRACT Loss of marine ecosystem resilience can lead to abrupt changes in ecosystem structure, so called regime shifts, which affects dynamics and function of the ecosystem. Loss of ecosystem provisioning services can have a substantial impact on human well-being. Reversing an ecosystem back to its original state after a regime shift can be difficult, costly and impossible. Management strategies applied after a regime shift can have a different future outcome than applied before the regime shift. In the face of climate change and intensification of nutrient loading and fishing, there is a need to understand how to manage marine ecosystems in order to prevent future regime shifts and to understand the outcome from applied management strategies. The Central Baltic Sea is a heterogeneous water body with hydrographical differences from the south to the north, and with Gotland basin suffering with hypoxic conditions and summer algal blooms and it provides an interesting opportunity to explore ecosystem response on a smaller spatial scale. Here I combine ecosystem modeling and future scenarios to explore past ecosystem dynamics and by testing future management strategies from two initial states explore the ecosystem outcome and resilience. The present study have successfully constructed an Ecopath with Ecosim food web model of Gotland basin from 1980 to 2009 where the results shows that past, i.e., 1980-2009 dynamics were in line with the changes occurring on a larger scale, the Central Baltic Sea. Results from future projections show that the level of nutrients has a large impact on the future outcome of the ecosystem and the influence initial ecosystem states have for future outcome is found to be low. Finally, I discuss drivers and processes behind the past changes and future projections in Gotland food web, bringing insights to the role of initial ecosystem state for future ecosystem- based management.
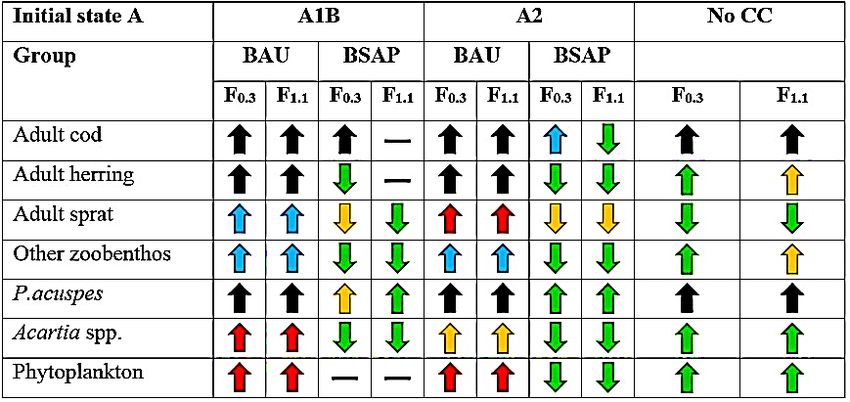
Table of Contents
INTRODUCTION ................................................................................................................................... 1
MATERIAL AND METHODS .............................................................................................................. 8
Case study area – Central Baltic Sea – Gotland basin ....................................................................... 8
Construction of Gotland Ecopath food web model ........................................................................... 10
Input data for Gotland Ecopath food web model .............................................................................. 12
Time-dynamic Ecosim model of Gotland food web ........................................................................... 13
Calibration and forcing data for Ecosim model of Gotland food web .............................................. 14
Calibration of Gotland food web model ............................................................................................ 16
Fitting procedure of Gotland food web model .................................................................................. 16
Future scenarios ................................................................................................................................ 17
Analysis of the model results – principle component analysis .......................................................... 20
Analysis of the model results – STARS .............................................................................................. 21
RESULTS.............................................................................................................................................. 22
Model performance ........................................................................................................................... 22
Model fit and validation ................................................................................................................ 22
Estimated biomass trends for Gotland food web from 1980 to 2009 ............................................ 26
Detected regime shifts in Gotland food web model ...................................................................... 27
Future scenario projections .............................................................................................................. 28
Trends in future biomass from two different initial states ............................................................. 28
Detected regime shifts from two different initial states ................................................................ 30
DISCUSSION ....................................................................................................................................... 33
Method and material discussions – limitations of the method .......................................................... 33
Gotland basin-past dynamics from 1980 to 2009.............................................................................. 35
Global and regional drivers of future ecosystem outcome ................................................................ 36
The role of initial ecosystem states for future resilience and management ....................................... 39
Conclusions and perspective for future studies ................................................................................. 42
ACKNOWLEDGEMENTS .................................................................................................................. 43
LITERATURE CITED.......................................................................................................................... 44
APPENDIX ........................................................................................................................................... 53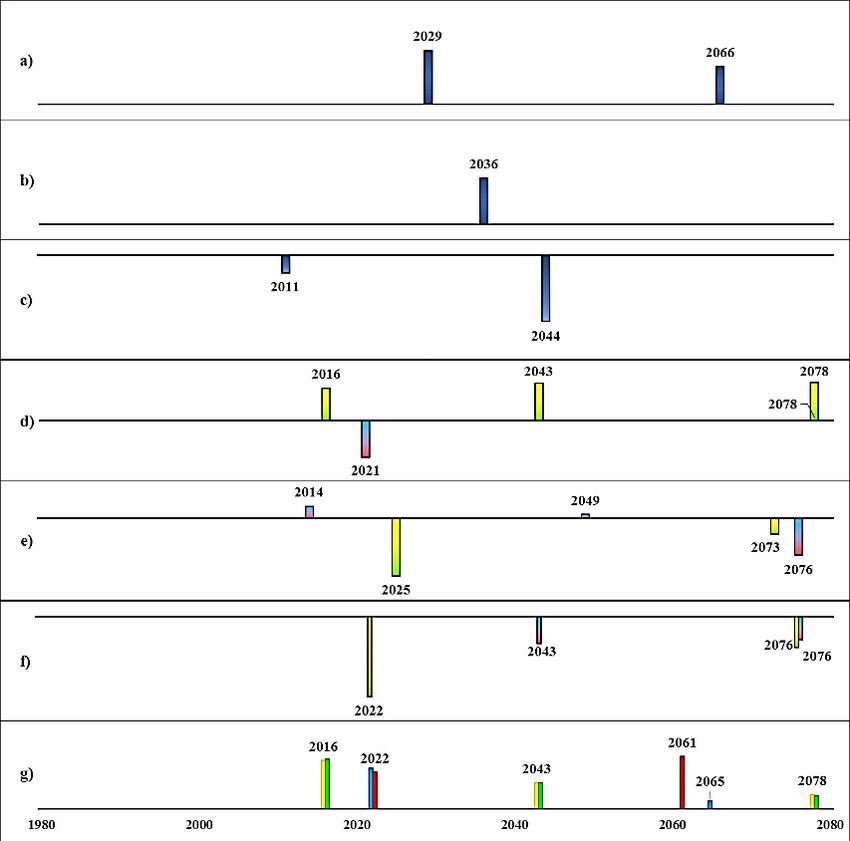
INTRODUCTION
Marine ecosystems worldwide have undergone large changes under the past decades. Events
such as eutrophication (Smith et al. 1999), jellyfish overtake (Cury & Shannon 2004) and fish
stock collapse (Myers & Worm 2003) are some examples of drastic changes in marine
ecosystem dynamics. Even if causes and drivers of change differ between ecosystems, climate
fluctuation and change have been documented to be closely coupled with changes in ecosystem
dynamics (Hoegh-Guldberg & Bruno 2010). An example of the close coupling is the change in
Northern Atlantic Oscillation (NAO) in late 1980s, which resulted in a synchronous change in
ecosystem dynamics in the North Sea, Northeast Atlantic and the Central Baltic Sea (Alheit et
al. 2005; Drinkwater et al. 2000). In addition to changes in climate, more direct human stressors
are exerting a high overall pressure on marine ecosystems due to its provisioning services of,
e.g., food, transportation, recreation and a recipient of nutrient and pollution run-off (Halpern
et al. 2008; Micheli et al. 2013).
The combination of large-scale (or global) external drivers, such as climate and regional human
stressors have caused several marine ecosystems to change their dynamics and internal
processes, resulting in ecological regime shifts (Lees et al. 2006). For example, a shift from
planktivorous fish dominated state to a jellyfish dominated state in the Northern Benguela (Cury
& Shannon 2004), the shift from a cod (Gadus morhua) to clupeid (i.e., sprat (Sprattus
sprattus)) dominance in the Central Baltic Sea (Möllmann et al. 2009) and the climate induced
trophic reorganization in the Gulf of Alaska (Anderson & Piatt 1999) are some of the marine
ecosystem regime shifts documented. According to the definition, e.g., by Holling (1973) and
Scheffer & Carpenter (2003) an ecological regime shift (further called regime shift) takes place
when a system is pushed from one stable state to an alternative state thus changing the
ecosystem dynamics, feedbacks and function. The change in dynamics and function might
result in that the new regime responds differently to management of, e.g., nutrients, than the old
regime would had (Folke et al. 2004). Large-scale global forces, e.g., climate, can trigger
regime shifts on a local level by affecting the internal feedback processes
that retain the system in its stable state (Scheffer et al. 2001; Scheffer & Carpenter 2003).
1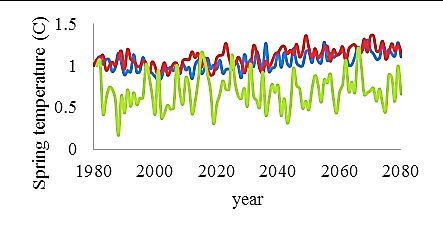
Ecological resilience is an ecosystem feature that determines a system’s ability to absorb
disturbances and changes without crossing a threshold and shifting into another state, i.e.,
undergoing a regime shift (Holling 1973; Gunderson 2000). Once a system has crossed a
threshold and moved into a new alternative state, the process of going back to the original
regime can be difficult due to the new dynamics and feedback processes, which keeps the
system locked into the new regime (Nyström et al. 2012; Scheffer et al. 2001).
High species diversity, multiple functional groups and high response diversity are found to
enhance the resilience of ecosystems (Folke et al. 2004). Resilience can change over time as
more perturbations are affecting the ecosystem dynamics. For example, intense local fishing
resulting in the removal of top predators, and hence causing a change in food web structure,
can make the system more sensitive to changes in climate and hydrography (Llope et al. 2011;
Österblom et al. 2010). Change in climate itself can affect key species such that the whole
community changes in character and subsequently the function of the ecosystem changes
(Schiel et al. 2004; Harley et al. 2006).
The Black Sea is an example of how the resilience of a marine ecosystem can be affected by
the interactive effect of multiple drivers and the difficulty to restore the resilience of the system
once it has shifted into another regime. The Black Sea has gone through two regime shifts in
the late 20th century. The first shift was in the 1970s when the ecosystem shifted from a state of
high abundance of large predator fish and high zooplankton abundance, to an ecosystem state
of high abundance of small pelagic fish species and an increasing biomass of phytoplankton
(Oguz & Gilbert 2007).
The second regime shift was in the late 1980s was due to the top-down stressor, i.e., fishing out
pelagic top predators in combination with bottom-up drivers, i.e., nutrient enrichment and
warmer climate, which resulted in a system state favorable for invasive species and
phytoplankton blooms (Daskalov 2003; Llope et al. 2011). This resulted in a decrease
abundance of small pelagic fish while the biomass of gelatinous carnivores, i.e., jellyfish
increased and became the prevalent predator of the system, resulting in a steep decline of main
food source zooplankton. The decrease of zooplankton lead to a decrease in grazing of
phytoplankton and with an increase of nutrients into the system, the Black Sea experienced
massive phytoplankton blooms in early 1990s (Oguz & Gilbert 2007; Daskalov 2003).
2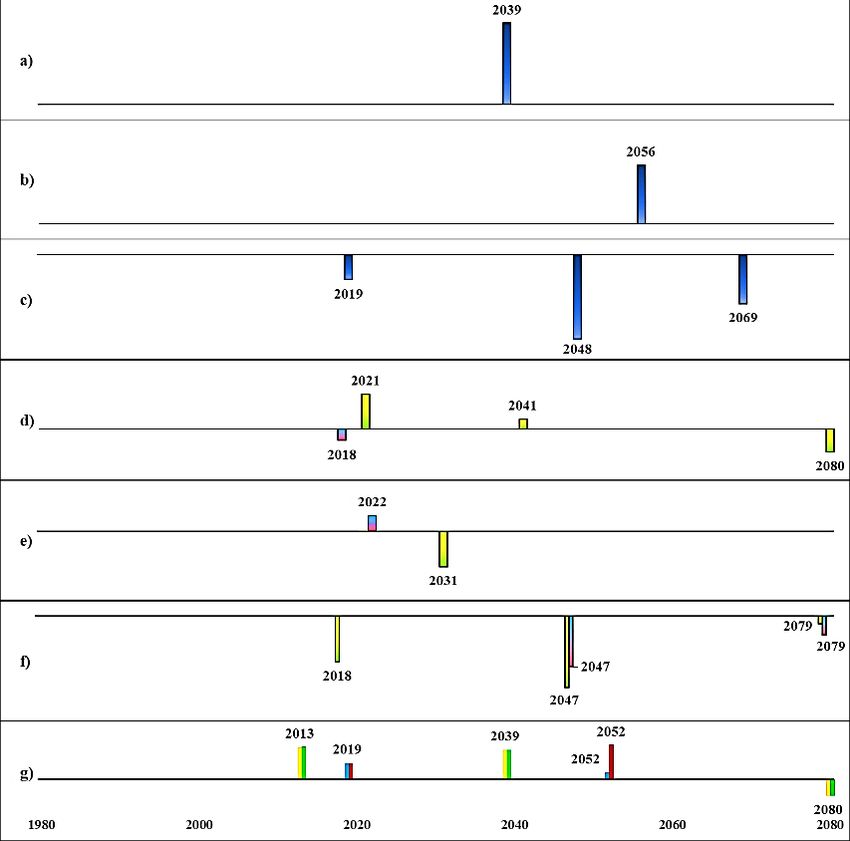
The Black Sea ecosystem has displayed a potential of partial recovery (Lancelot et al. 2002)
through the choice of management strategy, mainly management of nutrient loading and fishing
pressure, is vital in the Black Sea ecosystem recovery and restoration of resilience in the system
(Langmead et al. 2009). However, there are no indications of the Black Sea returning to the
original state pre-1960s, due to the establishment of jellyfish (Mnemiopsis leidyi) (Llope et al.
2011). These findings indicates the need for more knowledge on the interactive effects of
bottom-up and top-down drivers on the future resilience of marine ecosystems, how thresholds
function and how to manage the ecosystem for the future.
The Central Baltic Sea (CBS) has several common attributes with the Black Sea, change in
hydrography, nutrient increase and high fishing pressure are all drivers of the ‘discontinuous
regime shift’ in ecosystem structure that has been documented for the late 1980s (Möllmann et
al. 2009). The regime shift in the CBS was a reorganization within the trophic levels of fish,
mainly the two key commercial fish groups: the piscivorous cod and planktivorous sprat and
within the trophic level of zooplankton from P. acuspes domination to Acartia. spp. and
Temora. sp. (Alheit et al. 2005). In the mid-1970s, the hydrographical conditions, i.e., high
salinity and high oxygen levels were preferable for cod recruitment and P. acuspes resulting in
high cod recruitment and P. acuspes abundance (Mackenzie et al. 2007; Möllmann et al. 2009).
At the time, the fishing pressure was high on cod, the main fish predator in the system, which
controlled the abundance of sprat by predation and the abundance of P. acuspes was mainly
controlled by the level of salinity (Casini et al. 2009). The hydrography changed over time, the
temperature increased while salinity and oxygen decreased, leading to a decrease in cod
offspring survival and less preferable conditions for P. acuspes.
The bottom-up changes (i.e., hydrographic conditions) combined with a too high cod
fishing pressure in relation to the low reproductive success, resulted in steep decline in cod
stock between late 1980s and mid-1990s (Möllmann et al. 2009; Köster et al. 2005).
Temperature increased during the ‘transition period’ (i.e., late 1980s to mid-1990s) which was
preferable to zooplankton Acartia spp., Temora sp. (Möllmann et al. 2000; Möllmann et al.
2009) and sprat had a high reproductive success, resulting in an high abundant sprat stock
(Möllmann et al. 2008; Köster et al. 2003).
3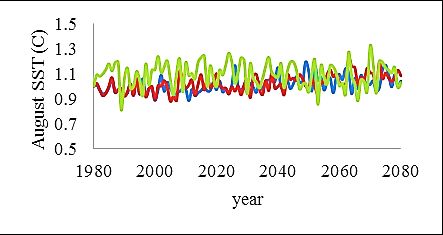
The high abundant sprat resulted in an increased predation pressure on cod egg after the
‘transition period’ (Köster & Möllmann 2000) and Casini et al. (2009) suggested that the high
sprat abundance also resulted in a decoupling of the link between P. acuspes and salinity, thus
shifting P.acuspes from being salinity controlled to sprat controlled. The same type of
decoupling have also been suggested occurring between cod recruitment and cod reproductive
volume and thereby indicating a shift in the main driver of cod recruitment (Casini et al. 2009).
Sprat continued being the abundant fish species in the system after the regime shift due to,
among other, the continued high cod fishing pressure and a failure for cod and P. acuspes to
recover in 1993 and 2003 when large inflow events of saline water from the North Sea were
observed (Möllmann et al. 2009).
The regime shift in the Central Baltic Sea was caused by unfavorable recruitment conditions
for cod (i.e., low salinity and oxygen depletion) combined with high cod fishing pressure
(Möllmann et al. 2009). In order to stabilize the Central Baltic Sea ecosystem for the future, a
combination of reduced nutrient loading and cod fishing pressure have been suggested (Casini
et al. 2008; Möllmann et al. 2009; Eero et al. 2012).
Currently, modeling is a widely used tool to create understanding of food web dynamics
under different future environmental scenarios and management strategies (Scheffer et al. 2001;
Harvey & Cox 2003; Meier et al. 2012). In the Baltic Sea for example, the BaltProWeb model,
a food web model of the Central Baltic Sea, has been used to create deeper understating of the
Central Baltic Sea ecosystem and trophic flows between multiple functional groups (Tomczak
et al. 2012). The BaltProWeb was recently used by Niiranen et al. (2013a) to study the food
web response to combined effects of global, i.e., climate and regional, i.e., nutrient loading and
fishing mortality, stressors. The study established that both global and regional drivers are
important to consider since both affect the future of the Central Baltic Sea food web (Niiranen
et al. 2013a).
Since the CBS ecosystem spans over a large area (2.4 x 105 km2; Tomczak et al. (2012))
where the hydrographical conditions, e.g., salinity levels, differ between the southern, northern,
eastern and western parts as well as coastal waters and the open sea (Leppäranta & Myrberg
2009), the BaltProWeb model might miss spatial differences in ecosystem response between
basins.
4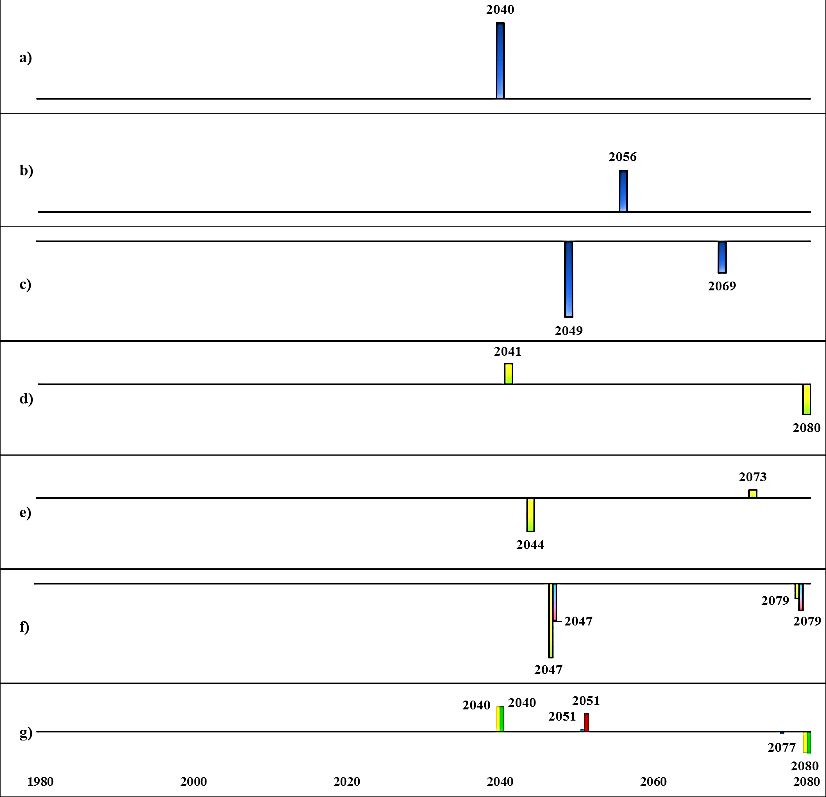
For example, cod spawning is mainly located in Bornholm basin (Köster et al. 2005;
Köster et al. 2001) while cod spawning in Gotland basin has been eradicated (Plikshs et al.
1999), mainly due to Gotland basin suffering from low oxygen and salinity levels resulting in
a non-successful cod spawning conditions (Köster et al. 2005). Phytoplankton distribution is
another factor where Bornholm basin display a decreasing trend while Gotland basin display
an opposing trend (Wasmund & Uhlig 2003). Thus, the Central Baltic Sea cannot be considered
a homogenous water body and with indications of ecosystem dynamics and environmental
conditions differing between basins within the CBS (MacKenzie et al. 2000; Köster et al. 2001;
pers. comm. Blenckner 2013), there is a need for a smaller spatial scale model.
For this thesis, the ecosystem of Gotland basin is the area in which the study is carried
out. Gotland basin is under a high anthropogenic pressure, extraction of fish by trawling and
nutrient enrichment from, e.g., agriculture and industries, are main drivers of change in Gotland
basin, resulting in bottom-up and top-down changes of the ecosystem structure (HELCOM
2010). Presently, there are two management strategies in place, which targets human behavior
in and around the Baltic Sea in an effort of reducing the risk of further changes in ecosystem
structure.
The Baltic Sea Action Plan (BSAP) aims at reaching a good environmental status by
2021 by reducing nutrient loading, especially nitrogen and phosphorus to the Baltic Sea
(HELCOM 2007). Recent studies have found that reduction in nutrient loading according to the
BSAP could result in higher oxygen concentrations, less sedimentation and an increase in
benthic biomass (Morf 2012; Timmermann et al. 2012). The action plan also addresses the
problem of shipping, hazardous substances and biodiversity conservation (HELCOM 2007) .
In order to facilitate the recovery of the Baltic cod stock, the European Council developed the
Cod Recovery Plan (CRP) comprised of actions to reduce cod fishing mortality and regulate
total allowable catch (EC 2007). The CRP sets the level of fishing mortality in the CBS to 0.3
for adult cod (ages 4 to 7 years), and a review of the recovery of the Eastern Baltic cod showed
that the low fishing mortality is one factor in the recent recovery of the Baltic cod stock (Eero
et al. 2012). In addition, OCEANA (2014), an organization solely focused on ocean
conservation and protection of marine ecosystems, has proposed the implementation of a
Marine Protected Area (MPA) in Gotland basin, outside the east coast of Gotland Island, and it
would cover approx. 6.8 % of Gotland basin (OCEANA 2014).
5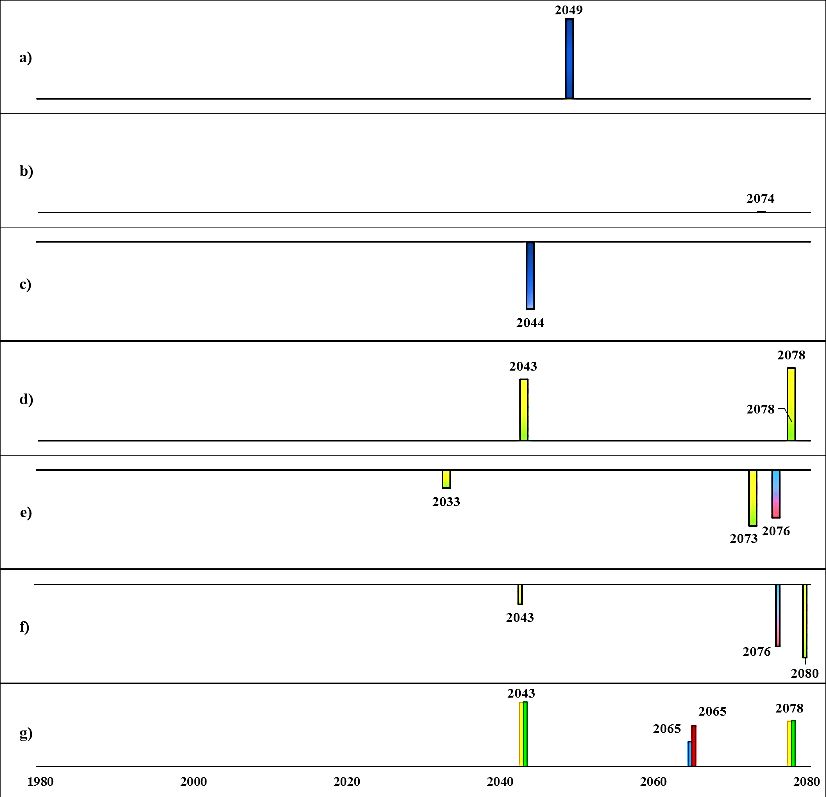
The suggested management strategy for the MPA is to tackle the two main threats to the area,
eutrophication and fishing, in order to deal with the, e.g., reduced cod spawning (OCEANA
2014). Finding management strategies that, not only, can manage global (i.e., climate change)
and regional anthropogenic (i.e., nutrient loading and cod fishing) interactions, i.e. ecosystem-
based management (Lindegren et al. 2010; McLeod et al. 2005), though also strategies that can
create a more resilience ecosystem towards future regime shifts is important (Thrush et al.
2009). This thesis will explore the future impact of the proposed management strategy by
OCEANA (2014) in Gotland basin.
The main aim of this thesis is to build a food web model for the Gotland basin to better
understand the past and potential future dynamics caused by changes in climate, nutrient and
fishing. All future nutrient-cod fishing scenarios will be explored in three possible future
climate scenarios to understand how the global driver of climate change could affect the future
of management strategies on a smaller spatial scale.
Scenarios will be applied to the model to explore potential future ecosystem response to a
hypothesized implementation of a Marine Protected Area in the whole area of Gotland basin,
with restricted cod fishing in accordance with the Cod Recovery Plan and a reduction of
nutrients by the use of the Baltic Sea Action Plan. The null hypothesis is a future of continued
eutrophication (business as usual) and a high cod fishing pressure.
In addition, different combinations of nutrient-cod fishing pressure will be tested to explore if
there are other types of management strategies, which may result in a favorable ecosystem with
less regime shifts and a high resilience. Therefore, the scenario modeling will be applied using
two different alternative states to explore the influence of the initial state on the ecosystem
outcome of the management strategies. The initial states tested are based on the past cod
dominated state and the today’s sprat dominated state. See conceptual figure (Fig. 1) for future
management combinations tested from two initial ecosystem states and the overall structure of
the thesis.
6
Fig. 1. Conceptual figure of thesis structure.
The figure displays the process in which the thesis has been conducted. Steps 1-11 represents different stages of the methodology and
analysis and are presented in “Material and Methods”.
Below step 8. are the tested nutrient-fishing scenarios (BAU: Business as usual, BSAP: Baltic Sea Action Plan
with fishing mortality: F0.3: Cod Recovery Plan: F1.1: “high cod fishing pressure) under climate scenario, A1B, A2
and ‘No CC (No further climate change), from initial ecosystem state A (i.e., today’s sprat dominated state) and initial ecosystem state B
(past cod dominated state).
7MATERIAL AND METHODS
Case study area – Central Baltic Sea – Gotland basin
The semi-enclosed Baltic Sea is one of the
largest bodies of brackish water in the world
(ICES 2008b) and it has a large catchment area
with over 85 million inhabitants living within
it (HELCOM 2010). The brackish water is not
an optimum for either marine nor freshwater
species, hence the relatively low species
richness (cf Elmgren and Hill 1997; Möllmann
et al. 2008).
Fig. 2. The Baltic Sea with ICES sub-divisions,
where area 28-2 is Gotland basin, the case study area.
(© FAO 1990-2014)
The three main fish species are Baltic cod (Gadus morhua), sprat (Sprattus sprattus) and herring
(Clupea harengus). They dominate the total fish biomass with 80 % (Elmgren 1984), and
despite low species diversity the system provides both socially and economically important
services, e.g., fishing, transportation and tourism (HELCOM 2010).
Over the last decades the Baltic Sea has experienced changes in nutrient loadings (Wulff et al.
1994; Fleming-Lehtinen et al. 2008), high overall fishing pressure (Elmgren 1989; Nyström et
al. 2012) and change in climate (MacKenzie & Köster 2004; Möllmann et al. 2008), which have
pushed the ecosystem to change its dynamics (HELCOM 2010; Korpinen et al. 2012). The
Central Baltic Sea (CBS) has some special oceanographic and geographical characteristics: i) a
salinity gradient that stretches from the southwest (10 psu) to the northeast (6 psu) (ICES
2008b) ii) irregular inflows of saltwater from the North Sea. The inflow brings salty, cold and
oxygenated water to the CBS and thereby oxygenates the deep basins (ICES 2008b). Larger
inflows have occurred two times since the 1980s one in 1993 and again in 2002-2003 (Fonselius
& Valderrama 2003; Lehmann et al. 2004) iii) a high level of hypoxia due to eutrophication in
the Central Baltic Sea (Cloern 2001)
8The Central Baltic Sea has been divided into five basins according to the International
Council for the Exploration of the Sea (ICES): sub-divisions: Bornholm (no. 25), Gdansk deep
(no. 26) and Gotland (no. 28-2) are the three main basin in the CBS (Fig. 2).
The case study area, Gotland basin (no. 28-2) is located in the middle of the Central Baltic Sea
and have an area of 39 380 km2 and an average depth of 87 m (Pers. comm. by Saskia Otto).
Characteristics of Gotland basin are algal blooms during warm summer months (Casini et al.
2008), eutrophication and oxygen depletion are all effects of nutrient enrichment and have
under recent decades increased in frequency and intensity in Gotland basin (HELCOM 2010;
HELCOM 2013; Hansson & Andersson 2013). Gotland basin has a permanent halocline 60-80
m, restricting the water exchange within the basin, creating a continuous hypoxic condition (the
deepest parts of the basin with a permanent anoxia) (Leppäranta & Myrberg 2009; Conley et
al. 2002). The salinity level is lower in Gotland basin than in the southern Baltic, resulting in
for example, a lower number of benthic species than in the southern Baltic where there the
salinity level is higher (cf Elmgren & Hill 1997). Modeling of future climate change projects
salinity levels and oxygen levels to decrease in Gotland basin (HELCOM 2013; Meier et al.
2012), thus negatively affect the survival conditions for, e.g., cod and P. acuspes hence the risk
for further regime shifts in the future.
9Construction of Gotland Ecopath food web model
Step 1. The software Ecopath with Ecosim (EwE) (Ver. 6.4; Pauly, Christensen, and Walters
2000; Christensen and Walters 2004; Christensen et al. 2008) was used to describe Gotland
food web for the baseline year 1980. The core equation (Eq. 1) of Ecopath for a trophic group
i, in an ecosystem consistent of n trophic groups:
Bi ∙ (P/B)i = ∑j Bj ∙ (Q/B)j ∙ DCij + Yi + Ei + BAi + Bi ∙ (P/B) (1-EEi) (Eq. 1)
Bi – biomass of group (i) in the ecosystem, Pi – total production of trophic group i
Qi – total consumption of trophic group j, DCij – the proportion of i in diet of group j.
Note, annual production rate (P/B) for group (i) is considered to be equivalent to total mortality
(Zi) for models in a steady state (Allen 1971). Thus, total mortality has been used to estimate
P/B for each fish group.
EwE defines production; Pi, for a trophic group (i) by components
i) predation mortality, i.e., the consumption of biomass by prey
(∑( Bj ∙ (Q/B)j ∙ DCij))
ii) export from the ecosystem by fisheries (Yi) and by net migration (Ei)
iii) biomass accumulation rate of (i) (BAi)
iv) other mortality (1-EEi), i.e., mortality that is not captured by the model e.g.
starvation, disease, etc. In Ecopath this is entered as 1 - Ecotrophic Efficiency (EEi).
EE is the proportion of production that is consumed within or caught from the
ecosystem. (Pauly et al. 2000; Christensen et al. 2008).
10Fig. 3. All 21 trophic groups of Gotland food web model. ‘Phytoplankton’ represents both summer and spring phytoplabkton. The lines
represents the trophic flow between groups.
The Ecopath model developed for this thesis described the food web of the Gotland basin in
1980 and consisted of 21 functional groups, from primary producers to top predator cod (Fig.
3). Selection of functional groups was based on their abundance and importance as indicated
by previous studies from the region (Harvey & Cox 2003; Österblom et al. 2007; Tomczak et
al. 2012; Casini et al. 2008). The model included two detritus groups, sediment (detritus (s))
and water-column (detritus (w)). Primary production was represented by spring and summer
phytoplankton. Microzooplankton is a vital part of the microbial loop hence it was included in
the mass-balance model (Calbet 2008).
Mesozooplankton was divided into four functional groups: Acartia spp. (Acartia sp.), Temora
sp. (Temora sp.), P.acuspes (Pseudocalanus sp.) and other zooplankton. Other zooplankton
consisted mostly of cladocerans, e.g., Podon intermedius, Evadne nordmanii and Bosmina
coregoni maritime (hence this group is called Cladocerans). Mysids is an important food item
for the fish community and was therefore identified as a separate functional group in the food
web (Dziaduch 2011).
11Three functional groups represented the benthic community, Monoporeia/Pontoporeia affinis
(Monoporeia/Pontoporeia), Saduria entomon (Saduria) and other zoobenthos, i.e.,
macrozoobenthos. The functional groups of fish - cod, herring and sprat were the three main
top predators of the food web. Each fish group was divided into multiple stanzas (age groups)
in order to represent the ontogenetic shifts in diet composition.
The division of stanzas were made based on the Tomczak et al. (2012) BaltProWeb model
(Appendix: Table A1), where the procedure of stanza division was based on Walters, Martell,
and Mahmoudi (2006).
Input data for Gotland Ecopath food web model
Step 2. Observation data for the baseline model of Gotland food web was collected from
ecological monitoring, fish stock assessments and literature, (Appendix: Table A2). In cases
where neither ecological studies nor literature could provide necessary data, discussions with
Blenckner, Niiranen and Otto (2013) were held in order to estimate the data missing.
Biomass (B), production per biomass rate (P/B), production per consumption (P/Q) and diet
composition (DC) (for diet proportions see Appendix: Table A3) were provided for functional
groups: bacteria, Monoporeia/Pontoporeia, Saduria and other zoobenthos. The functional
groups: microzooplankton, Acartia spp., Temora sp., P. acuspes and mysids were provided with
B, P/B, consumption per biomass (Q/B) and DC.
For fish groups were B and Q/B provided for the leading stanzas, i.e., adult (ad.) sprat, ad.
herring and ad. cod, and total mortality (Z) for all stanzas. Catch was provided for ad. cod,
small. cod, ad herring, juv. herring, ad. sprat and juv. sprat. Furthermore, ecological parameters
k-von Bertalanffy growth function (von Bertalanffy 1938) and the ratio of the weight at maturity
to the weight at infinity (Wmat/Winf ) were given to each fish group (Appendix: Table A4). The
final balanced Gotland food web baseline model, i.e., Ecopath model with observation data and
model estimates (Appendix Table A5) was the base for the next step; the time-dynamic Ecosim
model.
12Time-dynamic Ecosim model of Gotland food web
Step 3. Ecosim (Walters et al. 1997) was used to explore Gotland food web temporal dynamics
from 1980 to 2009 and in testing future management scenarios. Ecosim builds on several
equations from Ecopath and explains the rate of change in biomass (Eq. 2) (Christensen et al.
2008)
dBi / dt = P/Qi ∙ ∑j Qij - ∑j Qij + Ii – (MOi + Fi + ei) ∙ Bi (Eq. 2)
where dBi / dt represent the rate of biomass change (Bi) of group (i) during the time interval dt.
Production per consumption of group (i) (P/Qi) is followed by two summations i) the first
represent total consumption by group (i) ii) the second is the predation of all predators on the
same group (i).
Consumption rate (Qij), i.e., the amount of (i) consumed by group (j), is calculated using
“foraging arena”- concept. Foraging arena theory assumes that trophic interactions are limited
by spatial foraging arenas and the concept divides prey populations (i) into vulnerable and
invulnerable components (Ahrens et al. 2012).
Consumption rate can be explained as follows (Eq. 3) (Christensen & Walters 2004;
Christensen et al. 2008)
Qij = (aij ∙ vij ∙ Bi ∙ Pj ∙ Ti ∙ Tj ∙ Sij ∙ Mij / Dj) / (vij + v´ij ∙ Tij ∙ Mij + aij ∙ Mij ∙ Pj ∙ Sij ∙ Tj / Dj) (Eq.3)
where aij is the effective search rate for predator (i) feeding on prey (j). Base vulnerability vij
express the rate in which the prey move between vulnerable and invulnerable. Bi represents the
biomass of predator, and Pj the abundance of prey. Ti and Tj respectively represent relative
feeding time for predator and prey. Sij is defined by the user to seasonal or long-term forcing
effects (in this study this was set to long-term forcing). Mij is environmental time series, i.e.,
mediation forcing effects and Dj defines the effect of handling time as a limit to consumption
rate.
13Moreover, the equation for consumption rate (Eq. 3) includes a vital feature of Ecosim, the
ability to explore biomass control by different functional groups, i.e., predator control (top-
down) or prey control (bottom-up). This is modeled by vulnerabilities (v) i) low v (close to 1)
indicates that an increase of predation biomass does not cause any noticeable changes in
predation mortality the predator may cause on the given prey, ii) high v (>2), e.g., doubling of
predation biomass, will cause a doubling in predation mortality for the given prey.
Default values of v’s were set to vij = 2 and represents a mixed flow control (Christensen et al.
2008; Christensen & Walters 2004; Tomczak et al. 2012).
Calibration and forcing data for Ecosim model of Gotland food web
Step 4. For the calibration procedure, which was the first step in constructing the Ecosim model
of Gotland food web, calibration data and forcing data from 1980 to 2000 were applied. Fish
biomass (calibration data) (Appendix: Table A6) and fishing mortality (forcing data) (Fig. 4)
for ad cod, small cod, ad. herring, juv. herring, ad. sprat and juv. sprat were calculated from
Stochastic Multi-Species (SMS) model output (Lewy & Vinther 2004)
Fig. 4. Fishing mortality (F) for the three main fish species from 1980 to 2009, applied as forcing variable. Note the
different scales.
14Calibration data was used also for groups: phytoplankton, zooplankton, and benthos groups
(Appendix: Table A6; Fig. 8; Fig. 9; Fig. 10). In order to fit the model to applied calibration
data, environmental forcing variables were used (Appendix: Table A7; Fig. 5). The forcing
variables were applied as relative change from the baseline year 1980 and consisted of: spring
temperature, August sea surface temperature (August SST), salinity (psu), oxygenated area
(km2), cod reproductive volume (cod RV) and primary production (PP) (Fig. 5). The cod RV is
defined as the volume of water with a salinity concentration above 11 psu and oxygen
concentration higher than 2 ml/l and it is in these conditions that cod eggs survive (Plikshs et
al. 1999).
Fig. 5. Environmental forcing variables in relative changes from the baseline year 1980. Note the different scales.
15Calibration of Gotland food web model
Step 5. After the appliance of calibration data, each trophic group, as well as the entire food
web, was provided with a statistical goodness of fit measure, SS-value (Sum of Squares). The
SS-value is a measure of deviation of the model estimates from the calibration data, i.e., the
higher the SS-value, the more deviated model estimates are from calibration data. To obtain a
calibration and later on a final Gotland food web model with a low SS-value, an automated
fitting procedure was used.
Fitting procedure of Gotland food web model
Step 6. The fitting procedure was done in steps, and after each step, the model was refitted. The
refitting involves searching for the best-fit vulnerability values for the most sensitive predator-
prey relationships to changes in vulnerability values. This was done on 15-relationships and
after every combination tested a new SS-value was calculated.
However, a conservative approach was taken concerning predator-prey relationships of
phytoplankton, microzooplankton and bacteria, thus they were given the default vulnerability
of 2. The approach was taken due to a limited amount of information about the trophic
relationships of these groups, and when running the model, low vulnerabilities resulted in very
intensive cascades of primary production to upper trophic levels. The steps of the fitting
procedure were as follows (see Appendix: Table A8 for SS-improvement after each applied
step):
(1) Cod RV from the Central Baltic Sea was used to force cod egg production and August
SST (sea surface temperature, 0-10 m) was used to force sprat egg production.
(2) Spring temperature (March-May, 0-50 m) forced the prey search rates of Acartia spp.
and Temora sp.
(3) Annual deep-water salinity (80-100 m) was forcing the prey search rate of P. acuspes.
(4) The last forcing step on search rate was forcing by oxygenated area on Mysids and
Saduria.
(5) Finally, primary production was applied as production rate on phytoplankton.
16Step 7. After the fitting procedure, the model with the lowest SS-value was chosen. This model
was then run from 2001 until 2009 using environmental (env.) forcing data (Appendix: Table
A7; Fig. 4 & Fig. 5) resulting in model biomass estimations from 2001 to 2009 and a final
Gotland food web model.
Calibration data was available for 2001 to 2009 (hence referred to as validation data) for groups:
cod (small and ad.), herring and sprat (juv. and ad.), mysids, macrozoobenthos, zooplankton
and phytoplankton, therefor was the 2001-2009 suitable as model validation period.
Future scenarios
Step 8. Future simulations of Gotland food web were applied from two different initial states,
(i) and (ii), and were projected until year 2080 in order to test how different initial states affects
applied management strategies in regards to ecosystem outcome. See Fig. 1 for a schematics of
climate-nutrient-cod fishing scenarios applied from initial state A and initial state B.
i) Initial state A represented the post-regime shift state, i.e., from 2009. The state was
characterized by a high abundance of Acartia spp. and Temora sp. and a lower
abundance of P.acuspes. The cod / sprat ratio was larger, with a high sprat
abundance. Compared to the average levels in initial state B, were the average
temperature (spring and August) higher and primary production higher, salinity and
cod RV levels were lower and there was an increase of hypoxic area. The adult cod
mortality was on an average level of 1.0, i.e., high cod fishing pressure.
ii) Initial state B represented the ecosystem structure present in 1980 to 1984, pre-
regime shift in the CBS. Characteristic of the ecosystem at that time was, high
abundance of P.acuspes compared to Acartia spp. and Temora sp. The state had a
higher abundance of cod then sprat and the average cod mortality was 0.8.
The average temperature (spring and August) were higher than the baseline year,
i.e., 1980. Deep-water salinity had a stable concentration through the period and an
average salinity level of 10 psu.
17Each state was used as a baseline for running future climate, nutrient and cod fishing scenarios
(See Fig. 1, for future climate-nutrient-cod fishing scenarios). Climate-nutrient-cod fishing
were all applied gradually over a 10-year period from the initial state; initial state A from 2009
and initial state B from 1984. Three future climate scenarios were used, two based on IPCC
emission scenarios and one scenario with no further climate change (Fig. 6). Each climate
variable was applied as relative change from the baseline year 1980. The baseline year was
calculated as a 10-year average, in order to reduce the risk of 1980 being an anomaly year.
i) No further climate change (No CC): No CC is used to get a climate-neutral baseline
of each state. Time series for all forcing variables were generated based on the
historically normal distribution, given the mean and standard deviation of each
environmental variable from 1980 to 2009 (following the approach in Gårdmark et
al. 2013).
ii) A1B: characterized as a balance between the use of fossil fuel technology and
alternative technology (Nakićenović & Swart 2000).
iii) A2: slow technology change, a high population growth projected resulting in higher
greenhouse gas emissions than A1B (Nakićenović & Swart 2000).
Fig. 6. Oceanographic forcing under climate scenarios A1B. A2 and No CC. Presented as the relative change from the baseline year 1980.
18Two nutrient load scenarios Baltic Sea Action Plan (BSAP) and business as usual (BAU) were
combined with climate scenarios A1B and A2 (Fig. 7). The Rossby Centre Ocean (RCO)
circulation model was combined with Swedish Costal and Ocean Biochemical (SCOBI) model
to project oceanographic and biochemical variables (Meier et al. 2012) for the combined
Fig. 7. Biochemical forcing in climate scenario A1B, A2 and No CC. Presented as the relative changes from the baseline year 1980.
climate-nutrient load scenarios. The nutrient scenarios were based on Gustafsson et al. (2011)
i) BSAP: Baltic Sea Action Plan - reduction of nutrient load combined with a possible
50 % atmospheric nitrogen decrease.
ii) BAU: Business As Usual - assumes an agricultural growth in all Baltic Sea countries
thus increasing riverine nutrients, and the atmospheric nitrogen levels stay at current
level.
19Finally, two cod fishing mortality conditions were combined with climate-nutrient scenarios;
A1B-BAU-BSAP and A2-BAU-BSAP, and with the neutral climate-nutrient scenario NoCC.
high fishing pressure (F1.1) and CRP – Cod Recovery Plan (F0.3). The level of fishing mortality
in F1.1 scenario was from the work of Niiranen et al. (2013a), where fishing mortality was based
on the average mortality years 2002 to 2006.
Analysis of the model results – principle component analysis
Step 9 and 10. Four types of time series were analyzed, biomasses (biotic (bio)) and
environmental forcing variables (abiotic (abio)):
1) Calibration data from 1980 to 2000 (see step 4)
2) Calibration data (1980-2000) plus validation data (2001-2009) (see step 4 & 7)
3) Model-estimated biomasses (1980 to 2009) from Gotland food web model and
individual time series of environmental forcing variables (see Fig. 5).
4) Model-estimated biomasses from future scenarios and environmental forcing variables
were analyzed after the transition period, i.e., initial state A from 2020 to 2080, and
initial state B from 1995 to 2080.
To study the pattern of biomasses, principle component analysis (PCA) was chosen and each
type of time series were analyzed accordingly. The first step (step 9) was to log-transform biotic
data to make the pattern of change over time clear by making data less skewed thus facilitating
the PCA.
Step 10 was the PCA analysis and it was carried out using RStudio (2012) (version 3.0.2) and
the package vegan: Community Ecology Package (version 2.0-10) (Oksanen et al. 2013).
Groups with no calibration data (bacteria, microzooplankton, cod larvae, juv. cod and detritus)
were excluded from the PCA analysis of time series (1), (2) and (3). The PC1 score is used as
a proxy of ecosystem state for the biotic (PC1_bio) and then used for further regime shift
analysis (as in Möllmann et al. 2009).
20Analysis of the model results – STARS
Step 11. To identify the timing and magnitude of possible regime shifts, the Sequential method
for detection of regime shifts (STARS) (Rodionov 2005) was applied to PC1_bio and individual
time series of forcing variables.
The STARS method is based on sequential t-test, which explores a time series of data points
(e.g., let X1, X2, X3…Xn be a time series of data points). The method tests incoming data points
in the time series to determine whether the new value has a statistically significant deviation
from the mean value of the current regime, i.e., shift in mean (Rodionov 2005).
For analysis of PC1_bio and environmental forcing from time series (1), (2) and (3), the
significance level (p) was set to 0.05, the cut-off length 10 years and Huber’s weight parameter
was set to 2. The p-level of 0.05 was chosen due to the stochastic nature of environmental
monitoring and measuring data. The cut-off length of 10 years was choice due to the short
length of the time series analyzed. The Huber’s weight parameter defines the range in standard
deviation of when a measurement is considered an outlier to the mean of the regime, and the
value 2 means that all measurements that are less than 2 standard deviations have an equal
weight, thus not an outlier (Rodionov 2006).
For analysis of future time series and projections, i.e., time series type (4), the p-level was set
to 0.01, because I wanted to capture the regime shift with the highest deviation, i.e., the stronger
regime shifts. The cut-off length was set to 30 years, due to the longer time series and Huber’s
weight parameter was kept at as 2.
Regime Shift Index (RSI) is a measure of the total sum of normalized deviations from the
assumed mean values of the new regime, from which the difference from the mean level for the
current regime is statistically significant, and can indicate the magnitude of change between
regimes (Rodionov & Overland 2005).
21RESULTS
Model performance
Model fit and validation
Gotland food web model (represented by the black line in Fig. 8) was able to estimate
calibration data of fish biomass relatively well (represented by the blue dots in Fig. 8).
The estimated biomasses from Gotland food web model had a difficulty to simulate peaks in
calibration data, for example, juv. sprat, juv. herring and both cod groups. Adult sprat biomass
was underestimated while adult herring biomass was overestimated, despite the under-and
overestimation the results were able to capture the trends in calibration data. Even though the
estimated biomasses were unable to capture the peaks in cod biomass, the remaining time series
of calibration data were estimated well.
Fig. 8. Gotland food web model. Estimated absolute fish biomass (the black line), calibration data (the blue dots)
and validation data (the pink dots). Note the different scales.
22Biomass estimations of the benthos groups Saduria, Monoporeia/Pontoporeia, other zoobenthos
and mysids had a difficulty to simulate calibration data, possibly due to the scarce amount of
calibration data and gaps in the time series (Fig. 9).
Fig. 9. Gotland food web model. Estimated absolute zoobenthos biomass (the black line), calibration data (the blue dots)
and validation data (the pink dots). Note the different scales.
23Acartia spp. and P. acuspes were well simulated by the biomass estimations, while simulations
of Temora sp. was slightly overestimated, even if the overall trend in calibration data was
captured. Cladocerans have the largest deviation from the calibration data (Fig. 10). The
estimated biomass of phytoplankton was overestimated, though followed the overall trend in
calibration data.
Fig. 10. Gotland food web model. Estimated absolute zooplankton and phytoplankton biomass (the black line),
calibration data (the blue dots) and validation data (the pink dots). Note the different scales.
24The model includes four groups with no calibration time series (Fig. 11). Bacteria biomass has
an overall pattern of fluctuations with a strong decrease in 1988 and a peak in 1995. Biomass
of microzooplankton has a smooth trend though biomass peaks in 1994-1995 and 2003. Cod
larvae and juvenile cod have a trend of decrease and it is alike the biomass trend of small and
adult cod.
Fig. 11. Gotland food web model. Estimated absolute biomass of groups with no calibration data (the black line).
Note the different scales.
The Gotland food web model was calibrated for the period 1980 to 2000 (see step 4 in “Material
and Methods”), and not using calibration data for 2001 to 2009 enabled model validation of
estimate biomasses. Validation data (represented by the pink dots in Fig. 8, Fig. 9 and Fig. 10)
were available for groups: phytoplankton, zooplankton, other zoobenthos and for six fish groups
(adult cod, herring and sprat and small cod, juvenile herring and sprat). The validation data for
small and adult cod were well estimated by the model, and adult sprat and adult herring had an
overall trend of under-respectively overestimation of biomass (Fig. 8). The group ‘other
zoobenthos’ had three years of validation data that the biomass estimates had a difficulty to
estimate (Fig. 9). Estimations of Acartia spp., Temora sp. and P. acuspes followed the trend in
validation data, and Cladocerans had the same trend in estimated biomass throughout the time
series (Fig. 10) and validation data was not captured.
25Phytoplankton biomass was overestimated, though followed the trend in validation data. The
validation showed that the Gotland food web model was capable to estimate overall biomass
trends in validation data by the use of environmental forcing variables.
Estimated biomass trends for Gotland food web from 1980 to 2009
Gotland food web model estimated changes in annual biomass for all trophic groups, ranging
from almost no change, e.g., mysids to a 2.7-fold increase of Temora sp. biomass from 1980 to
the end of 2009 (see Fig. 8, Fig. 9 and Fig. 10). Phytoplankton biomass slowly decreased (from
9.3 to 8.4 t km-2), while Acartia spp. increased 1.4-fold in biomass (Fig. 5). P. acuspes
displayed a 0.45-fold decrease of biomass with two sharp declines, the first one in 2002-2003,
followed by one in 2007-2008.
Mysids and Saduria displayed different changes, mysids had a slight decrease (from 1.21 to
1.04 t km-2) in biomass despite a fluctuating biomass over time, while Saduria had a smooth
development with an increase (1.2-fold increase) of biomass (Fig. 9).
Juvenile and adult sprat showed a similar change over time, both groups were estimated to
increase in biomass (2.6-fold respectively 2.1-fold) and the same trend of increasing biomass
was estimated in juvenile and adult herring (1.9-fold respectively 1.8 increase). All stanzas of
cod decreased drastically, where small cod decreased by 98.5 % and adult cod 96.2 % (Fig. 8).
26Detected regime shifts in Gotland food web model
Regime shift detection was carried out on three time series for the past dynamics (see “material
and Methods” section: Analysis of model results – STARS).
The PC1_bio -score (proxy of ecosystem structure) from the first time series, i.e., only
calibration data from 1980 to 2000 had one regime shift detected in 1991 (RSI: 0.74). The
second time series, i.e., calibration and validation data 1980 to 2009, had on regime shift in
ecosystem structure in 2001 (RSI: 2.3). The third and last analyzed time series was a time series
of estimated biomasses from Gotland food web model, two regime shifts were detected, the
first in 1991 (RSI: 0.77) and the second in 2002 (RSI: 0.1).
The detected regime shifts in the Gotland food web (year 1991 and 2002) were precede, co-
occurred and followed by shifts in env. forcing variables (PC1_abio). Preceding the first regime
shift was a shift in salinity in 1988 (RSI: 0.7), followed by shifts in primary production and
oxygenated area, both occurred in 1991 (RSI: 0.65 respectively 0.14). In 1996 was one shift in
salinity (RSI: 1.1) and oxygenated area had a regime shift in 1999 (RSI: 0.55), followed by a
regime shift in primary production year 2000 (RSI: 0.03). Salinity had a regime shift in 2006
(RSI: 0.08), followed by a shift in spring temperature in 2007 (RSI: 0.36) and lastly primary
production had a regime shift in 2008 (RSI: 0.35). In Fig. 12, are all detected regime shifts
displayed on a timeline. August temperature and cod reproductive volume did not have any
detected regime shifts between 1980 to 2009.
Fig. 12. Timeline of detected regime shifts in Gotland food web model. The height of the bars represents RSI value, i.e., the magnitude of
change. No regime shifts were detected for August SST nor Cod RV, thus they are not represented in the timeline.
27Future scenario projections
Trends in future biomass from two different initial states
Environmental conditions were projected to change in the future (Fig. 6) and in conjunction
with those changes were changes in biomass of trophic groups (Fig. 13). Different climate,
nutrient loading and cod fishing scenarios resulted in a variety of future biomass trajectories
from each initial state of Gotland basin ecosystem. Here follows a description of general trends
in future biomass outcome and some differences between and within initial states.
Fig. 13. Average biomass trends for selected groups in different future scenarios (BAU, business as usual; BSAP, Baltic Sea Action Plan;
for fishing:
F0.3 = cod recovery plan, F1.1 = high cod fishing pressure) from initial state A and initial state B.
Arrows indicate an increase or decrease in biomass from the average past conditions in biomass 1980-2009 (REF-period) for the given
group. Initial state A to the left, period 2020-2080 compared to REF-period and initial state B to the left, period 1995-2080 compared to
REF-period.
Black arrow = ± ≥ 200 %, blue arrow = ± 150-199 %, red arrow = ± 100-149 %, yellow arrow = ± 10-99 %, green arrow = ± 10-49 % and
‘−‘ indicates no change in biomass (max ± 10 %).
The overall trend from both initial states in high nutrient scenario ‘business as usual’ (BAU)
was an increase of key functional groups, i.e., ad. cod, ad. herring, ad. sprat, ‘other zoobenthos’,
P.acuspes, Acartia spp. and phytoplankton, in all three climate scenarios. There were no
projected deviation from the general trend of increase in BAU-scenario, however the level of
change (i.e., the increase or decrease form the average past conditions, 1980 to 2009, i.e., REF-
period) differs between initial states and climate scenarios. For example, adult sprat has a very
high increase (i.e., + 150-199 %) of biomass from initial state A (high sprat, low cod ecosystem)
but from initial state B (high cod, low sprat ecosystem) the adult sprat had a lower increase (i.e.,
+ 100-149 %). Another interesting observation in the level of change between initial states,
there are more groups with a higher level of increase (i.e., black, blue and red arrows, see Fig.
13, for exact levels of change) from initial state A than initial state B.
28Lastly, there were no indications of cod fishing pressure affecting the level of change in a high
nutrient scenario (Fig. 13). However, in the biomass trajectory of adult cod (Appendix: Fig.
A16; Fig. A17), the low cod fishing pressure (F0.3) resulted in a higher level of biomass than
with a high cod fishing pressure (F1.1), thereby indicating an effect of cod fishing pressure on
ad. cod biomass development over time.
The general trend in a reduced nutrient loading scenario, i.e., Baltic Sea Action Plan
(BSAP) was an overall decrease of biomass from each initial state, though with some
exceptions. For example, was P. acuspes projected to increase in biomass from initial state A
and initial state B, in all climate scenarios. However, as pointed out with adult cod in BAU, the
biomass trajectory displayed a different outcome of P. acuspes, from initial state A were it was
projected to almost collapse in BSAP-scenario around the mid-2050s, in both climate scenario
A1B and A2 (Appendix: Fig. A16; Fig. A17).
Adult cod was also projected to increase in biomass from the REF-period (i.e., the average past
conditions for adult cod, 1980 to 2009), especially with a low fishing pressure (F0.3). A high
fishing pressure (F1.1) resulted in an almost cod stock collapse from the mid-2060s, this was
displayed from initial state A and initial state B, climate scenarios A1B and A2 (Appendix: Fig.
A16; Fig. A17).
Lastly, the overall level of change in biomass was lower (i.e., no change, green and yellow
arrows, see Fig. 13, for exact levels of change) with a reduced nutrient loading than with an
increase of nutrients (BAU), the exception being adult cod, which was projected to have a high
increase (i.e., an increase of 150-199 % and ≥ 200 %) with F0.3 from both initial states and
climate A1B and A2.
No further climate change was a scenario with no added or reduced nutrient loading. The
general trend was an increase of biomass from both initial state A and B, though with a higher
level of change than in BAU (i.e., green and yellow arrow, see Fig. 13 for exact levels of
change). No further climate change had one exception, adult sprat, was projected to have a low
decrease (i.e., decrease of 10-49 %) of average biomass from REF-period (i.e., average past
conditions, 1980-2009) (Fig. 13).
29Detected regime shifts from two different initial states
Future time series from each scenario combination were analyzed for eventual regime shifts in
ecosystem structure (PC1_bio) and in environmental forcing variables (PC1_abio) (see
“Material and Methods” step 9-11). The STARS-analysis detected a different amount of regime
shifts depending on nutrient loading, increase of nutrients as in business as usual (BAU) or with
a reduction of nutrients as in the Baltic Sea Action Plan (BSAP).
Projected future regime shifts under high nutrient loading
Initial state A (today’s sprat dominated system, Fig. 14) and initial state B (past cod dominated
system, Fig. 15) had two respectively three regime shifts detected in ecosystem structure
(PC1_bio) in climate A1B and A2, business as usual- high nutrient scenario. The timing of the
regime shifts were the same within each climate scenario, and very similar in magnitude despite
different cod fishing pressures, i.e., low (F0.3) and high (F1.1). The characteristics of detected
regime shifts from initial state A (Fig. 14) in ecosystem structure were their co-occurrences or
directly followed by a regime shift in primary production. For example, in climate scenario A2,
a regime shift in ecosystem structure was detected in 2040, which co-occurred with a regime
shift in salinity and was directly followed by a regime shift in primary production (in year 2041)
and in 2044 a shift in oxygenated area.
Fig. 14. Detected regime shifts from future scenarios (BAU, business as usual; BSAP, Baltic Sea Action Plan; for fishing:
F0.3 = cod recovery plan, F1.1 = high cod fishing pressure) from initial state A.
The height of the bars indicates the level of RSI, i.e., magnitude of change and bars with negative RSI are directed downwards.
a)Spring temperature b) August SST c) Salinity d) Primary production e) Oxygenated area f) Cod RV g) Ecosystem structure
(PC1_bio)
30You can also read

















































