SPX4 Acts on PHR1-Dependent and -Independent Regulation of Shoot Phosphorus Status in Arabidopsis1 OPEN - Plant Physiology
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
SPX4 Acts on PHR1-Dependent and -Independent
Regulation of Shoot Phosphorus Status
in Arabidopsis1[OPEN]
Marina Borges Osorio,a Sophia Ng,b,2 Oliver Berkowitz,a Inge De Clercq,c,d Chuanzao Mao,e Huixia Shou,e
James Whelan,a and Ricarda Jost a,3,4
a
Department of Animal, Plant, and Soil Sciences and Centre for AgriBioscience, Australian Research Council
Centre of Excellence in Plant Energy Biology, School of Life Sciences, La Trobe University, Bundoora, Victoria
3086, Australia
b
Australian Research Council Centre of Excellence in Plant Energy Biology, University of Western Australia,
Perth, Western Australia 6009, Australia
c
Ghent University, Department of Plant Biotechnology and Bioinformatics, 9052 Ghent, Belgium
d
VIB Center for Plant Systems Biology, 9052 Ghent, Belgium
e
State Key Laboratory of Plant Physiology and Biochemistry, College of Life Sciences, Zi Jin Gang Campus,
Zhejiang University, Hangzhou 310058, China
ORCID IDs: 0000-0003-4024-9233 (M.B.O.); 0000-0002-1677-863X (S.N.); 0000-0002-7671-6983 (O.B.); 0000-0001-8125-1239 (I.D.C.);
0000-0001-5126-2180 (C.M.); 0000-0001-6890-5672 (H.S.); 0000-0001-5754-025X (J.W.); 0000-0002-3819-6358 (R.J.).
Phosphorus (P) is an essential macronutrient for all living organisms and limits plant growth. Four proteins comprising a single
SYG1/Pho81/XPR1 (SPX) domain, SPX1 to SPX4, are putative phosphate-dependent inhibitors of Arabidopsis (Arabidopsis
thaliana) PHOSPHATE STARVATION RESPONSE1 (PHR1), the master transcriptional activator of phosphate starvation
responses. This work demonstrated that SPX4 functions as a negative regulator not only of PHR1-dependent but also of
PHR1-independent responses in P-replete plants. Transcriptomes of P-limited spx4 revealed that, unlike SPX1 and SPX2,
SPX4 modulates the shoot phosphate starvation response but not short-term recovery after phosphate resupply. In roots,
transcriptional regulation of P status is SPX4 independent. Genes misregulated in spx4 shoots intersect with both PHR1-
dependent and PHOSPHATE2-dependent signaling networks associated with plant development, senescence, and ion/
metabolite transport. Gene regulatory network analyses suggested that SPX4 interacts with transcription factors other than
PHR1, such as SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 and ARABIDOPSIS NAC DOMAIN CONTAINING
PROTEIN55, known regulators of shoot development. Transient expression studies in protoplasts indicated that PHR1 retention
in the cytosol by SPX4 occurs in a dose- and P-status-dependent manner. Using a luciferase reporter in vivo, SPX4 expression
kinetics and stability revealed that SPX4 is a short-lived protein with P-status-dependent turnover. SPX4 protein levels were
quickly restored by phosphate resupply to P-limited plants. Unlike its monocot ortholog, AtSPX4 was not stabilized by the
phosphate analog phosphite, implying that intracellular P status is sensed by its SPX domain via phosphate-rich metabolite
signals.
Phosphorus (P) is an essential macronutrient for all achieving sustainable P use in crop plants has become a
living organisms. Apart from nitrogen, it is the most major goal in plant breeding (Veneklaas et al., 2012;
limiting factor for plant growth and development in Heuer et al., 2017).
both natural and agricultural ecosystems. Phosphate To cope with Pi scarcity, plants have evolved a range
(Pi), the inorganic form of P, that is taken up and me- of adaptations, collectively known as the Pi-starvation
tabolized by plants, is a key substrate for most bio- response (PSR; Raghothama, 1999). PSR typically aims
chemical reactions in the cell. Even though P can be at increasing Pi uptake from soil particles (e.g. through
abundant in soils, it is mostly unavailable to plants due modifications in root system architecture alongside
to the conversion of Pi into organic compounds by mi- enhanced exudation of carboxylates and phosphatases)
croorganisms (Richardson and Simpson, 2011) and or maximizing Pi utilization, through increased Pi
fixation to soil particles (Hinsinger, 2001). In order to translocation from shoot to root, release from vacuolar
maintain agricultural productivity and provide food stores, and remobilization within tissues (Raghothama,
security for a growing global population, our already 1999). Two common root adaptations to increase Pi
heavy reliance on Pi fertilizers is bound to increase. Yet, uptake in higher plants (i.e. cluster-root formation and
this practice is neither economically nor environmen- mycorrhizal associations; Péret et al., 2014) are absent in
tally sustainable, as it leads to depletion of natural re- the Arabidopsis (Arabidopsis thaliana) model, yet this
sources and eutrophication of water bodies. Hence, model has allowed the identification of key regulators
332 Plant PhysiologyÒ, September 2019, Vol. 181, pp. 332–352, www.plantphysiol.org Ó 2019 American Society of Plant Biologists. All Rights Reserved.
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.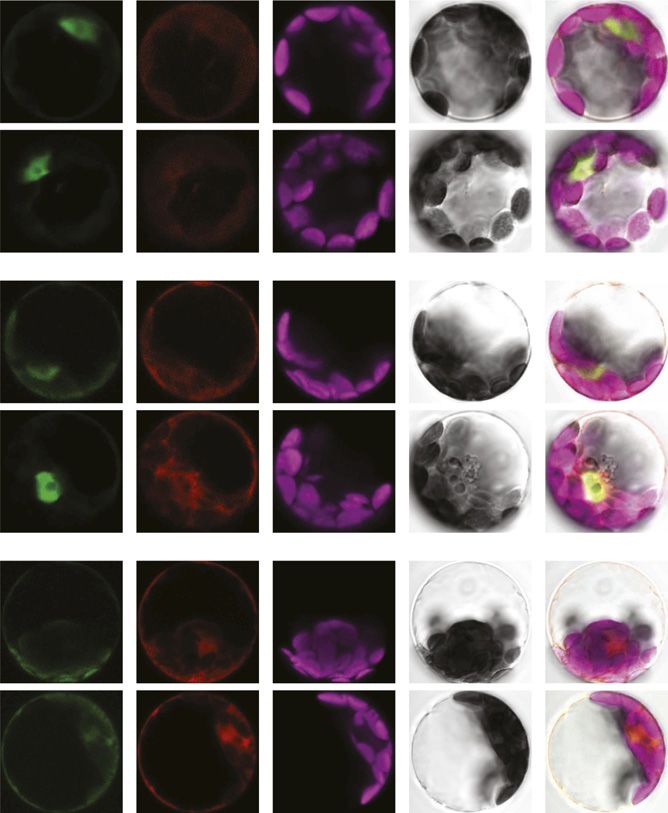
AtSPX4 Regulates Intracellular P Status in Shoots
of most acclimation responses (Rubio et al., 2001; Aung networks that regulate PSR in different plant organs
et al., 2006; Bari et al., 2006; Chen et al., 2007; Devaiah et (Wu et al., 2003; Devaiah et al., 2009) or biochemical
al., 2007, 2009; Camacho-Cristóbal et al., 2008; Duan pathways (Acevedo-Hernández et al., 2012).
et al., 2008; Bustos et al., 2010; Lundmark et al., 2011; Information on the molecular basis of P sensing and
Arpat et al., 2012; Puga et al., 2014; Chen and Schmidt, early signaling upstream of PHR1 is scarce (Abel et al.,
2015; Baek et al., 2017). The discoveries that there are 2002). While PHR1 transcripts are regulated by light
also negative regulators of PSR in P-sufficient plants and ethylene signals (Liu et al., 2017), they are not
(Mukatira et al., 2001) and that many PSR components directly affected by changes in Pi availability (Rubio
are shut down within hours of Pi resupply (Burleigh et al., 2001). Instead, PHR1 activity seems to rely on
and Harrison, 1999; Müller et al., 2004; Woo et al., additional layers of posttranscriptional regulation,
2012; Secco et al., 2013) pose a challenge to improving including SUMOylation by SAP AND MIZ1 (Miura
Pi uptake in agricultural systems: due to negative et al., 2005) and interaction with SYG1/PHO81/
feedback loops acting on root Pi uptake, crops such as XPR1 (SPX) domain-containing proteins (Lv et al.,
wheat (Triticum aestivum), oat (Avena sativa), or maize 2014; Puga et al., 2014; Wang et al., 2014; Qi et al.,
(Zea mays) currently use only between 10% and 30% 2017).
of the Pi fertilizer applied (Bezzola et al., 1994; The latter form a group of four proteins in Arabi-
Glendinning and Fertilizer Industry Federation of dopsis, SPX1 to SPX4, made up of a single SPX domain
Australia, 2000). It is hence of utmost importance to (Duan et al., 2008). Transcripts of SPX1, SPX2, and
gain a better understanding of the role of these negative SPX3 accumulate in roots and shoots of P-limited
regulators in modulating P efficiency in plants. plants in a PHR1-dependent manner (Duan et al.,
Transcriptional reprogramming in response to Pi 2008; Bustos et al., 2010). SPX4 transcripts, on the
starvation in Arabidopsis is largely regulated by the other hand, show only a weak suppression by Pi
GARP family of R2R3 MYB DOMAIN PROTEIN (MYB) withdrawal or knockout of PHR1 (Duan et al., 2008).
transcription factors, namely PHOSPHATE STARVA- Nucleus-localized SPX1 binds to and inhibits PHR1
TION RESPONSE1 (PHR1), PHR1-LIKE1 (PHL1), and activity in the presence of Pi (Puga et al., 2014). Similar
PHL2 (Rubio et al., 2001; Sun et al., 2016). Regulatory findings have been reported for the interaction of rice
regions of almost all PSR genes are greatly enriched in (Oryza sativa) orthologs, OsSPX1 and OsSPX2, with the
the P1BS cis-acting motif, and the absence of PHR1 and PHR1 ortholog OsPHR2 (Wang et al., 2014). Arabi-
PHL1 leads to misregulation of about 75% and 65% of dopsis SPX1 binds to monomeric PHR1 in the presence
Pi-starvation-induced and -repressed genes, respec- of either 5 mM Pi or 50 mM inositol hexakisphosphate
tively (Bustos et al., 2010). Other MYB factors such as in vitro but not to PHR1 dimers bound to twin P1BS-
MYB62 (Devaiah et al., 2009), GARP COILED-COIL7 binding sites (Qi et al., 2017). Eukaryotic SPX domains
(GCC7; Lundmark et al., 2011), HRS1 HOMOLOG2, were found to directly bind to inositol polyphosphate
and three R3-type MYBs (Chen and Schmidt, 2015) signaling molecules (IPs) in response to shifts in cellular
have been implicated in a number of PHR1-dependent Pi levels (Wild et al., 2016), positioning SPX proteins as
and -independent Pi starvation responses, indicating prime candidates for P(i) sensors in plants. In both
both functional redundancy and/or discrete regulatory Arabidopsis and rice, SPX1 and SPX2 display a signif-
icant degree of redundancy, with only double knockout
mutants showing substantial accumulation of Pi in
1
This work was supported by the Department of Industry, Inno- shoots and changes in the expression of PHR1-
vation, Science, Research, and Tertiary Education, Australian Gov- dependent PSR genes (Puga et al., 2014; Wang et al.,
ernment/Australian Research Council Centre of Excellence for Plant 2014). Shoot accumulation of Pi in OsPHR2 over-
Energy Biology (CE140100008). expression lines in rice could be reversed by simulta-
2
Current address: French Alternative Energies and Atomic Energy
neous overexpression of OsSPX1; this was attributed to
Comission, Cadarache, Institute of Biosciences and Biotechnologies,
Biology of Plant Development, F-13108 St-Paul-lez-Durance, France.
differential expression of the Pi transporter gene OsPT2
3
Author for contact: r.jost@latrobe.edu.au. in the root of these lines (Liu et al., 2010).
4
Senior author. PHOSPATE2 (PHO2)/UBIQUITIN-CONJUGAT-
The author responsible for distribution of materials integral to the ING ENZYME24, a ubiquitin E2 conjugase, is another
findings presented in this article in accordance with the policy de- well-characterized repressor of select PSR gene function
scribed in the Instructions for Authors (www.plantphysiol.org) is: in roots. PHO2 regulates root Pi uptake and its trans-
James Whelan (j.whelan@latrobe.edu.au). location to shoots via targeting of Pi transporter pro-
R.J., H.S., C.M., and J.W. conceived the project; M.B.O. carried out teins of the PHOSPHATE TRANSPORTER1 (PHT1)
the physiological characterization of the mutants, phosphate, and family (Huang et al., 2013; Park et al., 2014) as well as of
reporter gene assays; S.N. genotyped the spx4 mutants and generated
the Pi exporter PHO1 (Liu et al., 2012) for proteasomal
homozygous SPX4-LUC lines; R.J. performed the transient subcellu-
lar localization assays; I.D.C. conducted the regulatory network anal-
degradation. The complex down-regulation of PHO2
yses; O.B. carried out the RNA-seq, confocal microscopy, and transcripts in roots by select shoot-derived micro-
bioinformatic data analyses; R.J., J.W., and M.B.O. interpreted results RNA399 species and their proposed sequestration by
and drafted the article; all authors reviewed the article. antagonistic noncoding small RNAs INDUCED BY PI
[OPEN]
Articles can be viewed without a subscription. STARVATION1 (IPS1) and IPS2/AT4 are known to
www.plantphysiol.org/cgi/doi/10.1104/pp.18.00594 occur downstream of PHR1 (Fujii et al., 2005; Aung
Plant Physiol. Vol. 181, 2019 333
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.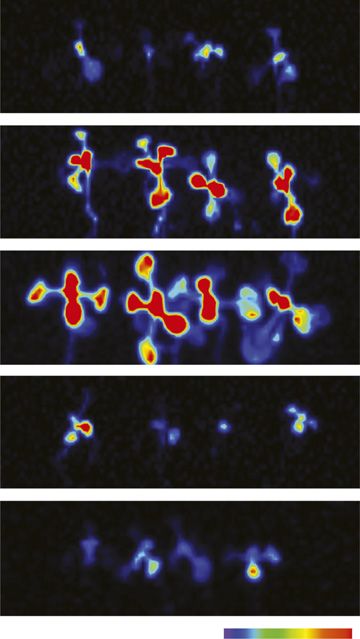
Osorio et al.
et al., 2006; Bari et al., 2006; Franco-Zorrilla et al., 2007; roots and shoots, going back to control levels within
Pant et al., 2008). In rice, both Ospho2 knockout and 24 h of Pi resupply (Duan et al., 2008; Fig. 1H). A similar
OsmiR399k overexpression lines showed enhanced expression pattern was described for the rice ortholog
transcript accumulation of OsSPX1 in roots but not OsSPX4 (Lv et al., 2014). In vitro, OsSPX4 protein levels
shoots (Liu et al., 2010). OsPHO2 expression in turn was fluctuate with Pi availability, with 26S proteasome-
also higher in roots, not shoots, of OsSPX1 over- dependent degradation dominating under P defi-
expression lines. Liu et al. (2010) concluded at the time ciency (Lv et al., 2014). To determine the stability of
that a root-specific negative feedback loop existed be- AtSPX4 in vivo, a SPX4pro:SPX4-LUC reporter line was
tween OsSPX1 and OsPHO2. The role of PHO2 in created in which the coding region of SPX4 is transla-
shoots, however, remains largely unknown, with evi- tionally fused to firefly luciferase (LUC) under the
dence suggesting a possible role in flowering time control of its native promoter, allowing us to monitor
regulation in both Arabidopsis and rice (Kant et al., SPX4 protein kinetics under varying Pi supplies in
2011; Kim et al., 2011; Li et al., 2017). seedlings. Pi resupply to P-starved seedlings led to
The function of a third negative regulator of PSR gene distinct Pi and SPX4-LUC accumulation kinetics in each
expression, SPX4, has recently been studied in rice. Lv organ: in shoots, Pi pools were restored within 2 d of Pi
et al. (2014) found that it, too, interacts with OsPHR2 in resupply (Fig. 1A), marked by a concomitant but slower
a P-status-dependent manner. However, unlike recovery of SPX4-LUC protein and activity (Fig. 1, C
OsSPX1, OsSPX4-GFP fusion protein was detected in and D). In roots, SPX4-LUC levels were restored to
both cytosol and nucleus of transgenic root cells as well those in control plants within 24 h of Pi resupply but
as in rice protoplasts. OsSPX4 was also shown to inhibit increased further until day 3 of Pi resupply (Fig. 1E).
the translocation of OsPHR2 from the cytosol into the This overshooting of SPX4-LUC levels in roots is
nucleus by bimolecular fluorescence complementation counterintuitive, given that Pi accumulation in this or-
assays (Lv et al., 2014). The authors were able to show gan peaks after 1 d of resupply (Fig. 1B). SPX4-LUC
that there is no functional redundancy in rice, given that levels in roots thus appear to be coupled to the stabili-
the spx4 T-DNA insertion line showed Pi accumulation zation of intracellular Pi pools in shoots rather than
and derepression of the PSR gene IPS1 in P-replete local Pi concentration.
shoots (Lv et al., 2014). The fact that this negative reg- SPX4 protein turnover changed in a P-dependent
ulator also undergoes rapid turnover in extracts of manner when SPX4pro:SPX4-LUC seedlings were trea-
P-deficient plants and is stabilized in the presence of ted (via their roots) with cycloheximide (CHX), an in-
either Pi or its analog phosphite (Phi; Lv et al., 2014) hibitor of de novo protein synthesis. LUC activity in
would furthermore suggest its immediate role in the shoots was found to be lower in P-replete and in Pi-r-
sensing of cellular P status. In addition, organ-specific esupply conditions compared with the corresponding
differences between root and shoot regulatory net- mock treatments (0.01% dimethyl sulfoxide [DMSO]).
works have been shown for OsSPX6, which is only This was shown both by LUC imaging (Fig. 1J) and
degraded in P-limited leaves but accumulates in roots quantification of LUC activity (Fig. 1I). Thus, increasing
(Zhong et al., 2018). SPX4-LUC levels in the presence of Pi requires de novo
Given the importance of overcoming negative regu- protein synthesis. Simultaneous CHX treatment of
lators of Pi uptake for increasing P use in plants, and the seedlings upon transfer from P-replete to P-deficient
distinct differences in PSR regulatory networks in medium for 24 h resulted in increased LUC activity
monocots and dicots, we focused on dissecting SPX4- (Fig. 1, I and J, 1d 2P), while 24-h CHX treatment of
dependent gene regulatory networks in Arabidopsis. seedlings already starved of Pi for 4 d resulted in a
Use of a luciferase reporter, protoplast transfection with significant decrease in LUC activity (Fig. 1, I and J, 5d
fluorescent fusion proteins, mutants of spx4, tran- 2P). Reduction of LUC protein and activity levels in
scriptome analyses under Pi-replete, -limited, and CHX-treated plants irrespective of Pi supply indicates
-resupply conditions, and comparison with PHR1 and that faster SPX4 degradation, not reduced de novo
PHO2 master regulators revealed that SPX4 acts as a protein synthesis, is a major factor affecting SPX4
modulator of transcription factor activities in P-replete turnover, ensuring that basal levels of functional SPX4
and P-limited shoots. Potential downstream targets of are still being maintained in P-starved shoots. Protein(s)
SPX4, other than PHR1, and the role of PHO2 are responsible for triggering SPX4 degradation seem to be
discussed. synthesized de novo in the early phase of Pi depletion,
which would explain the higher SPX4 accumulation in
CHX-treated shoots at the 1-d 2P time point compared
with control plants.
RESULTS To demonstrate the contribution of ubiquitin-
SPX4 Is a Short-Lived Protein with P-Dependent Turnover mediated SPX4 degradation, the effect of an inhibitor
of the 26S proteasome, MG132, on SPX4-LUC protein
Among the four Arabidopsis genes encoding single levels in P-starved seedlings was assessed. Unlike the
SPX domain proteins, named SPX1 to SPX4, SPX4 is the stabilizing effect of MG132 treatment on OsSPX4 levels
only one not induced by Pi starvation. In fact, SPX4 in extracts from P-starved rice seedlings (Lv et al., 2014),
transcripts were mildly repressed by Pi deprivation in no differences in LUC activity were observed in vivo by
334 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
AtSPX4 Regulates Intracellular P Status in Shoots
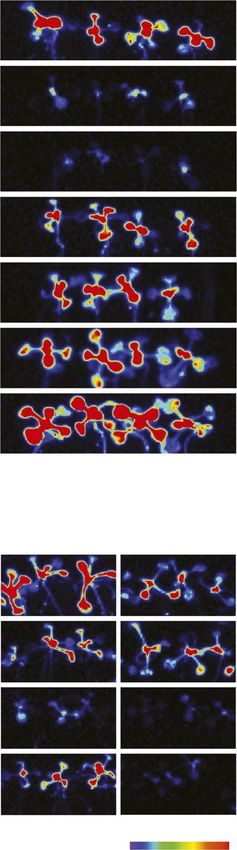
Figure 1. SPX4 protein stability is affected by P-dependent changes in protein turnover. A, B, and D to G, Organ Pi concentration
(A and B), quantification of LUC activity (D and E), and organ biomass (F and G) in seedlings grown under varying Pi supplies. CP
5 control (P-replete), 2P 5 P-limited, and 1P 5 Pi-resupplied plants. Values are means 6 SE of at least three independent bi-
ological replicates, each comprising pools of four to six organs. Asterisks indicate significant differences from P-replete plants (P ,
0.05, ANOVA/Tukey). FW, Fresh weight. H, Relative SPX4 transcript abundance in Col-0 roots and shoots grown under varying Pi
supplies. Relative expression is shown as 402DCt, compared with ACT7, UBC9, and UBC21 reference genes (n 5 3 independent
biological replicates). No significant differences were found across treatments (P , 0.05, ANOVA/Tukey). I, Quantification of
LUC activity in SPX4pro:SPX4-LUC shoots treated for 24 h with DMSO (control; black bars) or 50 mM CHX (red bars) under varying
Pi supplies. Values are means 6 SE of at least three independent biological replicates. Asterisks indicate significant differences
from DMSO treatment for each condition (P , 0.05, ANOVA/Tukey). C and J, LUC imaging of Arabidopsis SPX4pro:SPX4-LUC
shoots over a time course of Pi limitation and resupply (C) and upon CHX treatment (J). Luminescence is shown as units of pixel
intensity.
Plant Physiol. Vol. 181, 2019 335
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Osorio et al.
treating plants with MG132 in combination with CHX of Pi resupply, remaining highly induced at the 3-d time
(Park et al., 2014; Crozet et al., 2016; Supplemental Fig. point (Fig. 2C). LUC activity in Phi-treated shoots, on
S1). E-64d, an inhibitor of endosomal Cys proteases the other hand, was not significantly different (P . 0.05)
(Yamada et al., 2005), was also tested to no effect from that in P-starved shoots. In roots, SPX4 levels
(Supplemental Fig. S1). The divergent results between steadily increased upon Pi resupply, with 10- and 20-
OsSPX4 and AtSPX4 are likely due to our finding that fold increases after 1 and 3 d of treatment, respectively
SPX4 degradation requires de novo protein synthesis in (Fig. 2B). Phi treatment of P-starved roots led to a
Arabidopsis (Fig. 1I, 1d 2P), but the main finding of transient threefold higher LUC activity within 24 h.
P-dependent SPX4 turnover is consistent in both This weak LUC induction in Phi-treated roots coincided
species. with a slight transient increase in root Pi concentration
(Fig. 2D), most likely caused by scavenging of residual
Pi present on fresh agar plates. Our results indicate that
Phi Supply Does Not Mimic Pi Resupply in Restoring SPX4 does not recognize the metabolically inert Pi mi-
SPX4 Levels metic Phi, further supporting the involvement of or-
ganic P molecules, such as IPs, in primary sensing of
Phi (H2PO32) is a more reduced form of P and a Pi intracellular P status via direct binding to the SPX do-
mimetic that cannot be metabolized by plants main (Wild et al., 2016).
(McDonald et al., 2001). It can be used to discriminate
between processes that are directly dependent on Pi and
those that require an organic P metabolite (Jost et al., Coexpression with SPX4 Leads to Dose- and Pi
2015). Hence, we tested its ability to substitute Pi in Treatment-Dependent Retention of PHR1 in the Cytosol
stabilizing SPX4 levels in vivo. We compared the effect
of short-term (24 h) or medium-term (72 h) Pi or Phi In rice, OsSPX4 and OsPHR2 were shown to interact
supply to Pi-starved seedlings grown on vertical plates. in vivo via bimolecular fluorescence complementation
Phi treatment did not recover Pi pools (Fig. 2, D and E; analysis, resulting in retention of OsPHR2 in the cytosol
Jost et al., 2015), leading to a complete arrest of root and (Lv et al., 2014). Here, we wanted to address whether
shoot growth (Fig. 2, F and G). In contrast to OsSPX4 the cytosolic accumulation of PHR1 in Arabidopsis can
(Lv et al., 2014) and AtSPX1 (Puga et al., 2014), AtSPX4 be induced upon AtSPX4 coexpression and whether
levels in P-limited roots and shoots were not restored this depends on SPX4 abundance and/or cellular P
by provision of Phi (Fig. 2, B and C). In P-starved status. Upon transfection of Arabidopsis Columbia-0
shoots, LUC activity increased by fourfold within 24 h (Col-0) protoplasts with a construct expressing a
Figure 2. SPX4 protein levels in
P-limited shoots recover upon Pi but not
Phi treatment. A, Luciferase imaging of
Arabidopsis SPX4pro:SPX4-LUC shoots
comparing the effects of Pi resupply and
Phi supply to P-limited seedlings. Lumi-
nescence is shown as units of pixel in-
tensity. B to G, Quantification of LUC
activity (B and C), tissue Pi concentration
(D and E), and biomass (F and G) in roots
and shoots of P-limited, Pi-resupplied,
and Phi-supplied seedlings. Values are
means 6 SE of at least three independent
biological replicates, comprising pools
of four to six organs each. Asterisks in-
dicate significant differences from
P-limited seedlings grown for 4 d with-
out Pi supply (P , 0.05, ANOVA/Tukey).
FW, Fresh weight.
336 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
AtSPX4 Regulates Intracellular P Status in Shoots
C-terminal SPX4-RFP fusion under the control of the protoplasts would suggest P-status-dependent modifi-
35S promoter, mRFP fluorescence was detected with cation of their interaction.
similar intensity in both cytosol and nucleus, irre-
spective of Pi treatment (Supplemental Fig. S2). This
would suggest that, unlike the SPX4-LUC fusion Disruption of SPX4 Function Leads to Pi Overaccumulation
(Fig. 1), the fluorescent protein tag prevents P-status- in Shoots
dependent SPX4 turnover. This is most likely a reflec-
tion of the better suitability of luciferase as a reporter for In order to assess the physiological effects of dis-
protein turnover, given its much shorter half-life (4 min rupting SPX4, we isolated two T-DNA insertion lines,
to 4 h) compared with, for example, 18 h for GFP in SALK_019826 and SK40726, named spx4-1 and spx4-2,
plants (de Ruijter et al., 2003). Consistent with stably respectively. The molecular characterization of these
transformed 35S:PHR1-GFP lines (Rubio et al., 2001), lines is shown in Supplemental Figure S7. Sequencing
transfection with a 35Spro:PHR1-GFP construct resulted of T-DNA left-border amplicons confirmed that the
in strong nuclear fluorescence, irrespective of Pi supply single insertions in spx4-1 and spx4-2 are in the second
(Supplemental Fig. S3). Coexpression of SPX4-RFP and and third (last) exons of the SPX4 gene, respectively.
PHR1-GFP fusions led to a partial retention of eGFP Both insertions disrupt the third portion of the SPX
fluorescence in the cytosol, but only in P-replete Col-0 tripartite domain (PFam:03105; Supplemental Fig.
protoplasts (Supplemental Fig. S3). In P-limited proto- S7A), upstream of the last amino acid residue forming
plasts, the eGFP signal was almost exclusively found in the phosphate-binding cluster and of the Lys-binding
the nucleus. This differential localization was not ob- cluster, which are active sites involved in binding to IPs
served when the 35Spro:SPX4-RFP construct was coex- (Wild et al., 2016; Azevedo and Saiardi, 2017). SPX4
pressed with a 35S:GFP control (Supplemental Fig. S2). transcripts downstream of the insertion site were not
In P-replete spx4-1 cells, the relative shift in eGFP flu- detected in spx4-1 and were reduced in the weaker spx4-
orescence from nucleus to cytosol upon coexpression of 2 allele compared with the wild type (Supplemental
PHR1-GFP and SPX4-RFP appeared weaker than in Fig. S7C). When mutant alleles were grown side by side
Col-0 protoplasts (Supplemental Fig. S4). Expression of with their respective wild-type backgrounds (Col-0 for
SPX4-RFP on its own resulted in a much stronger nu- spx4-1 and Col-4 for spx4-2) in nutrient-rich soil, an
average of 22% reduction in rosette biomass was ob-
clear signal than in the other two genotypes. In
served (Supplemental Fig. S7D).
P-limited phr1-2 protoplasts, a weak cytosolic PHR1-
For the physiological characterization of the two SPX4
GFP signal was already detected upon single transfec-
mutant alleles under varying Pi supplies, seedlings were
tion with the 35Spro:PHR1-GFP construct (Supplemental
grown vertically on one-half-strength Murashige Skoog
Fig. S5, top). A stronger nuclear SPX4-RFP signal in
(MS) agar plates (see “Materials and Methods”). In ad-
P-limited phr1-2 protoplasts was observed upon PHR1-
dition to wild-type lines (Col-0 and Col-4), mutants
GFP coexpression. These results indicated a fine bal- of PHR1, phr1-1 (Rubio et al., 2001) and phr1-2
ance between SPX4 and PHR1 protein abundance and (SALK_067629), displaying reduced shoot Pi concentra-
their respective subcellular localization. To investigate tion under Pi deficiency (Nilsson et al., 2007; Bustos et al.,
this further, we used different ratios of SPX4-RFP and 2010), and PHO2 (pho2-1/CS8508), known to over-
PHR1-GFP constructs for transfection of spx4-1 proto- accumulate Pi in shoots (Dong et al., 1998), were added
plasts (Fig. 3). With increasing 35Spro:SPX4-RFP con- for comparison. The experiment comprised three con-
struct input, ranging from 0.25-fold to 4-fold excess ditions causing changes in plant Pi status (Fig. 4): under
over 35Spro:PHR1-GFP, the cytosolic eGFP signal be- Pi-replete conditions, seedlings were maintained on
came stronger, while the nuclear signal got fainter. At a sufficient Pi throughout the experiment (including two
1:1 SPX4-to-PHR1 ratio, this response was P-status transfers to fresh medium), thus having fully replete root
dependent (see Supplemental Fig. S4). At a 4:1 SPX4- and shoot Pi pools (Fig. 4, C and D) for optimal plant
to-PHR1 ratio, PHR1-GFP signal was almost exclu- growth (Fig. 4, A and B). Under Pi limitation, 7-d-old
sively cytosolic, irrespective of Pi treatment. A similar seedlings were deprived of Pi for 4 d, followed by an
dose response was observed upon transfection of phr1-2 additional 1 d on fresh P-deficient medium prior to
protoplasts (Supplemental Fig. S6). Together, these harvest. This 5-d treatment was enough to deplete root
data suggest an interaction of AtSPX4 and AtPHR1 in and shoot Pi pools in the wild type, leading to a 50%
the cytosol, as previously observed for the rice ortho- reduction in shoot growth (Fig. 4B). For Pi resupply,
logs (Lv et al., 2014). In contrast to rice, sequestration of P-limited seedlings were transferred to 1 mM Pi for 1 d
AtPHR1 by AtSPX4 in the cytosol is weaker, with both after 4 d of Pi deprivation. At this time point, accelerated
proteins still detected in the nucleus. PHR1 accumula- Pi uptake and root-to-shoot translocation were observed
tion in the cytosol occurs at lower SPX4-to-PHR1 ratios in the wild type, with roots accumulating almost twofold
in P-replete protoplasts, while a large excess of SPX4 more Pi than under continuous Pi supply (Fig. 4C), while
over PHR1 is required to achieve cytosolic retention of shoot Pi pools were 70% replete within 24 h (Fig. 4D),
PHR1 in P-limited cells. The fact that these changes in with shoots just starting to resume growth (Fig. 4B).
subcellular localization of PHR1 occur despite the lack Compared with the wild type, all mutant lines tested
of turnover of the SPX4-RFP fusion in P-limited displayed lower shoot biomass under optimal growth
Plant Physiol. Vol. 181, 2019 337
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Osorio et al.
conditions (Fig. 4B, P-replete), asserting that the main- accumulated more Pi than Col-0 irrespective of Pi supply
tenance of Pi homeostasis is linked to plant fitness and to similar levels as pho2-1 (Fig. 4D). The weaker
(Rouached et al., 2011; Linn et al., 2017). Mutants were knockdown allele, spx4-2, showed this effect only in
not impaired in root growth (Fig. 4A). P-replete seedlings. Since the T-DNA insertion in this line is
In our system, phr1 and pho2-1 mutants displayed the in the C-terminal portion of the functional domain, just 59 of
expected trends in organ Pi accumulation. Both phr1-1 and the coding region for the active Lys-binding cluster but after
phr1-2 showed faster depletion of their shoot Pi pools under the phosphate-binding cluster residues (Supplemental Fig.
Pi-limiting conditions (Fig. 4D). This is in line with the S7A; Wild et al., 2016), a partially functional SPX domain
positive role that PHR1 exerts over the regulation of Pi might explain the weaker response. The observed changes
uptake and translocation (Nilsson et al., 2007). On the other in Pi allocation place SPX4 as a central negative regulator of
hand, pho2-1 displayed the typical shoot Pi over- Pi acquisition, alongside PHO2.
accumulation phenotype irrespective of Pi supply (Fig. 4D)
as well as reduced Pi concentration in P-replete roots
(Fig. 4C). This is due to the negative effect of PHO2/UBC24 SPX4 Is a Negative Regulator of PSR Gene Expression in
on xylem Pi loading in roots via suppression of PHO1 (Liu Shoots, Not Roots
et al., 2012). By contrast, disrupting SPX4 led to higher in-
tracellular Pi accumulation in both P-replete roots and Knocking out the negative regulators SPX1 and SPX2
shoots. In shoots, the true knockout allele, spx4-1, led to derepression of PHR1 in P-replete as well as
Figure 3. SPX4 affects the translocation of PHR1 to the nucleus in a dose- and Pi-dependent manner. Transfecting spx4-1 mutant
protoplasts with equal amounts of 35Spro:SPX4-RFP and 35Spro:PHR1-GFP constructs leads to partial retention of PHR1-GFP in
the cytosol, but only in P-replete conditions (1P). In P-limited (2P) spx4-1 protoplasts, PHR1-GFP fluorescence is almost ex-
clusively found in the nucleus. Increasing the ratio of SPX4-RFP over PHR1-GFP fusion construct in the transfection reaction (from
0.25-fold in the top row of images to 4-fold in the bottom row of images) leads to more prominent eGFP fluorescence in the
cytosol. At the same time, RFP fluorescence intensity increases from the top to bottom row of images, but the signal remains
evenly distributed between nucleus and cytosol. Almost exclusive nuclear localization of the PHR1-GFP fusion protein in
P-limited protoplasts is disrupted only in the presence of a 4-fold excess of SPX4-RFP protein (bottom row of images). On the right,
the number of transfected protoplasts over the total number of cells scanned for fluorescence is given in parentheses. The images
shown are representative of each transfection reaction. Asterisks indicate that between one (1:1 ratio, 2P) and five (4:1 ratio, 1P)
protoplasts had detectable levels of RFP fluorescence only, indicating reduced cotransfection rates with PHR1-GFP. Chl, Chlo-
rophyll. Bars 5 5 mm.
338 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
AtSPX4 Regulates Intracellular P Status in Shoots
Figure 4. Disruption of SPX4 function
leads to Pi overaccumulation in shoots.
Tissue biomass (A and B) and phosphate
concentration (C and D) are shown for
Arabidopsis roots and shoots grown un-
der varying Pi supplies. Plants defective
in two key regulators of P signaling with
contrasting Pi accumulation profiles
(PHR1 and PHO2) were included for
comparison. Values are means 6 SE of at
least three independent biological rep-
licates, comprising pools of four to six
shoots and eight to 12 roots each. As-
terisks indicate significant differences
from the wild type under each condition
(P , 0.05, ANOVA/Fisher LSD). FW,
Fresh weight.
Pi-resupplied seedlings (Puga et al., 2014; Fig. 5; Set S2B) may account for their higher P i accumula-
Supplemental Table S1). This is in contrast to PSR genes tion (Fig. 4D). It is important to note that there is
in the wild type that are active upon Pi withdrawal. very little overlap (less than 5%) between SPX1/2-
The shoot Pi overaccumulation phenotype of the spx4 and SPX4-controlled differentially expressed genes
mutants prompted the hypothesis that SPX4 knock- (DEGs) in P-replete seedlings, indicating that SPX1/2
out would result in a similar misregulation of PSR control different subsets of PSR genes compared with
genes, both Pi-starvation induced (PSI) and Pi-s- SPX4. SPX4 knockout had very little effect on transcript
tarvation suppressed (PSS), in P-replete plants. Using profiles in roots (Fig. 5; Supplemental Data Set S1).
an RNA sequencing (RNA-seq) approach, the root and Among the 50 genes with more than 2-fold lower ex-
shoot transcriptome profiles from the stronger spx4- pression (FDR , 0.05) in P-replete spx4-1 versus Col-0
1 allele and the two phr1 alleles were compared with shoots, only 6% were PSS and only 22% were PHR1-
those of Col-0 seedlings across all three conditions dependent genes (Supplemental Table S1). SPX4 is one
(P replete, P limited, and Pi resupply; Supplemental of these genes, confirming gene knockout in spx4-1.
Data Sets S1 and S2). The two phr1 alleles showed In contrast to whole seedlings of the spx1 spx2
the expected misregulated PSR gene expression in double mutant (Puga et al., 2014), P i resupply to
P-limited roots and shoots and wild-type-like ex- P-limited spx4-1 seedlings did not lead to significant
pression profiles in P-replete and Pi-resupplied deregulation of PSR genes (Fig. 5; Supplemental
seedlings (Fig. 5; Supplemental Table S1). In P-replete Table S1). It is possible that the missing effect of
spx4-1 shoots, 166 genes were differentially expressed SPX4 knockout on PHR1-dependent genes could
compared with the wild type (Fig. 5; Supplemental be masked by genetic redundancy with SPX1
Table S1; Supplemental Data Set S2, B and C). and SPX2.
Seventy-two percent of differentially induced genes In summary, these expression profiles suggest
in P-replete spx4-1 shoots were PSI genes (i.e. re- that SPX4 is a negative regulator of a subset of
sponsive to changes in Pi supply in Col-0), but only PSR genes in shoots that are not all controlled by
16% of these were affected by PHR1 knockout PHR1 but possibly by another, yet-to-be-identified
(Supplemental Table S1). The induction of PHO1 transcription factor. Candidates could potentially
found in P-replete spx4-1 shoots (Supplemental Data be found in differentially expressed transcription
Plant Physiol. Vol. 181, 2019 339
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Osorio et al.
genes suppressed (68%) and 125 induced (32%;
Supplemental Data Set S3B). Within 1 d of Pi resupply
to roots, the expression of 49% of PSI and 27% of PSS
genes was back to control levels. Overall, similar trends
were observed in Col-4 (Fig. 6).
Compared with the wild type, all mutants failed to
fully respond to the changes in Pi availability
(Supplemental Data Sets S3, C–E, and S4, C–E). As ex-
pected, lines disrupted in PHR1 were the most affected
in all conditions. phr1-1 was reported to affect about
47% and 69% of P-responsive genes by at least twofold
in P-starved roots and shoots, respectively (Bustos et al.,
2010). Using the same selection criteria, about 27% of
root and 46% of shoot PSR genes showed a reduced
response in both phr1 mutants following 5 d of Pi
withdrawal (Fig. 6A, blue bars). The effect of PHR1
disruption was also evident on Pi resupply, with 23% of
root PSR genes and 26% of shoot PSR genes not
responding to the same extent as in Col-0 (Fig. 6B, blue
bars). SPX4 or PHO2 knockout also had a strong effect
on the PSR response in shoots, with 42% and 40% of
Figure 5. Knockout of SPX4 leads to ectopic PSR gene expression in
PSR genes misregulated in spx4-1 and pho2-1, respec-
P-replete shoots but not in roots. Knockout of SPX genes should lead to
the release of PHR1-activated gene expression under P-replete and
tively. The weaker spx4-2 allele still affected 29% of
Pi-resupply conditions, as shown here for DEGs in whole seedlings of shoot PSR genes (Fig. 6). In contrast to phr1 mutants, the
the spx1 spx2 double mutant (asterisk; Puga et al., 2014). PHR1 response to Pi resupply in shoots (or roots) was not
knockout primarily affects PSR gene expression in P-limited organs, significantly altered, with less than 13% of PSR genes
shown here as the common response to changes in Pi supply of both misregulated in spx4-1 and pho2-1 compared with Col-
phr1-1 and phr1-2 alleles (this work; greater than 2-fold change in 0, respectively (Fig. 6B). Knockout of these two genes
mutant versus the wild type; false discovery rate [FDR] , 0.05). The also had very little effect on the Pi-responsive tran-
spx4-1 mutant shows the expected response in P-replete shoots, but not scriptome in roots, with less than 14% of PSR genes
under Pi resupply. spx4-1 mutant roots show a similar transcript profile misregulated in the stronger spx4-1 allele and pho2-
to the wild type. See Supplemental Data Sets S1 and S2 for details.
1 compared with Col-0 (Fig. 6A; Supplemental Data
Set S3). Together, the expression profiles shown in
factors highlighted in Supplemental Data Set S2, B Figures 5 and 6 suggest that SPX4 acts as a negative
and C. regulator of a subset of shoot PSR genes in P-replete
shoots, while also being required for the expression of
more than 40% of PSR genes in P-limited shoots.
SPX4 Knockout Disrupts PSR Gene Expression in
P-Limited Shoots
SPX4 Predominantly Acts as a PHO2- and PHR1-
Given the relatively small number of DEGs in Dependent Modulator of PSR Gene Expression in Shoots
P-replete spx4-1 shoots compared with the wild type,
the magnitude of the transcriptome response to The finding that, in contrast to SPX1/2, SPX4 has a
changes in Pi supply was analyzed in each genotype significant impact on the orderly launch of the tran-
using two comparisons (Fig. 6; Supplemental Data Sets scriptomic PSR in P-limited shoots poses the question of
S3 and S4). In the first comparison, differences between whether this response is PHR1 dependent. In addition
P-limited and P-replete organs were assessed in each to the PSR genes with reduced responsiveness in the
genotype (Fig. 6A). The second comparison identified mutants described above, genes that were responsive in
expression changes in P-limited versus Pi-resupplied the mutants only (ancillary genes in Fig. 6 [gray bars];
organs (Fig. 6B). In P-limited shoots of Col-0, 1,148 log2 . 2, FDR , 0.05) were also identified, as these
genes (71%) were induced (PSI genes) and 469 genes show an additional response to Pi withdrawal or
(29%) were suppressed (PSS genes) compared with resupply not observed in the wild type. Together, these
P-replete conditions. Relative to P-limited Col-0 shoots, genes were termed genes misregulated in mutants
Pi resupply to this organ led to suppression of 857 genes (GMMs). Given similar patterns of perturbation of
(56%) and induction of 663 genes (44%), respectively. GMM expression in spx4-1 and pho2-1 shoots, the extent
Within 24 h of Pi resupply, the expression of 49% of PSI of overlap in GMMs between these two mutants, as
and 10% of PSS genes was back to control levels well as between spx4-1 and phr1, was investigated
(Supplemental Data Set S4B). Roots were less respon- further. Of the 545 GMMs induced in P-limited spx4-
sive to Pi withdrawal, with 459 genes induced (59%) 1 shoots, 63% and 45% were similarly affected in pho2-
and 313 suppressed (41%), and to Pi resupply, with 264 1 and phr1, respectively, and 35% were misregulated in
340 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
AtSPX4 Regulates Intracellular P Status in Shoots
Figure 6. Knockout of SPX4 specifically modulates the shoot Pi starvation response. A, Proportion of genes with altered ex-
pression in roots or shoots with changes in Pi supply across genotypes. The two phr1 alleles show 27% and 46% perturbation of
PSR gene expression in roots and shoots, respectively. The two spx4 alleles and pho2-1 show a similar disturbance of PSR gene
expression in P-limited shoots but not in roots. B, Proportion of genes with an altered response to Pi resupply in P-limited roots and
shoots across genotypes. phr1 mutants are impaired in their response to Pi resupply, with 23% and 26% of PSR genes misregulated
in roots and shoots, respectively. The spx4 and pho2-1 mutants have an almost wild-type-like response to Pi resupply. Shown are
PSR DEGs with a more than twofold altered expression ratio in each treatment comparison (red bars; FDR , 0.05). Misregulated
genes (blue bars) represent PSR genes with altered (greater than twofold expression difference) response in each mutant compared
with the wild type. Ancillary genes (gray bars) show a greater than fourfold change in expression ratio in individual mutants
(FDR , 0.05) but not in the wild type. See Supplemental Data Sets S3 and S4 for details.
all three mutants (Fig. 7B; Supplemental Data Set S4). Many of the downstream targets misregulated in
Gene Ontology (GO) term enrichment analysis of the 342 P-limited shoots of all three mutants are transcription
GMMs shared by both spx4-1 and pho2-1 revealed that factors, hinting at a more complex regulatory network.
genes associated with classical acclimation responses to To investigate whether specific transcription factor
Pi starvation, such as amino acid, ion, and lipid transport families were overrepresented among the GMMs, an in-
as well as flavonoid biosynthesis and jasmonic acid depth enrichment analysis was performed (Fig. 9A).
signaling, were overrepresented (Fig. 7A). A large Analysis of individual transcription factor families
number of transcription factors were among these genes. revealed that plant-specific growth-regulating factors
Many are associated with the regulation of the afore- (GRFs) were misregulated in all three genotypes, as
mentioned acclimation responses, while others are were REPRODUCTIVE MERISTEM (part of the B3 su-
regulators of organ development and meristem main- perfamily of transcription factors), trihelix (GT-element
tenance. About half the genes associated with these binding), and NIN-LIKE PROTEIN transcription
processes are PHR1 dependent. PHO2 was identified as factors.
an upstream regulator of 53% of the 211 GMMs sup- Hierarchical clustering of the expression ratios (log2
pressed by Pi withdrawal in spx4-1 shoots (Fig. 8B; fold difference in transcript abundance in P-limited
Supplemental Data Set S4D). PHR1 targets 39% of these versus P-replete mutant shoots) of misregulated tran-
SPX4-dependent GMMs; 34% of the PSS GMMs are scription factors in spx4-1 revealed that the majority of
shared between all three mutants. GMMs shared by transcription factors were PSI genes that failed to be
spx4-1 and pho2-1 mutants show enrichment of GO terms induced in P-limited mutant shoots (Fig. 9B;
associated with defense-related processes, and most of Supplemental Data Set S5A), acting downstream of
these genes are dependent on PHR1 (Fig. 8A). SPX4 (and PHO2; class 3b) or of SPX4 (PHO2) and
Plant Physiol. Vol. 181, 2019 341
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Osorio et al.
Figure 7. GMMs induced in P-limited
spx4-1 and pho2-1 shoots are associated
with classical acclimation responses as
well as plant development. A, GO terms
enriched in the 342 GMMs induced by Pi
withdrawal in both spx4-1 (514 mis-
regulated 1 31 ancillary) and pho2-
1 (414 misregulated 1 111 ancillary;
63% overlap in B) mutants are associated
with classical acclimation processes such
as ion, amino acid, and lipid transport,
flavonoid biosynthesis, and jasmonic acid
response. There is a strong association
with transcriptional regulation and devel-
opment. Shown in dark red are genes
differentially expressed in both phr1 al-
leles. B, Overlap of GMMs in P-limited
versus P-replete shoots of spx4-1, pho2-
1, and phr1. Misregulated and ancillary
GMMs for each mutant are listed in
Supplemental Data Set S4.
PHR1 (class 2). Among this group of genes, many were contained a number of candidates for direct down-
found to be ectopically induced in P-replete spx4- stream targets of SPX4: ANAC044 is an ancillary, up-
1 shoots (marked by asterisks in Fig. 9B), with ectopic regulated gene in P-replete spx4-1 and pho2-1 shoots as
meaning that these genes are PSR genes that would well as an ancillary PSI gene in P-limited phr1 shoots
normally respond to low P status (Supplemental Data (Fig. 9B; Supplemental Fig. S8). This makes ANAC044 a
Set S4B) but are already induced in P-replete spx4- target gene with the expected contrasting expression
1 shoots (Supplemental Data Set S2B). This particular between SPX4 and PHR1 knockout plants, as suggested
profile suggests that they are part of a PSR network in the rice model (Lv et al., 2014). ANAC055 is an an-
controlled by SPX4 via negative regulation of a tran- cillary PSI gene in P-limited spx4-1 and pho2-1 shoots
scription factor, such as PHR1 (class 2) or an unknown (Fig. 9B; Supplemental Fig. S8). AGL25/FLC showed
factor (class 3b). SPX4 also seems to act as a positive the opposite response and failed to be induced in
regulator of four transcription factors, AGAMOUS- P-limited spx4-1 shoots compared with the wild type
LIKE25 (AGL25)/FLOWERING LOCUS C (FLC), (Fig. 9B; Supplemental Data Set S2C; Supplemental Fig.
GRF5, ARABIDOPSIS NAC DOMAIN CONTAINING S8). Both ANAC055 and AGL25/FLC expression were
PROTEIN31 (ANAC031), and EMBRYO SAC DEVEL- wild type like in phr1-2 shoots. SPX4 (and PHO2) thus
OPMENT ARREST1, in P-limited shoots (marked by appear to modulate the expression of both transcription
hash marks in Fig. 9B; Supplemental Data Set S2C). A factors in P-limited shoots in a PHR1-independent
smaller group of transcription factors showed an an- manner. GMM transporters showed similar expres-
cillary response to changes in P status in spx4-1 (and the sion profiles (Fig. 9C; Supplemental Data Set 5B). This
other two mutants; class 3a in Fig. 9B). This class could indicate that transporters are prominent
342 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.AtSPX4 Regulates Intracellular P Status in Shoots
Figure 8. GMMs suppressed in P-limited spx4-1 and pho2-1 shoots are associated with plant defense. A, GO terms enriched in
the 116 GMMs suppressed by Pi withdrawal and shared by spx4-1 (161 misregulated 1 50 ancillary) and pho2-1 (227 mis-
regulated 1 12 ancillary; 53% overlap in B) are associated with defense-related processes such as programmed cell death and
salicylic acid response. Most of these genes are also GMMs in both phr1 alleles (dark red). B, Overlap of GMMs in P-limited versus
P-replete shoots of spx4-1, pho2-1, and phr1. Misregulated and ancillary GMMs for each mutant are listed in Supplemental Data
Set S4.
downstream targets of the transcription factors men- and cross-compared these with coexpression profiles
tioned above. The largest group was again formed by and experimentally confirmed protein-DNA interac-
PSI genes misregulated in all three mutants (Fig. 9C, tions. The network was further extended by adding
class 2) or in spx4-1 (and pho2-1; class 3b). Class 3b experimental evidence for protein-protein interactions
contained many ancillary GMMs that were ectopically between these regulators and downstream targets. This
expressed in P-replete spx4-1 shoots compared with resulted in 719 nodes and 3,900 edges that were visu-
Col-0 (marked by asterisks in Fig. 9C; Supplemental alized in Cytoscape (Fig. 10). Data from pho2-1 and phr1
Data Set S2B). Notably, these comprised not only PHO1 GRN analyses were subsequently used to further an-
but also iron transporter genes FERRIC REDUCTASE notate and color code this list (Fig. 10; Supplemental
DEFECTIVE3 and VACUOLAR IRON TRANS- Data Set S6).
PORTER-LIKE5, low-affinity nitrate transporter NI- Notably, eight of the identified upstream regulators
TRATE TRANSPORTER1.9, and four USUALLY predicted to bind to the promoters of GMMs were
MULTIPLE ACIDS MOVE IN AND OUT TRANS- transcription factors misregulated in spx4-1 shoots
PORTERS (UMAMIT) transporters (Supplemental (Supplemental Data Set S6, highlighted in yellow). Five
Fig. S8). of these, ANAC028, ANAC031, ANAC046, ANAC055,
Genes with asterisks or hash marks in Figure 9 hint at and WRKY75, were also identified as regulators of
separate functions of SPX4 as a negative regulator (as- pho2-1 and phr1 GMMs (Supplemental Data Set S6,
terisks) of PSI genes in P-replete shoots or a positive column B, highlighted in purple).
regulator (hash marks) in P-limited shoots. However, PSI and PSS target genes misregulated in spx4-
the majority of GMMs appear to be targets of both SPX4 1 shoots formed two subnetworks that were under
activities, showing a slight derepression in P-replete the immediate control of distinct transcription factor
shoots and a lack of full induction in P-limited shoots. clusters (Fig. 10): PSS targets were predominantly reg-
This is possibly the reason why absolute differences ulated by NAC and WRKY transcription factors. Of
from treated Col-0 shoots are small (Fig. 5) while the those, WRKY35 and a known regulator of PSR gene
effect on log2 fold changes in P-limited over P-replete expression, WRKY75 (Devaiah et al., 2007), were
shoots is significant (Fig. 6A). themselves differentially expressed in shoots of spx4-
1 and pho2-1 (WRKY35; class 3b in Fig. 9B) or all three
mutants (WRKY75; Supplemental Data Set S2C).
Gene Regulatory Network Analysis Reveals Central ANAC044-binding sites were also enriched in pro-
Upstream Regulators of PSR Affected by SPX4 moters of PSS-GMMs. Due to its differential expression
profile mentioned above, ANAC044 is a candidate for a
Given the high number and complex Pi-dependent direct target of both SPX4 and PHR1 (Fig. 9B;
expression profiles of transcription factors mis- Supplemental Fig. S8). PSI targets misregulated in spx4-
regulated in spx4-1 shoots, we conducted a gene regu- 1 shoots, on the other hand, were controlled by a more
latory network (GRN) analysis using the TF2Network complex regulatory network comprising HMGBD15,
tool to identify upstream regulators (Kulkarni et al., SPLs, RGLs, ANAC055, AGL20/SOC1, AGL25/FLC,
2018). The tool searched the corresponding GMM pro- and PHR1-related MYBs such as GCC7 and PHL1
moters for enriched transcription factor-binding sites (Bustos et al., 2010; Lundmark et al., 2011; Fig. 10).
Plant Physiol. Vol. 181, 2019 343
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Osorio et al.
Figure 9. Transcription factors and
transporters show significantly altered
expression profiles in spx4-1, pho2-1,
and phr1 mutant shoots. A, Heat map of
transporters and individual transcription
factor (TF) families enriched among
spx4-1 GMMs (misregulated and ancil-
lary genes in Supplemental Data Set S4).
B and C, Hierarchical clusters of ex-
pression ratios of individual transcrip-
tion factor (B) or transporter (C)
transcripts across genotypes that fail to
be correctly induced (red) or suppressed
(blue) in P-limited versus P-replete spx4-
1 shoots compared with the wild type
(misregulated PSR-GMMs) or that are
ectopically expressed in spx4-1 only
(ancillary GMMs). Genes differentially
expressed in spx4-1 versus wild-type
shoots (Supplemental Data Set S2) are
marked by asterisks (P replete) or hash
marks (P limited). D, Genes were
assigned to classes according to their
expression profiles depicted in stylized
bar charts (shown for PSI genes only).
Class 1 5 SPX1/2, which act as negative
regulators of PHR1. Class 2 5 altered
expression of PSR genes in spx4-1,
(pho2-1), and phr1. SPX4 acts as a neg-
ative (*) or positive (#) modulator of
PHR1. Class 3a 5 ectopic expression in
spx4-1 but not in Col-0. SPX4 acts as a
repressor of target genes in P-limited
shoots. Class 3b 5 genes with altered
PSR gene expression in spx4-1 (and
pho2-1). SPX4 acts as a PHR1-
independent negative (*) or positive (#)
modulator of PSR gene expression. Class
4 (not shown) 5 ectopic expression in
all three mutants but not in Col-0.
AGL20 was identified as an upstream regulator The ANAC055 transcription factor is predicted to target
of ANAC055 and AGL25, with AGL20 experimen- the promoters of PSI-GMMs in all three mutants and
tally confirmed to bind to the ANAC055 promoter PSS-GMMs in pho2-1 and phr1 (Supplemental Data Set
(Supplemental Data Set S6). AGL20 transcripts were S6). The latter group shares 80% common targets, so
suppressed in P-limited over P-replete wild-type and ANAC055 and PHR1 may converge on their promoters
phr1 shoots while being expressed in both spx4-1 and to suppress them. In the former group, there is very
pho2-1 shoots, irrespective of P status (Fig. 9B). This little overlap (18% of ANAC055 targets among spx4-
would suggest that SPX4 and PHO2 are negative reg- 1 and phr1 PSI-GMMs), hinting at competition be-
ulators of AGL20 in P-limited shoots. As mentioned tween the two transcription factors for common targets.
above, transcripts of ANAC055 were highly abundant PSI and PSS gene targets appear to be further connected
in P-limited spx4-1 and pho2-1 shoots, while AGL25 through their regulation by another group of
transcripts showed the opposite response (Fig. 9B; NAC transcription factors. Three of those, ANAC028,
Supplemental Fig. S8). AGL20 would thus activate ANAC031, and ANAC046, were differentially expressed
ANAC055 and repress AGL25 expression. The con- in spx4-1 shoots compared with the wild type (Fig. 9B;
trasting expression profiles of the latter would fur- Supplemental Data Set S2). ANAC031 is a predicted
thermore support the GRN analysis prediction that downstream target of AGL20, which would therefore be
ANAC055 also binds to the AGL25 promoter, acting as predicted to act as a transcriptional repressor of
a repressor (Supplemental Data Set S6). The expression ANAC031. ANAC028 and ANAC046 target promoters of
of all three transcription factors is PHR1 independent. Pi-responsive GMMs in all three mutants (Supplemental
344 Plant Physiol. Vol. 181, 2019
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.AtSPX4 Regulates Intracellular P Status in Shoots
Figure 10. Gene regulatory networks of SPX4-dependent transcriptional responses. The TF2Network tool (Kulkarni et al., 2018)
identified ANAC055 as a potential upstream regulator of PSR in all three mutants (Supplemental Data Set S6). It is itself a target of
HMGBD15, RGL2/3, and SPL2/10/11 (predicted protein-DNA interaction). ANAC055 transcript abundance is high in P-limited
spx4-1 and pho2-1 shoots, with no Pi response in the wild type (WT). AGL20/SOC1 binds to the ANAC055 promoter (confirmed
protein-DNA interaction), and AGL20 and AGL25/FLC are both connected to ANAC055 through protein-protein interactions.
ANAC028, ANAC031, and ANAC046 are candidates for SPX4-, PHO2-, and PHR1-dependent regulators, as they target pro-
moters of PSI and PSS GMMs in all three mutants. Predicted binding of transcription factors (TFs) to target gene promoters is
indicated with light gray lines. Confirmed transcription factor-DNA interactions are depicted in dark gray. Protein-protein in-
teractions are shown as black lines. Upstream regulators are highlighted as larger nodes. Protein-protein interactions between
transcription factors in the pho2-1 network are shown in gold. Asterisks mark transcription factors coexpressed in spx4-1 and
pho2-1. Hash marks highlight transcription factors with an altered PSR in all three mutants. For better visibility, ANAC was
shortened to NAC.
Data Set S6), which would make them common down- overlapping with PHR1 targets. If this were the case,
stream targets of SPX4, PHO2, and PHR1. these MYB transcription factors would form a smaller
PHR1 could not be placed within this network, as no subnetwork controlling GMMs induced upon Pi with-
DNA or protein interaction data were available in the drawal in all three mutants (Fig. 10), consistent with our
TF2Network database. One can assume that target finding that only about one-third of SPX4-dependent
genes predicted for PHL1 and GCC7 are partially genes are controlled by PHR1.
Plant Physiol. Vol. 181, 2019 345
Downloaded on February 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































