Temporary streams in temperate zones: recognizing, monitoring and restoring transitional aquatic-terrestrial ecosystems - Loughborough ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Opinion
Temporary streams in temperate
zones: recognizing, monitoring
and restoring transitional aquatic-
terrestrial ecosystems
Rachel Stubbington,1* Judy England,2 Paul J. Wood3 and Catherine E.M. Sefton4
Temporary streams are defined by periodic flow cessation, and may experience
partial or complete loss of surface water. The ecology and hydrology of these
transitional aquatic-terrestrial ecosystems have received unprecedented attention
in recent years. Research has focussed on the arid, semi-arid, and Mediterranean
regions in which temporary systems are the dominant stream type, and those in
cooler, wetter temperate regions with an oceanic climate influence are also
receiving increasing attention. These oceanic systems take diverse forms, includ-
ing meandering alluvial plain rivers, ‘winterbourne’ chalk streams, and peatland
gullies. Temporary streams provide ecosystem services and support a diverse
biota that includes rare and endemic specialists. We examine this biota and illus-
trate that temporary stream diversity can be higher than in comparable perennial
systems, in particular when differences among sites and times are considered;
these diversity patterns can be related to transitions between lotic, lentic, and ter-
restrial instream conditions. Human impacts on temperate-zone temporary
streams are ubiquitous, and result from water-resource and land-use-related
stressors, which interact in a changing climate to alter natural flow regimes.
These impacts may remain uncharacterized due to inadequate protection of
small temporary streams by current legislation, and hydrological and biological
monitoring programs therefore require expansion to better represent temporary
systems. Novel, temporary-stream-specific biomonitors and multi-metric indices
require development, to integrate characterization of ecological quality during
lotic, lentic, and terrestrial phases. In addition, projects to restore flow regimes,
habitats, and communities may be required to improve the ecological quality of
temporary streams. © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc.
How to cite this article:
WIREs Water 2017, 4:e1223. doi: 10.1002/wat2.1223
INTRODUCTION
*Correspondence to: rachel.stubbington@ntu.ac.uk
1
School of Science and Technology, Nottingham Trent University,
T emporary streams (TS) are lotic ecosystems in
which water sometimes stops flowing.1 Defining
TS based on flow intermittence (i.e., the loss of lotic
Nottingham, UK
2
Research, Analysis and Evaluation, Environment Agency,
surface water movement) disguises a critical distinc-
Wallingford, UK tion: the presence or absence of surface water,2 with
3
Centre for Hydrological Ecosystem and Science, Department of persistent pools being isolated, extensive, or continu-
Geography, Loughborough University, Loughborough, UK ous in some systems, whereas others lose all surface
4
Centre for Ecology and Hydrology, Wallingford, UK water. A second key distinction between intermit-
Conflict of interest: The authors have declared no conflicts of inter- tence regimes is predictability: some systems experi-
est for this article. ence predictable flow cessation or drying during
Volume 4, July/August 2017 1 of 17
© 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc.
This is an open access article under the terms of the Creative Commons Attribution-NonCommercial License, which permits use, distribution and
reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.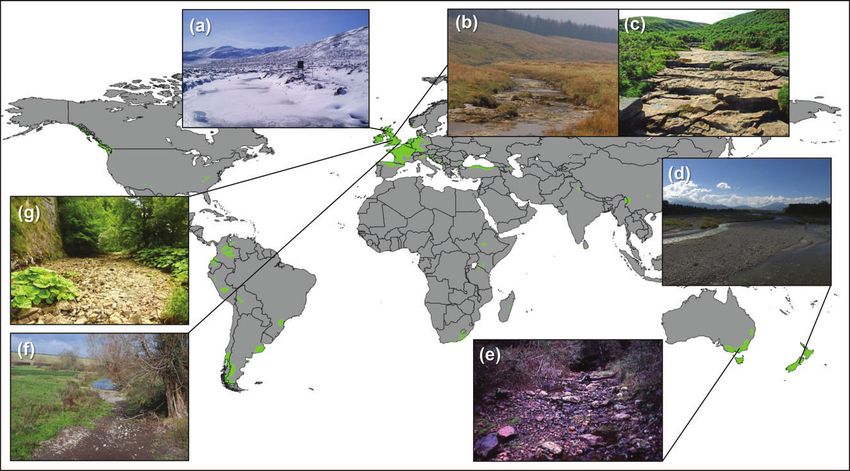
Opinion wires.wiley.com/water
summer or dry seasons, whereas unpredictable wet- temperatures and year-round precipitation, and occur
dry cycles characterize other streams. Although TS primarily in north-western Europe and south-eastern
have been conceptualized as coupled ecosystems Australasia (Figure 1).
that transition between aquatic and terrestrial We highlight the diverse range of TS that occur
conditions,3,4 current definitions remain freshwater- in oceanic regions, the high biodiversity that these
focussed. From a terrestrial perspective, TS can be streams support, and the ecosystem functions that oce-
viewed as linear features that intersect otherwise con- anic TS provide. We outline threats to the ecological
tinuous habitat patches and experience periodic integrity of TS, and identify opportunities to combat
inundation. Equally, their substrates can be concep- current impacts within a context of international legis-
tualized as permanent habitats that repeatedly fluctu- lation. We identify research priorities, including the
ate between aquatic and terrestrial conditions.5 work needed to facilitate incorporation of TS into
TS can be the dominant lotic ecosystem type in hydrological and biological monitoring programs.
arid, semi-arid, and Mediterranean-climate regions, Although variable terminology has been used to
where their hydrology and ecology are relatively well- describe TS, for simplicity, we use: TS to refer to eco-
studied. TS are also common in temperate regions with systems that lose flowing surface water and often expe-
cooler, wetter climates, their occurrence reflecting inter- rience complete drying, and intermittent to describe
actions between climatic drivers (e.g., precipitation and such systems; we recognize that these simple definitions
temperature), physical catchment characteristics reflect our primarily freshwater-related perspective.
(e.g., bedrock and overlying sediments), and human
influences (e.g., water abstraction, effluent discharge,
and land use). We complement recent global TS research A DIVERSE RANGE OF TEMPORARY
by focusing on systems in temperate regions with an oce- STREAMS OCCUR IN OCEANIC
anic influence on the climate (Cfb under the Köppen cli- REGIONS
mate classification; hereafter, ‘oceanic’). Oceanic
Although dry streambeds may be perceived as sym-
climates are characterized by cool, moderate
bolizing human impacts including water abstraction
F I G U R E 1 | Examples of temporary streams in oceanic (Cfb) environments: (a) a snow-covered mountain stream, Scotland, UK; (b) a ponded
peatland tributary of Green Field Beck, England, UK; (c) the bedrock-dominated channel of Deepdale Gill, England, UK; (d) the alluvial plain River
Orari, New Zealand; (e) the forested Lerderderg River, Australia; (f ) the winterbourne headwaters of the chalk River Till, England, UK; and (g) the
karst River Manifold, England, UK. Photo credits: A. Youngson (a); L. Brown (b); J. Clift (c); F. Burdon (d); A. Boulton (e); A. House (f );
R. Stubbington (g).
2 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017
WIREs Water Temporary streams in temperate zones
and climate-change-related drought,6 TS are natural dry-channel specialists in tropical, sub-tropical, and
ecosystems in oceanic climates (Figure 1). For exam- alpine TS, with inundation-tolerant life stages pro-
ple, flow cessation and drying is natural in channels posed for some taxa.5,22,23 Such terrestrial specialists
underlain by fissured karst and porous chalk bed- are unknown in oceanic dry channels, which may
rock, and in alluvial plain rivers flowing over perme- reflect the reduced extent of drying or the limited
able deposits (Figure 1). However, TS diversity research conducted.24 Beetles associated with
remains poorly characterized across oceanic regions, exposed riverine sediments25 in perennial rivers expe-
and in particular, headwater streams that experience rience repeated inundation, and specialists with com-
long dry periods have received limited attention,7 parable tolerance of hydrological fluctuations may
despite comprising a considerable proportion of river colonize dry streambeds.
networks and making a disproportionate contribu-
tion to regional biodiversity.8
The spatial arrangement of perennial and Local-scale Lotic Biodiversity Typically
intermittent reaches varies between systems. Inter-
mittence may increase gradually with progression
Declines as Intermittence Increases
upstream due to seasonal water table fluctuations, During flowing phases, site-specific aquatic commu-
as in ‘winterbourne’ chalk streams9 and systems dis- nity diversity (i.e., α-diversity, defined in Box 1 and
secting upland landscapes.7 Elsewhere, a mosaic of explored in Box 2) is typically lower at intermittent
segments with varying intermittence reflects local sites compared to equivalent perennial sites, as
changes in geomorphological controls.10 River regu- demonstrated for invertebrates across climate zones12
lation and water-resource management can pro- and within oceanic regions.19,26 Sites with greater
foundly alter flow intermittence regimes. Over- intermittence are typically inhabited by a subset of
abstraction can cause naturally perennial reaches to the generalists found at perennial and less intermit-
dry,9 water supply diversions and effluent discharges tent sites.12 However, where flowing phases are long-
can result in artificial perennialization,11 and lotic lasting and intermittent reaches are short in spatial
reaches upstream of impoundments may become extent, site-specific lotic diversity may increase over
lentic.
BOX 1
TEMPORARY STREAM COMMUNITIES TEMPORARY STREAM COMMUNITY
ARE BIODIVERSE DIVERSITY: DEFINING DIVERSITY
TS communities are often dominated by generalists12 The taxonomic diversity of an assemblage
(i.e., those also found elsewhere) including resilient describes its richness and evenness, i.e., the
taxa that colonize when appropriate habitats become number of taxa present and the relative contri-
available: lotic taxa when flow resumes, lentic taxa bution each makes to total abundance. Alpha
when pools form, and terrestrial taxa when sedi- (α) diversity considers the taxa present locally,
ments dry. In addition, where the evolutionary driver in an individual sampling unit (Figure 2(a)).
of intermittence is sufficiently strong12 and predicta- Beta (β) diversity describes heterogeneity
ble13 to favor specialization, TS specialists may among sites or times and comprises ‘variation’
enhance biodiversity. During flowing phases, some and ‘turnover’ components. Variation
specialist caddisfly larvae inhabit intermittent springs β-diversity refers to differences in assemblages
in karst networks,14 and winterbourne chalk streams among a set of sampling units (Figure 2(b))
support characteristic plant communities9,15 and spe- whereas turnover β-diversity describes differ-
cialist stonefly16 and mayfly17 nymphs, the latter ences among sampling units positioned along a
including nationally rare species in the UK.18 spatial (S1, S2) or temporal (T1, T2) environmen-
tal gradient (Figure 2(c)). Together, α and β
Other aquatic flora and fauna are headwater
components determine gamma (γ), or total
specialists,8,9,15 their isolation increasing endemicity,
regional-scale diversity. Diversity measures can
as reported for a blackfly restricted to few English
also be applied to nontaxonomic categories,
winterbourne chalk streams.19 During pool phases,
e.g., functional diversity considers the range of
specialists of temporary lentic habitats may colonize; traits (characteristics influencing an organism’s
for example, caddisfly larvae largely restricted to response to the environment) possessed by taxa
floodplain ponds20 may also inhabit TS pools.21 In in an assemblage.
addition, terrestrial invertebrate assemblages include
Volume 4, July/August 2017 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. 3 of 17Opinion wires.wiley.com/water
F I G U R E 2 | Concepts of diversity illustrated using a theoretical stream network: (a) α-diversity; (b) variation β-diversity; and (c) turnover
β-diversity along simplified two-site (S1, S2) and two-time (T1, T2) ‘gradients’. Small filled symbols indicate different ‘types,’ e.g., taxa or traits,
their size representing their relative abundance; larger circles and semi-circles indicate sampling units; blue lines represent a plan view of the
stream network. In each theoretical case, the left-hand side of the network (as viewed) has higher diversity than the right-hand side; the patterns
observed in temporary streams are described in Box 2.
BOX 2
degrees of intermittence (stβ1-2, Figure 3(a) and
(b)). Equally, variation β-diversity of lotic com-
TEMPORARY STREAM COMMUNITY munities may be higher among intermittent
DIVERSITY: ENVIRONMENTAL sites sampled during flowing phases (vβ1-2,
HETEROGENEITY PROMOTES HIGH Figure 3(a)) compared to perennial sites (vβ1,2,
FLOWING-PHASE BIODIVERSITY Figure 3(b)), due to spatial variation in environ-
mental conditions.32 Temporal turnover, spatial
Figure 3 compares typical patterns of α-, β-, and
turnover, and variation components of
γ-diversity (defined in Box 1) for communities in
β-diversity combine to increase γ-diversity (γ,
temporary and perennial stream networks. A-
Figure 3(a) and (b)) in temporary compared to
diversity is typically lower in temporary com-
perennial stream networks.
pared to perennial streams (α1, Figure 3(a) and
(b)), reflecting community nestedness: the taxa
at sites with greater intermittence are subsets
of those at perennial and less intermittent time and become comparable at intermittent and per-
sites.12 Over time, lotic diversity may become ennial sites27; diversity may even be higher at
comparable at intermittent and perennial sites intermittent sites due to greater local habitat
(α2, Figure 3(a) and (b)) as flowing phases heterogeneity.28
increase in duration, and may even become The typical pattern of local reductions in diver-
higher at intermittent sites (α3, Figure 3 sity with increasing intermittence also characterizes
(a) and (b)) characterized by local-scale habitat TS lentic communities, although in contrast to flow-
heterogeneity. ing phases, lentic specialists often replace generalist
Typical seasonal progression from flowing, taxa as wet-phase duration decreases.20,29 Finally,
to pool, to dry conditions represents a temporal the dry-phase diversity of terrestrial communities has
gradient in environmental conditions, and in been little researched in oceanic TS; a rare study by
response, turnover β-diversity is enhanced by Corti and Datry24 found that invertebrate assem-
shifts between lotic (ttβ1), lentic (ttβ2), and ter-
blages in a dry alluvial plain channel were subsets of
restrial biota (ttβ3, Figure 3(a)), as explored in
those in adjacent riparian zones. This pattern
Box 3; equivalent habitat changes and related
matches that observed for lotic invertebrates,12 but
shifts in community composition are modest in
perennial streams (ttβ1-3, Figure 3(b)). Turnover
contrasts with Steward et al.’s finding that 20% of
β-diversity along a spatial, longitudinal gradient terrestrial invertebrate taxa were unique to dry chan-
may also be higher in TS, due to sequential nels in alpine, tropical, and sub-tropical climates.22
replacement of taxa adapted to different Further research is required to examine lentic and
dry-phase diversity in TS channels across climate
4 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017WIREs Water Temporary streams in temperate zones
(a) Temporary (b) Perennial highlighting downstream perennial reaches and other
stream network stream network aquatic and terrestrial refuges as additional colonist
sources.12 Following pool formation, persisting lotic
Diversity at multiple sites during
biota are joined by lentic colonists, the latter reflect-
α1 stβ 2 α1 stβ 2
ing interactions between refuge availability and
stβ 1 stβ 1
taxon-specific dispersal abilities31; implications for
flowing phases
pool diversity are described in Box 3. Following
vβ 1 vβ 2 α2 vβ 1 vβ 2
α2
BOX 3
α3 α3
TEMPORARY STREAM COMMUNITY
ttβ 1 ttβ 1 DIVERSITY: TEMPORAL VARIABILITY IN
Diversity at FLOWING LOTIC, LENTIC, AND TERRESTRIAL
one site COMMUNITY RICHNESS
FLOWING
during an
annual cycle
ttβ 3 ttβ 3 ttβ 2 Figure 4 illustrates how sequential use may
ttβ 2
allow lotic, lentic, and terrestrial taxa to share
Total
an instream space.
diversity γ γ
When flow resumes, lotic taxa richness
FI GU RE 3 | Typical patterns of alpha (α), beta (β), and gamma increases rapidly as populations from refuges
(γ) diversity of communities in (a) a temporary stream network and within the intermittent reach proliferate35 and
(b) a perennial stream network, at multiple sites during flowing colonists arrive from elsewhere36 (Figure 4(a)).
phases (blue lines), and at one site during an annual cycle (black Generalists (which also inhabit perennial streams)
circles). The size of blue, filled symbols allows comparison of panes typically dominate this community, particularly as
(a) and (b) and is proportional to diversity, i.e., larger symbols indicate flowing phase duration increases.37 A few TS spe-
higher diversity at temporary or perennial sites; symbol sizes should cialists are also present, and lentic generalists
not be compared within a pane. Shapes indicate differences in may inhabit suitable microhabitats. If inundation-
community composition. Abbreviations: stβ, ttβ and vβ, spatial tolerant terrestrial taxa exist23 and remain within
turnover, temporal turnover, and variation β-diversity, respectively. the channel, their wet-phase persistence is proba-
Superscript letters allow comparison of (a) and (b). Definitions of bly short-lived.
diversity measures are provided in Box 1 and patterns are described in When flow ceases, extensive and connected,
Box 2. or sparse and isolated pools may form. Richness
may initially increase in persistent pools as len-
zones, including characterization of temporal changes tic colonists join lotic refugees, but later
and the extent to which increasing local-scale terres- decreases due to poor habitat suitability, declin-
trial biodiversity offsets declining aquatic diversity as ing water quality, and intense biotic interac-
intermittence increases. tions (Figure 4 (b)).5 TS pool specialists remain
undocumented in temperate regions but may
comprise a comparable biota to other tempo-
Environmental Controls on Local rary lentic waters20; limited global evidence
Biodiversity Extend Beyond Habitat suggests such specialists as minor contributors
Boundaries to pool communities.38
During dry phases, generalist terrestrial taxa
Reach-scale diversity reflects not only timing in a
arrive from the riparian zone and catchment,
hydrological cycle and local habitat characteristics,
increasing in richness as the dry phase proceeds
but also environmental influences and metacommu-
(Figure 4(c))39; dry-channel specialists have not
nity dynamics which extend beyond a local site and been identified in any temperate TS.5 Aquatic
interact to influence colonization processes.30,31 Fol- richness declines as desiccation-sensitive taxa
lowing flow resumptions, the occurrence of perennial are lost, but the ‘seedbank’ of desiccation-
upstream reaches is a notable influence on lotic colo- tolerant aquatic life-stages that persists within
nization rates, with drifting organisms promoting drying sediments40 maintains moderate rich-
rapid community recovery.10 However, as flowing ness.4,41 Seedbank richness declines with sedi-
phase duration increases, differences in peak lotic ment moisture, making oceanic assemblages
diversity among intermittent sites with and without relatively taxa rich.40,41
perennial upstream reaches may disappear,
Volume 4, July/August 2017 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. 5 of 17Opinion wires.wiley.com/water
concept that we suggest be extended to encompass
terrestrial taxa (Figure 4, Box 3).
As well as temporal changes, transitions in
environmental conditions along spatial, longitudinal
(a) gradients may enhance biodiversity (i.e., spatial
(b) turnover β-diversity, explored in Box 2) in TS com-
pared to equivalent perennial stream lengths, with
sequential taxon replacements reflecting adaptation
FLOWING to different degrees of intermittence. For example,
the macrophyte communities of chalk stream head-
waters are characterized by longitudinal zonation,
with specific assemblages associated with perennial
reaches and those with typical annual dry periods of
2–4, 4–6, and >6 months42 (Figure 5); site-specific
community composition also differs between wet
and dry years. Similarly, spatial environmental het-
erogeneity among multiple sites32 may mean that
flowing-phase variation in lotic community composi-
Key (c) tion (i.e., variation β-diversity, explored in Box 2) is
Lotic taxa Generalist taxa ? Hypothesized pattern
higher among intermittent compared to perennial
Lentic taxa Specialist taxa sites, as observed in arid,32 subtropical,2 and Medi-
Terrestrial taxa Symbol size reflects relative taxa richness terranean regions.28 During dry phases, differences
in terrestrial invertebrate assemblage composition
F I G U R E 4 | Turnover of lotic, lentic, and terrestrial taxa during (a) increase among-reach diversity,24 aligning with
flowing, (b) pool, and (c) dry habitat phases in a temporary stream aquatic patterns.2,28,32 Further research is required
reach. Arrows indicate a typical annual cycle of environmental to compare regional patterns, and to examine biota
changes; reversals (e.g., transitions from pool to flowing conditions)
previously combined as “nonaquatic” in freshwater
and omissions (e.g., flowing-dry-flowing or flowing-pool-flowing
studies.15,42
transitions) may also occur. Taxonomic patterns are those observed
and hypothesized for invertebrate communities, with some plant
Endemic species including TS and headwater
community data suggesting comparable patterns. specialists also increase diversity among sites,8,19
with one study of seven Mediterranean-climate head-
water TS describing 13 new aquatic or semi-aquatic
drying, the rate and extent of colonization by terres- insect species.28 The range of taxa restricted to iso-
trial organisms may reflect the characteristics of mar- lated headwaters may remain underestimated, in par-
ginal habitats, such as riparian vegetation, bank-top ticular for small and taxonomically challenging
height, bank slope, and bank-face materials.25 organisms.43
Research is needed to examine the environmental Regional-scale aquatic diversity (i.e., γ-diver-
controls of dry-phase colonization trajectories, and sity, explored in Box 2) may be highest in systems
to identify factors promoting dry-phase community with greater intermittence due to spatial and tempo-
establishment. ral environmental heterogeneity.2 In addition, total
TS diversity estimates are increased when both
aquatic and terrestrial taxa are recognized, as
Spatial and Temporal Environmental recorded for invertebrate24 and plant commu-
Variability Increases Biodiversity in TS nities.42 Further collaborations between aquatic and
Transitions between flowing, pool, and dry phases terrestrial ecologists are needed to quantify the total
result in greater temporal variation in environmental biodiversity of biotic groups across full hydrological
conditions in TS compared to perennial streams. In cycles.5,24,44 In addition, whereas macroscopic com-
response, shifts between communities dominated by munities of oceanic TS including chalk streams and
lotic, lentic, and terrestrial taxa mean that intermit- alluvial plain rivers are sufficiently well-
tent sites can have higher biodiversity than perennial characterized to warrant description of general pat-
sites when an annual cycle is considered,33 as terns, the biodiversity of systems including upland
explored in Boxes 2 and 3. Bogan and Lytle headwater TS requires further research, as do under-
described this sequential use of one location by lentic represented biotic groups including meiofauna and
and lotic taxa as ‘taxonomic time-sharing’34, a diatoms.
6 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017WIREs Water Temporary streams in temperate zones
FI GU RE 5 | In chalk streams including the River Misbourne (England, UK), macrophyte communities are characterized by longitudinal zonation
during the summer months, and differ between sites with (a) perennial flow, (b) shorter (typically 2-4 month) and (c) longer (typically 4-8 month)
annual dry periods, as described by Westwood et al..33 Photo credits: J. England (a); N. Holmes (b-c).
TEMPORARY STREAMS HAVE Ecosystem services that benefit people may also
IMPORTANT ECOSYSTEM differ regionally, with cultural services reflecting the
ubiquity of TS (Table 1). Native Australian folklore
FUNCTIONS
refers to TS fauna, and recreational events are held in
TS, including dry channels, have recently been concep- dry channels.23 In contrast, dry channels are viewed as
tualized as biologically active, biodiverse, and impor- symptomatic of poor ecosystem health in oceanic sys-
tant in ecosystem functioning.23 However, some wider tems such as English chalk streams,6,15 with recrea-
roles, such as acting as navigation corridors for biotic tional uses such as fishing and associated perceptions
migrations across landscapes, may be restricted to of high value largely restricted to flowing phases.
Mediterranean45 and arid46 regions with extensive Linked to these contrasting public perceptions of
intermittent networks (Table 1). Other functions may ‘value’, TS provisioning services include food and
also vary depending on climate.47 For example, dry water in arid landscapes53 but are minimal in devel-
streambeds in arid landscapes may be moist microhabi- oped temperate regions. In contrast, the regulating
tats favored by small mammals22 whereas those in service of flood mitigation, common to perennial and
tropical regions may be hotter and drier than the adja- intermittent streams, may be greater in densely popu-
cent riparian zone and so avoided by fauna.46 In con- lated temperate zones.
trast, pools may provide drinking water for livestock As well as supporting high biodiversity includ-
and wild animals across climate zones, particularly ing rare and endemic taxa, TS provide refuges for spe-
during droughts.48 Some ecosystem functions may dif- cialists outcompeted by generalists in perennial
fer depending on dry phase duration. For example, streams. For example, the absence of a dominant
occasional flow in arid streams provides rare opportu- grazing snail has been linked to greater diversity of
nities for aquatic organism dispersal23; in contrast, other invertebrate grazers in TS.8,28 Equally, natives
occasional drying of oceanic TS may facilitate dispersal threatened by non-native invasive species may use TS
of terrestrial organisms including mammals49 and tur- as refuges. For example, a greater decline in non-
tles50 across landscapes usually fragmented by water.51 native than native fish richness with increasing inter-
Other functions may be broadly comparable across cli- mittence has been attributed to poor adaptation to
mates, including organic matter processing: dry phases habitat contraction in Mediterranean streams.57
allow organic matter to accumulate before release and TS biota perform important ecosystem func-
transformation following rewetting and flow tions. During wet phases, aquatic invertebrates sup-
resumptions.52 port food webs that include fish as top aquatic
Volume 4, July/August 2017 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. 7 of 17Opinion wires.wiley.com/water
TABLE 1 | Temporary stream (TS) ecosystem functions provided by (a) dry channels (b) pools and (c) flowing stream habitats. Evidence of each
function is provided at a global scale and compared to oceanic zones, with functions hypothesized where data is lacking.
(a) Dry channels (b) Pools (c) Flowing streams
Ecosystem Oceanic zone Oceanic zone Oceanic zone
function Global evidence function Global evidence function Global evidence function
Migration/ Used by small Lower importance None None Established (but Lower
navigation mammals, due to limited contested54) importance
corridors reptiles, and spatial and perennial river due to limited
for birds in temporal extent; function; likely for spatial and
vertebrates deserts46 and local movements TS temporal
Mediterranean may be extent; may be
TS45 facilitated used by bats.
Inhabited by Moist habitat used Lower importance Not known Not known Not known Not known
terrestrial by hares, mice, due to
vertebrates and shrews in availability of
Namib Desert46 moist riparian
habitats
Habitat for Dry channel Lower importance Not known Not known Invertebrate specialists Invertebrate
specialist invertebrates in due to shorter dominate some arid specialists
biota tropical and dry phases; TS55 occur in TS
alpine streams22 none identified24 and springs14
Refuge for Seedbank Enhanced function Established, major Refuge for many Refuges from Greater
lotic taxa established as in oceanic TS dry-phase taxa, particularly competitive taxa importance
survival due to high refuge, e.g., in if habitats are including non-native due to
mechanism sediment Mediterranean, similar to invasive taxa, and widespread
across climate moisture semi-arid and flowing streams, from predation by river
zones40 content4,40 arid streams55 e.g., in fish in regulation and
regulated / Mediterranean28,57 invasive
lowland and other species in
streams56 temperate TS58 perennial
streams
Dispersal Easier cross- Higher importance May facilitate local May facilitate local Dispersal of aquatic Greater dispersal
channel due to limited dispersal of dispersal of and riparian flora of aquatic and
dispersal by spatial and lentic taxa lentic taxa and fauna riparian flora
terrestrial temporal between pools between pools established across and fauna due
organisms in windows in climate zones to long wet
arid zones due which terrestrial phases
to longer dry organisms can
phases disperse51
Carbon CO2 efflux from Not calculated for Organic matter Organic matter Organic matter Greater
cycling and dry organic temperate retained and retained and processing processing in
organic matter zones; processed in processed in following flow oceanic zones:
matter quantified in a considered pools pools60 resumptions processing
processing Mediterranean lower than arid established across declines as
river and up- zones59 climate zones drying
scaled to other increases61
regions59
Recreation Events, e.g. a ‘dry Limited importance Waterholes in Reduced Established across Lower
river race’ held due to extent of Australia importance due regions importance;
in Australia; larger systems; provide interest to limited spatial e.g. adjacent routes more
provide shaded TS in karst along walking and temporal walking routes likely to follow
walking routes landscapes can routes extent perennial
e.g. in Spain act as caving rivers
entry points
(continued overleaf )
8 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017WIREs Water Temporary streams in temperate zones
TABLE 1 | Continued
(a) Dry channels (b) Pools (c) Flowing streams
Ecosystem Oceanic zone Oceanic zone Oceanic zone
function Global evidence function Global evidence function Global evidence function
Provision of Aestivating fish Not known Drinking water for Lower importance Drinking water for Groundwater
food and consumed in deer, humans, due to higher humans and and surface
water for Botswana; cattle and livestock in water livestock in arid water
people grazed in Egypt; arid and semi- availability in zones62 abstraction for
48,62
crops grown in arid zones oceanic zones public water
India; water supply and
found by agriculture
digging23 across
temperate
zones9
predators63 and extend into riparian and terrestrial dominated by urban and agricultural land uses, caus-
habitats: adults of insects with aquatic juveniles subsi- ing or exacerbating shifts to greater intermittence,
dize terrestrial food webs upon emergence.64 These including artificial drying events in perennial streams
pulsed inputs are pronounced in TS if insects emerge and increased drying in TS. For example, decreased
en masse before drying, and where high aquatic insect discharge in the intermittent River Selwyn in
abundance is linked to exclusion of predators by New Zealand has been linked to abstraction for irriga-
intermittence.65 As drying phases proceed, aquatic tion76; the naturally perennial River Garry in Scotland
invertebrates trapped in contracting pools or stranded experienced regular drying due to water diversion for a
on dry streambeds are rich pickings for riparian pre- hydroelectric scheme77; and peak water consumption
dators.66 Equally, riparian and terrestrial organisms may exacerbate widespread discharge reductions dur-
that colonize dry channels may be engulfed when flow ing droughts.74 However, patterns vary depending on
resumes,52 and subsidies to terrestrial environments
other environmental factors including geology: streams
are therefore reciprocated by energy inputs to aquatic
underlain by aquifers with low storage capacity may
webs, supporting early flowing-phase colonists.58
experience severe discharge declines during droughts,
whereas in those supplied by porous aquifers, winter
recharge may sustain summer flows.74 In addition,
INTERACTING STRESSORS
widespread river regulation in temperate regions may
COMPROMISE TEMPORARY STREAM limit natural high- and low-flow extremes, with com-
HEALTH pensation flows released from impoundments main-
Changing rainfall and runoff patterns are altering taining a minimum discharge that reduces
flow regimes,67 although patterns are difficult to intermittence. Agricultural land use can also reduce
characterize68 or to relate to climatic drivers.69 hydrological variability and cause artificial perenniali-
Global-scale models predict future decreases in sum- zation; for example, small agricultural dams release
mer discharge in the northern hemisphere,67 and water downstream at a steady rate for irrigation.78
some studies indicate future declines in mean annual The effects of altered hydrology and other
runoff in oceanic western Europe,70 causing summer human influences on instream communities may be
discharge reductions.69,71 Hydrological extremes considerable, especially when artificial temporary or
including drought (i.e., significantly below-average perennial streams are created. Increases in intermit-
water availability over an extended period72) may tence typically reduce aquatic biodiversity, as com-
also be increasing in some global73 and European74 munities become subsets of taxa associated with
regions, causing drying of oceanic rivers previously perennial flow12. Contrary to common perceptions,79
considered perennial.41 However, the inherent varia- artificial perennialization also reduces ecological
bility of drought disturbances and their interaction quality, because TS specialists may be lost as biotic
with nonclimatic drivers of change make both recent interactions with competitors and predators intensify.
trends and future predictions difficult to confirm.69,75 Where lost taxa played important ecosystem roles,
Climatic drivers interact with groundwater and for example as top predators80 or leaf-litter shredders
surface water abstractions in temperate regions that release energy for other feeding groups,61
Volume 4, July/August 2017 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. 9 of 17Opinion wires.wiley.com/water
communities can shift to alternate states.81 Loss of initiatives89 may inform such designations. However,
natural hydrological variability threatens organisms regardless of data availability, TS classification is
adapted to fluctuating conditions, and equally, where challenging because flow regimes are highly variable
shifts between flow extremes become more common, within and between systems and years.
survival within refuges may be compromised.41 Following recognition, mapping, and classifica-
tion of TS, the communities characterizing unim-
pacted ecological quality (i.e., WFD ‘biological
REGULATORY MONITORING quality elements’ indicative of reference conditions90;
SHOULD BETTER REPRESENT Habitats Directive ‘qualifying features’ of favorable
status) should be established, to facilitate robust
TEMPORARY STREAMS future status assessments. However, reference condi-
The EU Water Framework Directive (WFD) requires tions are conceptualized as a single benchmark
EU Member States to attain at least ‘good ecological against which other water bodies can be compared,
status’ in surface water bodies, with status deter- which may be inadequate to represent TS: ecosystems
mined by monitoring to compare sites with unim- that, by definition, transition between lotic, lentic,
pacted ‘reference conditions’ for characterized river and terrestrial conditions in both space and time.
typologies. Equally, designation as a protected Spe- Recent Mediterranean research initiatives87,91,92 have
cial Area of Conservation (SAC) under the EU Habi- made recommendations that may inform adaptation
tats Directive requires monitoring and reporting of of biomonitoring programs in oceanic regions to
the conservation status of Annex I habitats and characterize peak aquatic community diversity
Annex II species. SACs include intermittent river despite this variability. Specifically, the use of estab-
reaches, for example, some chalk streams in southern lished perennial-stream metrics may be appropriate
England are designated for their plant communities in TS flowing-phase assessments, if flowing phases
and for individual invertebrate and fish species. The are sufficiently long and predictable to allow sam-
EU Biodiversity Strategy to 2020 has set specific tar- pling that coincides with peak diversity92 and to
gets for the attainment of ‘favorable’ SAC conserva- allow robust status determination based only on lotic
tion status, determined for habitats by comparison assemblages.
with natural ecosystem ‘structure and function’. However, sensitivity to intermittence and to
In these policy and legislative contexts, recogni- environmental degradation typically covary, mean-
tion and mapping of TS represents a fundamental ing that metrics developed to quantify community
first step towards their monitoring and protection in integrity in perennial streams may underestimate the
and beyond temperate zones.82 Another key priority ecological quality of TS, particularly where flowing
is classification of TS into ecologically robust typolo- phases are short and unpredictable.93 Equally, per-
gies, including discrimination between artificial and ennial metrics may overestimate TS ecological status
natural perennial and intermittent flow.82–84 Ecologi- if applied to samples collected as flow recession
cally relevant classification should recognize the flow forces organisms (whose persistence is threatened by
regime, facilitated by catchment-scale hydrological declining water quality) to share a shrinking sub-
monitoring that includes intermittent reaches and merged habitat area. Development of TS-specific
that differentiates between lentic and dry no-flow indices is therefore required,92 and balanced, robust
states. However, underrepresentation of TS in gau- ecological status assessments should employ novel
ging station networks,85 limited characterization of multi-metric indices that integrate community-level
longitudinal hydrological variability, and difficulties data encompassing the temporal (i.e., lotic, lentic,
in distinguishing between different no-flow states and terrestrial phase) and spatial (e.g., longitudinal)
make such informative, long-term hydrological data variability characteristic of TS, and that use hydro-
scarce. In addition, the indices used to classify hydro- logical data to inform interpretation of sampled
logical regimes have been developed for perennial assemblages. Research priorities include the develop-
streams86 and therefore require supplementation in ment of dry-phase biomonitors, with terrestrial
TS by new descriptors of the magnitude, frequency, invertebrates suggested as one potential indicator of
duration, timing, and rate of change for flow- TS dry-phase health.23 Functional as well as struc-
cessation and drying events.85,87 Qualitative descrip- tural (i.e., taxonomic) approaches to community
tion of TS typologies based on expert opinion may characterization should be investigated,84 and envi-
be a necessary interim measure to facilitate TS moni- ronmental DNA is a potential game-changer that
toring; hydrologically relevant environmental integrates terrestrial and aquatic biodiversity
datasets,88 remote sensing data,87 and citizen science information.94
10 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017WIREs Water Temporary streams in temperate zones
In association with the development of new TS- dense vegetation that maintains seedbanks may limit
specific indices, national regulatory agencies may seek instream macrophyte assemblages.42 Bankside vege-
to expand regulatory monitoring networks to tation also provides habitat for a riparian fauna
improve representation of TS,95 firstly, to recognize including potential dry-channel colonists, and, as in
that multiple monitoring sites may be needed to repre- perennial systems, stabilizes banks and intercepts pol-
sent variation in environmental conditions and cate- luted water. The wider services provided by riparian
gorized ecological status of water bodies with both vegetation mean that planting schemes implemented
perennial and intermittent reaches. Secondly, expan- with nonecological goals (e.g., flood mitigation; cli-
sion may be appropriate to better represent headwater mate change adaptation) may have serendipitous
streams, which, despite their recognized biodiversity effects on TS ecosystem health.
value and ecosystem service provision,47,96 may be Projects involving physical alteration of the
excluded from WFD-related monitoring programs streambed and banks (e.g., sediment manipulation,
due to their size: the WFD target of good ecological channel remeandering, bank reprofiling, and bed
status applies to ‘all bodies of surface water,’97 but raising) may be implemented to restore the natural
streams with catchment areasOpinion wires.wiley.com/water
met. The accuracy of such assessments may be landscape-level influences. In contrast, headwater
improved through recognition of taxon-specific co- catchments with minimal anthropogenic land use
sensitivity to intermittence and environmental may provide opportunities to maximize biodiversity
quality,93 and of broad-scale influences on commu- gains.107 Collaborative, interdisciplinary, interna-
nity composition including metacommunity dynamics tional projects that unite academic researchers, pol-
and catchment land uses.31,106 Where interventions icymakers, and regulatory agencies will bring
to enhance ecological quality are required, these are together currently fragmented knowledge and begin
also most effective (but most challenging) when to address the research and management challenges
implemented at broad spatial scales that recognize that TS present.107,108
ACKNOWLEDGMENTS
We thank the British Ecological Society Aquatic Group and Nottingham Trent University, respectively, for
funding and hosting a meeting on temporary rivers and streams in June 2016. We also thank meeting partici-
pants, as well as Dr. Katie Smith, for contributing to useful discussions which informed development of ideas
in this manuscript. This work was informed by the authors’ involvement in COST Action CA15113 (SMIRES,
Science and Management of Intermittent Rivers and Ephemeral Streams, www.smires.eu), supported by COST
(European Cooperation in Science and Technology). The views expressed in this paper are those of the authors
and do not necessarily represent the views of their organizations.
FURTHER READING
COST (European Cooperation in Science and Technology). Action CA151113 Science and Management of Intermittent
Rivers and Ephemeral Streams (SMIRES); 2016. Available at: http://www.cost.eu/COST_Actions/ca/CA15113 and http://
www.smires.eu/ (Accessed November 29, 2016).
The 1000 Intermittent Rivers Project. Available at: http://1000_intermittent_rivers_project.irstea.fr/ (Accessed November
29, 2016).
Intermittent River Biodiversity Analysis & Synthesis. Available at: http://irbas.cesab.org/ (Accessed April 17, 2017).
REFERENCES
1. Leigh C, Boulton AJ, Courtwright JL, Fritz K, 5. Dell AI, Alford RA, Pearson RG. Intermittent pool
May CL, Walker RH, Datry T. Ecological research beds are permanent cyclic habitats with distinct wet,
and management of intermittent rivers: an historical moist and dry phases. PLoS One 2014, 9:e108203.
review and future directions. Freshwater Biol 2016, https://doi.org/10.1371/journal.pone.0108203.
61:1181–1199. https://doi.org/10.1111/fwb.12646.
6. Shute J. Why Britain’s chalk rivers are under threat
2. Rolls RJ, Heino J, Chessman BC. Unravelling the joint [online]; 2016. Jersey, UK: Telegraph Media Group.
effects of flow regime, climatic variability and dispersal Available at: http://www.telegraph.co.uk/news/
mode on beta diversity of riverine communities. Fresh- 2016/06/06/why-britains-chalk-rivers-are-under-threat/.
water Biol 2016, 61:1350–1364. https://doi.org/10. (Accessed December 18, 2016).
1111/fwb.12793.
7. Goulsbra C, Evans M, Lindsay J. Temporary streams
3. Larned ST, Datry T, Arscott DB, Tockner K. Emerging in a peatland catchment: pattern, timing, and controls
concepts in temporary-river ecology. Freshwater Biol on stream network expansion and contraction. Earth
2010, 55:717–738. https://doi.org/10.1111/j.1365- Surf Proc Land 2014, 39:790–803. https://doi.org/10.
2427.2009.02322.x. 1002/esp.3533.
4. Datry T, Corti R, Philippe M. Spatial and temporal
8. Meyer JL, Strayer DL, Wallace JB, Eggert SL,
aquatic–terrestrial transitions in the temporary Albar-
Helfman GS, Leonard NE. The contribution of head-
ine River, France: responses of invertebrates to experi-
water streams to biodiversity in river networks. J Am
mental rewetting. Freshwater Biol 2012, 57:716–727.
Water Resour Assoc 2007, 43:86–103. https://doi.org/
https://doi.org/10.1111/j.1365-2427.2012.02737.x.
10.1111/j.1752-1688.2007.00008.x.
12 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017WIREs Water Temporary streams in temperate zones
9. Mainstone CP, Holmes NT, Armitage PD, Hydrobiologia 2016. https://doi.org/10.1007/s10750-
Wilson AM, Marchant JH, Evans K, Solomon D. 016-28
Chalk rivers – nature conservation and management; 56-x.
1999. WRc Report to English Nature and the Environ- 21. Leberfinger K, Herrmann J. Secondary production of
ment Agency, Medmenham, 184. invertebrate shredders in open-canopy, intermittent
10. Larned ST, Hicks DM, Schmidt J, Davey AJ, Dey K, streams on the island of Öland, southeastern Sweden.
Scarsbrook M, Arscott DB, Woods RA. The Selwyn J N Am Benthol Soc 2010, 29:934–944. https://doi.
River of New Zealand: a benchmark system for allu- org/10.1899/09-179.1.
vial plain rivers. River Res Appl 2008, 24:1–21.
22. Steward AL, Marshall JC, Sheldon F, Harch B,
https://doi.org/10.1002/rra.1054.
Choy S, Bunn SE, Tockner K. Terrestrial invertebrates
11. Jarvie HP, Neal C, Withers PJ. Sewage-effluent phos- of dry river beds are not simply subsets of riparian
phorus: a greater risk to river eutrophication than agri- assemblages. Aquat Sci 2011, 73:551–566. https://doi.
cultural phosphorus? Sci Total Environ 2006, org/10.1007/s00027-011-0217-4.
360:246–253. https://doi.org/10.1016/j.scitotenv.2005.
23. Steward AL, von Schiller D, Tockner K, Marshall JC,
08.038.
Bunn SE. When the river runs dry: human and ecologi-
12. Datry T, Larned ST, Fritz KM, Bogan MT, Wood PJ, cal values of dry riverbeds. Front Ecol Environ 2012,
Meyer EI, Santos AN. Broad-scale patterns of inverte- 10:202–209. https://doi.org/10.1890/110136.
brate richness and community composition in tempo-
rary rivers: effects of flow intermittence. Ecography 24. Corti R, Datry T. Terrestrial and aquatic invertebrates
2014, 37:94–104. https://doi.org/10.1111/j.1600-0587. in the riverbed of an intermittent river: parallels and
2013.00287.x. contrasts in community organisation. Freshwater Biol
2016, 61:1308–1320. https://doi.org/10.1111/fwb.
13. Welborn GA, Skelly DK, Werner EE. Mechanisms 12692.
creating community structure across a freshwater habi-
tat gradient. Annu Rev Ecol Syst 1996, 1:337–363. 25. Sadler JP, Bell D, Fowles A. The hydroecological con-
https://doi.org/10.1146/annurev.ecolsys.27.1.337. trols and conservation value of beetles on exposed riv-
erine sediments in England and Wales. Biol Conserv
14. Smith H, Wood PJ, Gunn J. The influence of habitat
2004, 118:41–56. https://doi.org/10.1016/j.biocon.
structure and flow permanence on invertebrate com-
2003.07.007.
munities in karst spring systems. Hydrobiologia 2003,
510:53–66. https://doi.org/10.1023/B:HYDR.000000 26. Meyer A, Meyer EI, Meyer C. Lotic communities of
8501.55798.20. two small temporary karstic stream systems (East
Westphalia, Germany) along a longitudinal gradient of
15. Holmes NT. Recovery of headwater stream flora fol-
hydrological intermittency. Limnologica 2003,
lowing the 1989–1992 groundwater drought. Hydrol
33:271–279. https://doi.org/10.1016/S0075-9511(03)
Process 1999, 13:341–354. https://doi.org/10.1002/
80022-1.
(SICI)1099-1085(19990228)13:3
3.0.CO;2-L. 27. Řeznícková P, Pařil P, Zahrádková S. The ecological
16. Hammett MJ. Nemoura lacustris Pictet, 1865 effect of drought on the macroinvertebrate fauna of a
(Plecoptera: Nemouridae) – an addition to the British small intermittent stream – an example from the Czech
list. Entomologist’s Mon Mag 2012, 148:43–45. Republic. Int Rev Hydrobiol 2007, 92:514–526.
https://doi.org/10.1002/iroh.200610997.
17. Bratton JH. A review of the scarcer Ephemeroptera
and Plecoptera of Great Britain. Research & Survey in 28. Dieterich M, Anderson NH. The invertebrate fauna of
Nature Conservation. Nature Conservancy Council, summer-dry streams in western Oregon. Arch Hydro-
No. 29; 2010. Peterborough, UK: Joint Nature Con- biol 2000, 147:273–295. https://doi.org/10.1127/
servation Committee. Available at: http://jncc.defra. archiv-hydrobiol/147/2000/273.
gov.uk/page-2552 (Accessed March 12, 2017). 29. Wissinger SA, Greig H, McIntosh A. Absence of spe-
18. Chadd R, Extence C. The conservation of freshwater cies replacements between permanent and temporary
macroinvertebrate populations: a community-based lentic communities in New Zealand. J N Am Benthol
classification scheme. Aquat Conserv 2004, Soc 2009, 28:12–23. https://doi.org/10.1899/08-007.1.
14:597–624. https://doi.org/10.1002/aqc.630. 30. Datry T, Bonada N, Heino J. Towards understanding
19. Armitage PD, Bass J. Long-term resilience and short- the organisation of metacommunities in highly
term vulnerability of South Winterbourne macroinver- dynamic ecological systems. Oikos 2016,
tebrates. Proc Dorset Nat Hist Archaeol Soc 2013, 125:149–159. https://doi.org/10.1111/oik.02922.
134:43–55. 31. Li F, Sundermann A, Stoll S, Haase P. A newly devel-
20. Hill MJ, Death RG, Mathers KL, Ryves DB, White JC, oped dispersal capacity metric indicates succession of
Wood PJ. Macroinvertebrate community composition benthic invertebrates in restored rivers. Sci Total Envi-
and diversity in ephemeral and perennial ponds on ron 2016, 569–570:1570–1578. https://doi.org/10.
unregulated floodplain meadows in the UK. 1016/j.scitotenv.2016.06.251.
Volume 4, July/August 2017 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. 13 of 17Opinion wires.wiley.com/water
32. Schriever TA, Lytle DA. Convergent diversity and trait 44. Soininen J, Bartels P, Heino J, Luoto M, Hillebrand H.
composition in temporary streams and ponds. Eco- Toward more integrated ecosystem research in aquatic
sphere 2016, 7:e01350. https://doi.org/10.1002/ and terrestrial environments. BioScience 2015,
ecs2.1350. 65:174–182. https://doi.org/10.1093/biosci/biu216.
33. Korhonen JJ, Soininen J, Hillebrand H. A quantitative 45. Sánchez-Montoya MM, Moleón M, Sánchez-
analysis of temporal turnover in aquatic species assem- Zapata JA, Tockner K. Dry riverbeds: corridors for
blages across ecosystems. Ecology 2010, 91:508–517. terrestrial vertebrates. Ecosphere 2016, 7:e01508.
https://doi.org/10.1890/09-0392.1. https://doi.org/10.1002/ecs2.1508.
34. Bogan MT, Lytle DA. Seasonal flow variation allows 46. Coetzee CG. The distribution of mammals in the
‘time-sharing’ by disparate aquatic insect communities Namib desert and adjoining inland escarpment. Sci
in montane desert streams. Freshwater Biol 2007, Pap Namib Desert Res Station 1969, 40:23–36.
52:290–304. https://doi.org/10.1111/j.1365-2427. 47. Acuña V, Hunter M, Ruhí A. Managing temporary
2006.01691.x. streams and rivers as unique rather than second-class
35. Perrow M, Leeming D, England J, Tomlinson M. Life ecosystems. Biol Conserv 2017. https://doi.org/10.
after low flow–ecological recovery of the River Mis- 1016/j.biocon.2016.12.025.
bourne. British Wildlife 2007, 18:335–346. 48. Cooper SM, Perotto-Baldivieso HL, Owens MK,
Meek MG, Figueroa-Pagan M. Distribution and inter-
36. Vander Vorste R, Malard F, Datry T. Is drift the pri-
action of white-tailed deer and cattle in a semi-arid
mary process promoting the resilience of river inverte-
grazing system. Agric Ecosyst Environ 2008,
brate communities? A manipulative field experiment in
127:85–92. https://doi.org/10.1016/j.agee.2008.03.0
an intermittent alluvial river. Freshwater Biol 2016,
04.
61:1276–1292. https://doi.org/10.1111/fwb.12658.
49. Lugon-Moulin N, Hausser J. Phylogeographical struc-
37. Bogan MT, Boersma KS, Lytle DA. Resistance and
ture, postglacial recolonization and barriers to gene
resilience of invertebrate communities to seasonal and
flow in the distinctive Valais chromosome race of the
supraseasonal drought in arid-land headwater streams.
common shrew (Sorex araneus). Mol Ecol 2002,
Freshwater Biol 2015, 60:2547–2558. https://doi.org/
11:785–794. https://doi.org/10.1046/j.1365-294X.
10.1111/fwb.12522.
2002.01469.x.
38. Feminella JW. Comparison of benthic macroinverte- 50. Mockford SW, Herman TB, Snyder M, Wright JM.
brate assemblages in small streams along a gradient of Conservation genetics of Blanding’s turtle and its
flow permanence. J N Am Benthol Soc 1996, application in the identification of evolutionarily signif-
15:651–669. https://doi.org/10.2307/1467814. icant units. Conserv Genet 2007, 8:209–219. https://
39. Sánchez-Montoya MM, von Schiller D, Ruhí A, doi.org/10.1007/s10592-006-9163-4.
Pechar GS, Proia L, Miñano J, Vidal-Abarca MR, 51. Storfer A, Murphy MA, Spear SF, Holderegger R,
Suárez ML, Tockner K. Responses of ground-dwelling Waits LP. Landscape genetics: where are we now? Mol
arthropods to surface flow drying in channels and Ecol 2010, 19:3496–3514. https://doi.org/10.1111/j.
adjacent habitats along Mediterranean streams. Ecohy- 1365-294X.2010.04691.x.
drology 2016, 9:1376–1387. https://doi.org/10.1002/
52. Corti R, Datry T. Invertebrates and sestonic matter in
eco.1733.
an advancing wetted front travelling down a dry river
40. Stubbington R, Datry T. The macroinvertebrate seed- bed (Albarine, France). Freshwater Sci 2012,
bank promotes community persistence in temporary 31:1187–1201. https://doi.org/10.1899/12-017.1.
rivers across climate zones. Freshwater Biol 2013, 53. Jacobson PJ, Jacobson KN, Seely MK. Ephemeral Riv-
58:1202–1220. https://doi.org/10.1111/fwb.12121. ers and their Catchments: Sustaining People and
41. Stubbington R, Gunn J, Little S, Worrall TP, Wood PJ. Development in Western Namibia. Windhoek,
Macroinvertebrate seedbank composition in relation to Namibia: Desert Research Foundation of Namibia;
antecedent duration of drying and multiple wet-dry 1995, 164.
cycles in a temporary stream. Freshwater Biol 2016, 54. O’Neal BJ, Stafford JD, Larkin RP. Migrating ducks in
61:1293–1307. https://doi.org/10.1111/fwb.12770. inland North America ignore major rivers as leading
42. Westwood CG, Teeuw RM, Wade PM, Holmes NT, lines. Ibis 2015, 157:154–161. https://doi.org/10.1111/
Guyard P. Influences of environmental conditions on ibi.12193.
macrophyte communities in drought-affected headwa- 55. Bogan MT, Boersma KS, Lytle DA. Flow intermittency
ter streams. River Res Appl 2006, 22:703–726. https:// alters longitudinal patterns of invertebrate diversity
doi.org/10.1002/rra.934. and assemblage composition in an arid-land stream
43. Fenchel TO, Finlay BJ. The ubiquity of small species: network. Freshwater Biol 2013, 58:1016–1028.
patterns of local and global diversity. Bioscience 2004, https://doi.org/10.1111/fwb.12105.
54:777–784. https://doi.org/10.1641/0006-3568(2004) 56. Verdonschot R, van Oosten-Siedlecka AM, ter
054[0777:TUOSSP]2.0.CO;2. Braak CJ, Verdonschot PF. Macroinvertebrate survival
14 of 17 © 2017 The Authors. WIREs Water published by Wiley Periodicals, Inc. Volume 4, July/August 2017You can also read

















































