Role of Pneumococcal NanC in the Severe Disease of Streptococcus pneumoniae
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Role of Pneumococcal NanC in the Severe Disease of
Streptococcus pneumoniae Superinfection with Influenza
Avijit Dutta, Yi-Yin Chen, Tse-Ching Chen, Chia-Shiang Chang, Yu-Lin Huang, Ting-An Chen,
Yung-Chang Lin, Chun-Yen Lin, Yu-Chia Hsieh and Ching-Tai Huang
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
ImmunoHorizons 2021, 5 (4) 210-218
doi: https://doi.org/10.4049/immunohorizons.2100020
http://www.immunohorizons.org/content/5/4/210
This information is current as of January 14, 2022.
References This article cites 37 articles, 12 of which you can access for free at:
http://www.immunohorizons.org/content/5/4/210.full#ref-list-1
Email Alerts Receive free email-alerts when new articles cite this article. Sign up at:
http://www.immunohorizons.org/alerts
ImmunoHorizons is an open access journal published by
The American Association of Immunologists, Inc.,
1451 Rockville Pike, Suite 650, Rockville, MD 20852
All rights reserved.
ISSN 2573-7732.RESEARCH ARTICLE
Adaptive Immunity
Role of Pneumococcal NanC in the Severe Disease of
Streptococcus pneumoniae Superinfection with Influenza
Avijit Dutta,*,† Yi-Yin Chen,‡ Tse-Ching Chen,§,{ Chia-Shiang Chang,† Yu-Lin Huang,† Ting-An Chen,†
Yung-Chang Lin,||,# Chun-Yen Lin,**,†† Yu-Chia Hsieh,‡,‡‡,1 and Ching-Tai Huang*,†,§§,1
*Research Center for Emerging Viral Infections, College of Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan; †Division of Infectious Diseases,
Department of Medicine, Chang Gung Memorial Hospital, Guishan, Taoyuan City, Taiwan; ‡Division of Infectious Diseases, Department of Pediatrics, Chang
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
Gung Children's Hospital, Chang Gung Memorial Hospital, Guishan, Taoyuan City, Taiwan; §Department of Pathology, Chang Gung Memorial Hospital, Guishan,
Taoyuan City, Taiwan; {Department of Pathology, College of Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan; ||Division of Hematology and
Oncology, Department of Medicine, Chang Gung Memorial Hospital, Guishan, Taoyuan City, Taiwan; #Division of Hematology and Oncology, College of
Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan; **Division of Hepatogastroenterology, Department of Medicine, Chang Gung Memorial
Hospital, Guishan, Taoyuan City, Taiwan; ††Division of Hepatogastroenterology, College of Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan;
‡‡
Division of Infectious Diseases, Department of Pediatrics, College of Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan; and §§Division of
Infectious Diseases, College of Medicine, Chang Gung University, Guishan, Taoyuan City, Taiwan
ABSTRACT
Bacterial superinfection aggravates the disease of influenza. Streptococcus pneumoniae is the most common bacterial pathogen.
Synergistic virulence has been demonstrated between influenza neuraminidase and pneumococcal NanA and NanB. NanC, the other
pneumococcal neuraminidase infrequently present in clinical isolates, is not well characterized. In this study, we report that
superinfection with a NanC-negative pneumococcus strain suppresses anti-influenza immunity and impairs viral clearance with higher
TGF-b activation in mice. Bacterial load in the lungs also increases as the host immunity is suppressed. NanC-positive isogenic mutant
reverses wild type S. pneumoniae--mediated immune suppression and facilitates virus clearance. However, it causes more severe
disease as the augmented inflammation causes collateral damage. Both virus-mediated damage and immune response--mediated
inflammation are important for pathogenesis of severe influenza. Inflammation may be more critical than virus-mediated damage in
influenza with bacterial superinfection. ImmunoHorizons, 2021, 5: 210–218.
Influenza virus infection is a substantial burden on public virus and S. pneumoniae. There are three pneumococcal NAs:
health worldwide. Bacterial superinfection aggravates influenza NanA, NanB, and NanC. Synergistic virulence has been shown
with increased morbidity and mortality. Streptococcus pneumo- between NanA and influenza NA. NanB may compensate for
niae is the most common pathogen for superinfection (1--6). the effect of NanA (7). NanC is the least characterized as it is
Neuraminidase (NA) is a virulence factor for both influenza not present in the majority of clinical isolates of S. pneumoniae.
Received for publication March 2, 2021. Accepted for publication March 5, 2021.
Address correspondence and reprint requests to: Dr. Yu-Chia Hsieh or Dr. Ching-Tai Huang, Division of Infectious Diseases, Department of Pediatrics, Chang Gung
Children’s Hospital, Chang Gung Memorial Hospital, Chang Gung University, 5 Fu-Hsin Street, Kwei-Shen, Taoyuan City, Taiwan 33333 (Y.-C.H.) or Division of
Infectious Diseases, Department of Medicine, Chang Gung Memorial Hospital, Chang Gung University, 5 Fu-Hsin Street, Kwei-Shen, Taoyuan City, Taiwan 33333 (C.-
T.H.). E-mail addresses: yuchiahsieh@gmail.com (Y.-C.H.) or chingtaihuang@gmail.com (C.-T.H.)
ORCID: 0000-0002-2084-5348 (Y.-C.L.).
1
Y.-C.H. and C.-T.H. contributed equally to this work.
This work was supported by the Research Center for Emerging Viral Infections from The Featured Areas Research Center Program within the framework of the Higher
Education Sprout Project by the Ministry of Education of Taiwan; Ministry of Science and Technology of Taiwan projects MOST-109-2634-F-182-001, MOST-109-
2321-B-182A-003-MY3 (A.D.), MOST-106-2314-B-182A-160-MY3, and MOST-109-2314-B-182A-150-MY3 (C.-T.H.) and the Special Researcher Recruitment
Program (MOST-105-2811-B-182A-006; MOST-106-2811-B-182A-011; MOST-108-2811-B-182A-517; and MOST-109-2811-B-182A-509) (A.D.); and by Medical
Research Project Fund of Chang Gung Memorial Hospital of Taiwan CMRPG3J0721-3, CMRPG3J1501-2, and CMRPG3J1511-2 (C.-T.H.).
Abbreviations used in this article: DANA, 2-deoxy-2,3-dihydro-N-acetylneuraminic acid; F, forward; HA, hemagglutinin; NA, neuraminidase; PR8, A/PR/8/34; R,
reverse; spec, spectinomycin.
This article is distributed under the terms of the CC BY-NC 4.0 Unported license.
Copyright © 2021 The Authors
210 https://doi.org/10.4049/immunohorizons.2100020
ImmunoHorizons is published by The American Association of Immunologists, Inc.ImmunoHorizons ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA 211
It is present in one third to half of the tested strains (8--11). Transgenic mice
NanC, just like NanA and NanB as well as influenza NA, The TCR transgenic mouse line 6.5, which expresses a TCR
cleaves a2,3-linked sialic acids to produce N-acetylneuraminic recognizing an I-Ed--restricted HA epitope (110SFERFEIFPKE120;
acids (9, 12--14). 2-Deoxy-2,3-dihydro-N-acetylneuraminic acid generously provided by H. Von Boehmer, Harvard University,
(DANA) is the N-acetylneuraminic acid NanC produces. As Boston, MA) were BALB/c genetic background. We used
DANA inhibits NanA and NanB enzymatic activity, it is inter- mAb against I-Ed--restricted HA-specific TCR (anti-6.5) as a
esting to learn the role of NanC in influenza with pneumococ- marker for 6.5 CD41 T cells. In some experiments, we used
cal superinfection (10, 15, 16). Thy-1.1 as a surrogate marker for 6.5 TCR if Thy-1.11 6.5
The clinical outcome relies on the timing of the two infec- CD41 T cells were adoptively transferred into Thy-1.2/1.2
tions in the mouse experimental model. Influenza virus infec- recipients (17--20). Wild type BALB/c mice were purchased
tion with S. pneumoniae superinfection, both in nonlethal doses, from the National Animal Laboratory Center (Taipei City,
on day 3 to day 7 causes 100% lethality. Influenza virus and S. Taiwan). All mice were maintained in a specific pathogen-
pneumoniae coinfection on the same day causes 50% mortality, free environment and were used for experiments between
and influenza virus infection with a prior exposure of S. pneu- the ages of 8--24 wk.
moniae 7 d earlier causes no lethality (2). As the lethal syner-
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
gism requires an influenza virus infection before the challenge Infection and monitoring
with S. pneumoniae (2, 6), influenza virus infection--mediated The A/PR/8/34 (PR8) strain of influenza virus was produced
alterations of host respiratory epithelium and host immunity in the allantoic fluid of 10-d-old embryonated chicken eggs and
were the focus of research. However, bacterial factors of the S. characterized by a core facility at the Chang Gung University.
pneumoniae may play a role as well through the interactions Mice were inoculated intranasally during light isoflurane anes-
thesia with stated doses of virus in 50 ml PBS (17--20). A clini-
with influenza virus and host immune response during the
cal isolate of S. pneumoniae serotype 3 (Taian strain, 21) was
superinfection.
grown in Todd--Hewitt broth supplemented with 0.5% yeast
In this study, we study the role of pneumococcal NanC on
extract to midlogarithmic phase (OD at 600 nm = 0.2--0.5). The
anti-influenza immunity during the superinfection through the
bacterial cultures were then diluted (using PBS) to the desired
use of our influenza hemagglutinin (HA) Ag-specific transgenic
cell density (CFU/ml) using a previously determined standard
mouse model. In our model, influenza virus in the lungs reach
curve (OD versus actual CFU) generated for pneumococci. The
its peak by days 2--4, and no plaque forming viable virus can be
nominal sizes of the inoculating doses were confirmed by viable
detected by day 14 if mice survive the infection (17--20). We do
counts after plating on blood agar plates. We inserted pneumo-
adoptive transfer of naive influenza HA-specific CD41 T
coccal nanC gene into the Taian bacterial strain as described
cells at the time of influenza virus infection to study the
below and cultured the nanC-inserted bacteria similarly. Mice
Ag-specific T cell response to influenza virus infection received superinfection through intranasal inoculation of bacte-
in vivo. All the HA-specific CD41 T cells have all the ria in 50 ml PBS on day 4 post--influenza virus infection during
TCRs responsive to influenza HA only (17--20). They re- light isoflurane anesthesia. Infected mice were monitored every
spond to the infection with a Th1 phenotype and help in day for disease manifestation. Mice with severe sickness were
virus clearance but also contribute to lung inflammation euthanized humanely in accordance with the guidelines provid-
(17, 19). We use this model to study influenza with S. pneu- ed by the institutional ethical committee.
moniae superinfection and the superinfection-mediated al-
teration of anti-influenza immunity. Virus titer determination
We measured live virus titer in organs of influenza virus--
infected mice using a modified Madin Darby canine kidney cell
MATERIALS AND METHODS (American Type Culture Collection) plaque assay (17--20). The
organs were collected at the indicated times in 1 ml DMEM,
Ethics statement snap frozen in liquid nitrogen, and stored at 80 C until ana-
All experiments involving the use of mice were performed in lyzed. The 10-fold dilutions of the tissue homogenates were
accordance with protocols approved by the Animal Care and prepared in DMEM supplemented with 10% FCS, antibio-
Use Committee of the Chang Gung Memorial Hospital of Tai- tic--antimycotic (15240-062, Life Technologies BRL), and
wan (approval no. 2015092102). The committee recognizes that 0.00025% trypsin. A total of 500 ml of each dilution was
all the experiments involving the use of mice were carried out added to confluent monolayers of Madin Darby canine kid-
in strict accordance with the Animal Protection Law by the ney cells in six-well plates and incubated for 1 h at 35 C
Council of Agriculture (Executive Yuan, Taipei City, Taiwan) with 5% CO2. Each well of the culture received 2 ml of an
and the guidelines as shown in the guide for the Care and Use agar overlay (0.3%) and was incubated for 3 d. Cells were
of Laboratory Animals as promulgated by the Institute of Labo- fixed with 10% formalin, agar overlay was removed, and
ratory Animal Resources (National Research Council). fixed monolayers were stained by crystal violet (0.02% in
https://doi.org/10.4049/immunohorizons.2100020212 ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA ImmunoHorizons
2% ethanol). The results were presented as PFU/ml = cytokine staining, single-cell suspensions (5--10 million cells/
(mean number of plaques 2)/(dilution factor). well of a 24-well plate) were restimulated for 5 h with 100 mg/
ml MHC class II--restricted HA peptide (110SFERFEIFPKE120)
Bacterial load determination for clonotypic CD41 T cells in the presence of 5 mg/ml brefel-
At the indicated days, lungs were collected from infected mice. din A (Sigma-Aldrich). The concentration of brefeldin A was
Each lung was kept in 1 ml DMEM. Serial dilutions of the tis- maintained throughout the intracellular cytokine staining. Re-
sue homogenates were plated to blood agar. Bacterial cultures stimulated cells were surface stained with anti-6.5 and anti-
were then diluted using PBS and the number of viable pneumo- CD4, as described above, fixed in intracellular fixation buffer
cocci was determined using a previously determined standard (eBioscience), washed, and stained in permeabilization buffer
curve (OD at 600 nm versus actual CFU) generated for pneu- (eBioscience) containing fluorochrome-conjugated Abs against
mococci (21). target cytokines. Phosphorylated ZAP-70 and phosphorylated
STAT1 were stained by following protocols provided by the
Histopathology supplier (BD Biosciences). T-bet was stained ex vivo without
The lungs from experimental mice were harvested on the stat- restimulation. All fluorochrome-conjugated Abs were from BD
ed days postinfection and fixed with 10% neutral buffered for- Biosciences, except for T-bet and isotype IgG1k (eBioscience).
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
malin solution. Following fixation, the lungs were embedded in Cells were acquired on FACSCalibur, and analysis was per-
paraffin, and 5-mm sections were cut. Sections were stained formed using CellQuest Pro (BD Biosciences) or FlowJo (Tree
with H&E and scored blindly. The infiltration of inflammatory Star) software.
cells, including lymphocytes, neutrophils, and plasma cells, was
separately scored by the pathologist as negative, 11, 21, or 31, Pneumococcal strain characterization and NanC insertion
according to the density of infiltration. Vasculitis and fibrosis Genomic DNA of the S. pneumoniae serotype 3 clinical isolate,
were also scored as negative, 11, 21, or 31 based on the severi- Taian (21, 22), was prepared, and NanA, NanB, and NanC genes
ty. Overall inflammation in the lungs was represented by the were PCR amplified using the nanA forward (F) (5'-ATA-
average of these scores, as described earlier (17--20). GACGTGCGCAAAATACAGAATCA-3') and reverse (R) (5'GT-
CGAACTCCAAGCCAATAACTCCT-3') primers, nanB F (5'-AC-
Adoptive transfer TACGAGGTGTTAATCGTGAAGG-3') and R (5'-CCAATACCC-
Clonotypic HA-specific CD41 TCR transgenic T cells were pre- GCAGGCATAACATC-3') primers, and the nanC F (5'-TGGG-
pared from pooled spleen and lymph nodes of 6.5 transgenic GTAAGTACAAACAAGAGG-3') and R (5'-CTAATGGTACT-
mice. Clonotypic percentage was determined by flow cytomet- GGCGAAAATCA-3') primers. PCR conditions were 30 cycles of
ric analysis. The naive phenotype was confirmed by profiles of denaturation at 96 C for 30 s, annealing at 55 C for 15 s, and
activation markers CD44 and CD62L. 6.5 CD41 T cells (2.5 extension at 72 C for 45 s when screening for nanA and nanB.
106) were injected through the tail veins of the recipient mice Thirty cycles of denaturation at 96 C for 30 s, annealing at
in 0.2 ml of HBSS (17--20). 51 C for 15 s, and extension at 72 C for 45 s were used to
screen nanC (22).
NA activity on cell surface The nanC-inserted strain was generated by inserting a single
NA activity was measured ex vivo by flow cytometry with sur- copy of the nanC gene into the noncoding region between the
face binding of FITC-coupled peanut agglutinin to the desialy- HMPREF0837_12134 and HMPREF0837_12135. The nanC was
lated cells from the lungs of influenza virus--infected and S. PCR amplified from another clinical isolate using nanC-F_Pspec-
pneumoniae--superinfected mice. Cells from the lungs of naive R (5'-GGTACTAATCAAAATAATGAAAAAAAATATTAAAC-3',
noninfected mice served as controls (17, 18). nanC-R2) and nanC-R (5'-TTAATTCTTTTTCAGATCTTCA-
ATCTTAAATG-3') primers. A promoter region of spectinom
Flow cytometry and intracellular cytokine assay ycin (spec) antibiotic cassette was also PCR amplified from
Cell suspensions of the lungs were incubated on ice with satu- pDL278 plasmid using spec-promoter-F (5'-GAAGATCGA-
rated concentrations of fluorochrome-labeled mAbs in FACS TTTTCGTTCGTG-3') and Pspec-R_nanC-F (5'-GTTTAATA-
buffer (PBS plus 0.5% BSA and 0.02% NaN3). Donor HA-spe- TTTTTTTTCATTATTTTGATTAGTACC -3') primers. Both
cific CD41 T cells were identified using mAbs, as follows: bio- products were fused using spec-promoter-F and nanC-R2 pri-
tin-conjugated anticlonotypic TCR 6.5 (provided by H. Von mers and cloned into pJET 1.2 cloning vector (Thermo Fisher
Boehmer); streptavidin-PE or streptavidin-PerCp; and PerCP- Scientific). The whole spec antibiotic cassette was then PCR
or FITC-conjugated anti-CD4. The anticlonotypic TCR 6.5 Ab amplified from pDL278 plasmid using spec-promoter-F
was replaced by anti-Thy1.1 (CD90.1) Abs in the case of trans- (5'-GAAGATCGATTTTCGTTCGTG-3') and spec-R (5'-GTTAG-
ferring Thy1.11 donor cells into Thy1.2-recipient mice or anti- TAGGCAGTATCTTAATCG-3') primers. The cassette was subcl-
Thy1.2 (CD90.2) Abs in case of transferring Thy1.21 donor cells oned into the NcoI site of Psp-nanC::pJET plasmid to generate
into Thy1.1-recipient mice. FITC-conjugated CD44 and PE-con- Psp-nanC-spec::pJET plasmid. The plasmid was PCR amplified
jugated CD62L were used for surface staining. For intracellular using pJET-F (5'-CGACTCACTATAGGGAGAGCGGC-3') and
https://doi.org/10.4049/immunohorizons.2100020ImmunoHorizons ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA 213 pJET-R (5'-AAGAACATCGATTTTCCATGGCAG-3') primers to prepare a Psp-nanC-spec PCR fragment. The fragment was then ligated into the inverse PCR product of p12133- 12136::pGEM-T plasmid to generate p12133-Psp-nanC-spec- 12136::pGEM-T plasmid. This plasmid was then trans- formed into the wild type S. pneumoniae strain, Taian, and selected with spec (250 mg/L). Statistical analyses Data were represented as mean 6 SD. We used GraphPad Prism version 5 for Student t test analyses. We considered p values
214 ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA ImmunoHorizons
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
FIGURE 4. NanC attenuated the membrane desialylation and TGF-b activation of influenza Ag-specific CD4+ T cells.
Mice received adoptive transfer of naive influenza HA-specific CD41 T cells and influenza virus on the same day. They were superinfected with ei-
ther wild type or NanC-competent S. pneumoniae bacteria on day 4 with control group of mice receiving PBS on day 4. Other control group mice
received adoptive transfer on day 0 and single infection on day 4 with influenza virus, wild type S. pneumoniae, or NanC-competent S. pneumo-
niae. (A) Membrane desialylation and (B) TGF-b activation of HA-specific CD41 T cells in the lungs on day 6, as described in text. Naive cells are
from control mice that received cell transfer but no infection. Histograms are representatives. Values are mean 6 SD of the experimental groups,
normalized by the values in naive cells (n = 6/group. ***p < 0.000, two-tailed p values for unpaired t test).
NanA and NanB of S. pneumoniae suppressed influenza induction (Fig. 5C), and phosphorylation of ZAP-70 and
Ag-specific immunity STAT1 (Fig. 5D, 5E).
We adoptively transferred naive HA-specific CD41 T cells
from the 6.5 CD41 TCR transgenic mice into wild type mice. NanC of S. pneumoniae reversed NanA/NanB-mediated
Recipient mice were infected with influenza virus on the same suppression of influenza Ag-specific immunity
day and superinfected with S. pneumoniae bacteria on day 4. Contrary to the infection of wild type S. pneumoniae, infection
Control groups of recipient mice received infection of influenza of nanC-inserted S. pneumoniae isogenic mutant strain caused
virus or S. pneumoniae bacteria on the same day. Influenza vi- minimum surface desialylation (Fig. 4A, left panel) and no
rus infection caused membrane desialylation and some TGF-b TGF-b activation (Fig. 4B, left panel) of the HA-specific CD41
activation of HA-specific CD41 T cells in the lungs (Fig. 4). T cells. Superinfection with the nanC-inserted pneumococcal
The Th1-specific transcription factor T-bet was also induced in strain also caused only minimal surface desialylation and TGF-
the absolute majority of the HA-specific CD41 T cells in the b activation, which was even less than influenza virus infection
lungs (Fig. 5). Infection of wild type S. pneumoniae did not only (Fig. 4, middle and right panels). Consequently, the Th1
activate HA-specific CD41 T cells (data not shown) but suppression was reversed. There were higher levels of activa-
caused surface desialylation and TGF-b activation to some tion profile CD44highCD62Llow and inflammatory Th1 cytokines
extent (Fig. 4A, 4B, left panels). Both the surface desialyla- IFN-g and TNF-a. There were more T-bet induction and high-
tion and TGF-b activation of HA-specific CD41 T cells er levels of ZAP-70 and STAT1 phosphorylation as well (Fig. 5).
were significantly more with the superinfection of wild type
S. pneumoniae (Fig. 4B, middle panels). The enhanced TGF- NanC mediated aggravated lung pathology, despite more
b response was associated with an attenuated Th1 response comprehensive virus eradication
of HA-specific CD41 T cells, as evidenced by lower levels NanC reversed the NanA/NanB-mediated suppression of influ-
of activation profile CD44highCD62Llow (Fig. 5A), inflamma- enza virus--specific host immunity. There was more efficient
tory Th1 cytokines IFN-g and TNF-a (Fig. 5B), T-bet clearance of the virus. There was less virus in the lungs; even
https://doi.org/10.4049/immunohorizons.2100020ImmunoHorizons ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA 215
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
FIGURE 5. NanC augmented activation of influenza Ag-specific CD4+ T cells.
Naive influenza HA-specific CD41 T cells were adoptively transferred at the time of influenza virus infection. On day 4, mice were superinfected
with wild type or nanC-inserted S. pneumoniae bacteria, and HA-specific CD41 T cells in the lungs were analyzed on day 2 after superinfection, as
described in the text. Compared with the superinfection of wild type, nanC-inserted S. pneumoniae causes expanded CD44highCD62Llow popula-
tion (A), more IFN-g and TNF-a production (B), increased T-bet induction (C), and increased ZAP-70 and STAT1 phosphorylation (D and E) of the
HA-specific CD41 T cells in the lungs. Mice with no infection or with influenza virus infection only served as controls. Dot plots and histograms are
representatives, and values are mean 6 SD, representative of at least three similar experiments (n = 6/group. ***p < 0.0001).
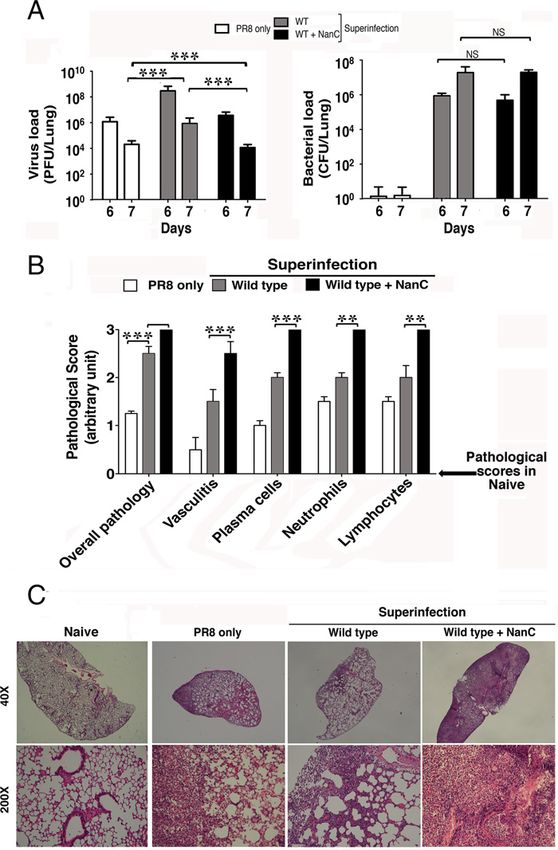
the bacterial loads were comparable (Fig. 6A). However, de-
DISCUSSION
spite much less virus, NanC-competent isogenic S. pneumoniae
aggravated the lung pathology (Fig. 6B). Compared with focal Our results demonstrate altered host--pathogen interaction
inflammation with infiltration of neutrophils, plasma cells, and with the pneumococcal NanC in S. pneumoniae during influen-
lymphocytes inside the alveolar spaces only caused by wild za virus infection with bacterial superinfection. Without NanC,
type S. pneumoniae, NanC-competent isogenic strain caused se- S. pneumoniae superinfection attenuates anti-influenza immuni-
vere diffuse vasculitis (Fig. 6C). ty with increased activation of immunosuppressive cytokine
https://doi.org/10.4049/immunohorizons.2100020216 ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA ImmunoHorizons
pneumococcal NanC, S. pneumoniae superinfection intensi-
fies anti-influenza immunity with suppressed TGF-b acti-
vation and augmented proinflammatory IFN-g and TNF-a
responses of the HA-specific CD41 T cells. Mice clear the
virus more efficiently with the augmented proinflamma-
tory response. Still, they succumb to death even earlier
than the death of mice with superinfection of NanC-defi-
cient S. pneumoniae bacteria. The superinfection of nanC-
inserted S. pneumoniae caused severe lung pathology with
diffused vasculitis and perivascular cuffing.
The a2--3--linked sialosides are on the epithelium surface of
both mouse and human respiratory tract (18, 23). They are the
substrates for NAs of both influenza virus (18, 20) and S. pneu-
moniae (10, 13, 14). We observed more membrane desialylation
of HA-specific CD41 T cells with the superinfection of NanC-
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
deficient S. pneumoniae than that with the infection of influen-
za virus only. It may then be assumed that the NanC may addi-
tionally cleave more sialosides upon superinfection if all three
pneumococcal NAs (NanA, NanB, and NanC) are present in S.
pneumoniae. However, there was less membrane desialylation
with the superinfection of NanC-competent than NanC-defi-
cient S. pneumoniae. NanC is known to produce DANA from
the sialosides, and DANA inhibits the enzymatic activity of
pneumococcal NanA and NanB (10). Perhaps DANA-mediated
inhibition resulted in less membrane desialylation. NanC may
also compete for the same sialoside substrates. The inhibition
and/or competition did not compromise the pneumococcal su-
perinfection as the bacterial loads in the lungs were comparable
between NanC-deficient wild type and NanC-competent iso-
genic S. pneumoniae strains.
The viral NA cleaves latency-associated peptide from the la-
tent TGF-b to activate the immunosuppressive form of the cy-
tokine, active TGF-b, during influenza virus infection (17, 24).
FIGURE 6. NanC enhanced virus eradication, but augmented inflam- Pneumococcal NanA also activates TGF-b during S. pneumoniae
mation caused severe lung pathology and aggravated disease. infection (25). We found augmented TGF-b activation with the
Naive BALB/c mice were infected with 500 PFU PR8 strain H1N1 influ- superinfection of nanC-negative wild type S. pneumoniae. Active
enza virus on day 0 and then superinfected with 50,000 CFU wild type TGF-b is a broad-spectrum immunosuppressive cytokine that
or nanC-inserted S. pneumoniae bacteria on day 4. Mice were sacri- inhibits both innate and adaptive immune responses. It controls
ficed on day 6 or 7, and lungs were collected. (A) Viral and bacterial the innate response by inhibiting NK cells, macrophages, and
loads in the lungs on stated days after viral infection. (B) Lung patholo- neutrophils. It also regulates the adaptive response by inhibit-
gy scores of infected mice, relative to negative scores in naive mice ing Ag presentation of dendritic cells, activation of T cells, and
with mock intranasal PBS, and (C) representative photographs of H&E- suppression of effector T cell function (26--28). Such immune
stained lung sections on day 7 after viral infection. Control mice re- suppression acts as a proviral factor during influenza virus in-
ceived influenza virus on day 0 and PBS on day 4. Naive mice received fection (29). We also found higher virus burden upon superin-
PBS, both on days 0 and 4. Photographs are representatives, and values fection of nanC-negative wild type S. pneumoniae than with
are mean 6 SD, representative of at least three similar experiments influenza virus infection only.
(n = 6/group. **p < 0.001, ***p < 0.0001, two-tailed p values for un- The NanC of S. pneumoniae inhibited TGF-b activation and
paired t test). NS, nonsignificant. unleashed the IFN-g and TNF-a responses of HA-specific
CD41 T cells. Attenuation of anti-influenza immunity by super-
TGF-b in the influenza HA-specific CD41 T cells. Attenuated infection with wild type S. pneumoniae was reversed, and virus
immunity impairs virus clearance, and mice succumb to infec- clearance was more efficient in mice with superinfection of
tion with high virus burden. Bacterial burden also increases in NanC-competent S. pneumoniae. The IFN-g response is the first
this immunosuppressive environment. Alveolar deposits and fo- line of host defense in the control of viral infections (30). The
cal infiltration of immune cells around the alveolar spaces in response helps in virus clearance, and IFN-g supplement pro-
the lungs are associated with their death. In contrast, with tects mice from lethal influenza virus infection (31, 32). It is
https://doi.org/10.4049/immunohorizons.2100020ImmunoHorizons ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA 217
essential for CTL activity, both in acute and memory responses 8. Berry, A. M., and J. C. Paton. 2000. Additive attenuation of viru-
(33, 34). However, IFN-g is a proinflammatory cytokine and ag- lence of Streptococcus pneumoniae by mutation of the genes encod-
ing pneumolysin and other putative pneumococcal virulence
gravates lung inflammation in acute influenza as well (17, 35).
proteins. Infect. Immun. 68: 133--140.
Cellular infiltration in the lungs decreases with IFN-g deficien- 9. Pettigrew, M. M., K. P. Fennie, M. P. York, J. Daniels, and F. Ghaffar.
cy in IFN-g knockout mice. Influenza virus infection still gets 2006. Variation in the presence of neuraminidase genes among
controlled in these mice with enhanced mucosal immunity and Streptococcus pneumoniae isolates with identical sequence types.
alternative Th2-predominant responses (35--37). Superinfection Infect. Immun. 74: 3360--3365.
10. Xu, G., M. J. Kiefel, J. C. Wilson, P. W. Andrew, M. R. Oggioni, and
of NanC-competent S. pneumoniae bacteria intensified IFN-g
G. L. Taylor. 2011. Three Streptococcus pneumoniae sialidases: three
response of the influenza HA-specific CD41 T cells in our ex- different products. J. Am. Chem. Soc. 133: 1718--1721.
periments. The augmented inflammation caused damage and 11. Walther, E., M. Richter, Z. Xu, C. Kramer, S. von Grafenstein, J.
aggravated disease and mortality, even with more comprehen- Kirchmair, U. Grienke, J. M. Rollinger, K. R. Liedl, H. Slevogt, et al.
sive virus eradication. This observation is in line with a previ- 2015. Antipneumococcal activity of neuraminidase inhibiting arto-
carpin. Int. J. Med. Microbiol. 305: 289--297.
ous report that demonstrated the contribution of anti-influenza
12. Gut, H., G. Xu, G. L. Taylor, and M. A. Walsh. 2011. Structural ba-
IFN-g response in the aggravated lung pathology of influenza sis for Streptococcus pneumoniae NanA inhibition by influenza
with S. pneumoniae superinfection (38). Taken together, both
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
antivirals zanamivir and oseltamivir carboxylate. J. Mol. Biol. 409:
virus-mediated damage and immune response--mediated in- 496--503.
flammation are important pathogenic mechanisms for severe 13. Xu, G., X. Li, P. W. Andrew, and G. L. Taylor. 2008. Structure of the
influenza. It is well known that management of inflammation catalytic domain of Streptococcus pneumoniae sialidase NanA. Acta
Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 64: 772--775.
may be as important as control of microbial agents in infectious 14. Xu, G., J. A. Potter, R. J. Russell, M. R. Oggioni, P. W. Andrew, and
diseases. Moderation of inflammation may be even more impor- G. L. Taylor. 2008. Crystal structure of the NanB sialidase from
tant in influenza with superinfection of certain bacteria, such Streptococcus pneumoniae. J. Mol. Biol. 384: 436--449.
as NanC-competent S. pneumoniae. 15. Owen, C. D., P. Lukacik, J. A. Potter, O. Sleator, G. L. Taylor, and
M. A. Walsh. 2015. Streptococcus pneumoniae NanC: structural in-
sights into the specificity and mechanism of a sialidase that produ-
DISCLOSURES ces a sialidase inhibitor. J. Biol. Chem. 290: 27736--27748.
16. Walther, E., Z. Xu, M. Richter, J. Kirchmair, U. Grienke, J. M. Roll-
The authors have no financial conflicts of interest. inger, A. Krumbholz, H. P. Saluz, W. Pfister, A. Sauerbrei, and M.
Schmidtke. 2016. Dual acting neuraminidase inhibitors open new
opportunities to disrupt the lethal synergism between Streptococcus
REFERENCES pneumoniae and influenza virus. Front. Microbiol. 7: 357.
17. Dutta, A., C. T. Huang, T. C. Chen, C. Y. Lin, C. H. Chiu, Y. C. Lin,
1. Louria, D. B., H. L. Blumenfeld, J. T. Ellis, E. D. Kilbourne, and D. E. C. S. Chang, and Y. C. He. 2015. IL-10 inhibits neuraminidase-acti-
Rogers. 1959. Studies on influenza in the pandemic of 1957-1958. II. vated TGF-b and facilitates Th1 phenotype during early phase of in-
Pulmonary complications of influenza. J. Clin. Invest. 38: 213--265. fection. Nat. Commun. 6: 6374.
2. McCullers, J. A., and J. E. Rehg. 2002. Lethal synergism between in- 18. Dutta, A., C. T. Huang, C. Y. Lin, T. C. Chen, Y. C. Lin, C. S. Chang,
fluenza virus and Streptococcus pneumoniae: characterization of a and Y. C. He. 2016. Sterilizing immunity to influenza virus infection
mouse model and the role of platelet-activating factor receptor. J. requires local antigen-specific T cell response in the lungs. Sci. Rep.
Infect. Dis. 186: 341--350. 6: 32973.
3. Morens, D. M., J. K. Taubenberger, and A. S. Fauci. 2008. Predomi- 19. Dutta, A., S. C. Miaw, J. S. Yu, T. C. Chen, C. Y. Lin, Y. C. Lin, C. S.
nant role of bacterial pneumonia as a cause of death in pandemic in- Chang, Y. C. He, S. H. Chuang, M. I. Yen, and C. T. Huang. 2013. Al-
fluenza: implications for pandemic influenza preparedness. J. Infect. tered T-bet dominance in IFN-g-decoupled CD41 T cells with at-
Dis. 198: 962--970. tenuated cytokine storm and preserved memory in influenza. J.
4. Rynda-Apple, A., K. M. Robinson, and J. F. Alcorn. 2015. Influenza Immunol. 190: 4205--4214.
and bacterial superinfection: illuminating the immunologic mecha- 20. Huang, C. T., C. Y. Hung, T. C. Chen, C. Y. Lin, Y. C. Lin, C. S.
nisms of disease. Infect. Immun. 83: 3764--3770. Chang, Y. C. He, Y. L. Huang, and A. Dutta. 2017. Rapamycin adju-
5. Damasio, G. A., L. A. Pereira, S. D. Moreira, C. N. Duarte dos Santos, vant and exacerbation of severe influenza in an experimental mouse
L. M. Dalla-Costa, and S. M. Raboni. 2015. Does virus-bacteria coin- model. Sci. Rep. 7: 4136.
fection increase the clinical severity of acute respiratory infection? 21. Hsieh, Y. C., C. H. Hsiao, P. N. Tsao, J. Y. Wang, P. R. Hsueh, B. L.
J. Med. Virol. 87: 1456--1461. Chiang, W. S. Lee, and L. M. Huang. 2006. Necrotizing pneumococ-
6. Walters, K.-A., F. D’Agnillo, Z.-M. Sheng, J. Kindrachuk, L. M. cal pneumonia in children: the role of pulmonary gangrene. Pediatr.
Schwartzman, R. E. Kuestner, D. S. Chertow, B. T. Golding, J. K. Pulmonol. 41: 623--629.
Taubenberger, and J. C. Kash. 2016. 1918 pandemic influenza virus 22. Hsieh, Y. C., P. N. Tsao, C. L. Chen, T. L. Lin, W. S. Lee, P. L. Shao,
and Streptococcus pneumoniae co-infection results in activation of C. Y. Lee, P. R. Hsueh, L. M. Huang, and J. T. Wang. 2008. Estab-
coagulation and widespread pulmonary thrombosis in mice and hu- lishment of a young mouse model and identification of an allelic
mans. J. Pathol. 238: 85--97. variation of zmpB in complicated pneumonia caused by Streptococ-
7. Camara, M., T. J. Mitchell, P. W. Andrew, and G. J. Boulnois. 1991. cus pneumoniae. Crit. Care Med. 36: 1248--1255.
Streptococcus pneumoniae produces at least two distinct enzymes 23. Walther, T., R. Karamanska, R. W. Chan, M. C. Chan, N. Jia, G. Air,
with neuraminidase activity: cloning and expression of a second C. Hopton, M. P. Wong, A. Dell, J. S. Malik Peiris, et al. 2013. Glyco-
neuraminidase gene in Escherichia coli. Infect. Immun. 59: 2856-- mic analysis of human respiratory tract tissues and correlation with
2858. influenza virus infection. PLoS Pathog. 9: e1003223.
https://doi.org/10.4049/immunohorizons.2100020218 ROLE OF NanC IN S. PNEUMONIAE SUPERINFECTION WITH INFLUENZA ImmunoHorizons
24. Carlson, C. M., E. A. Turpin, L. A. Moser, K. B. O’Brien, T. D. Cline, 32. Weiss, I. D., O. Wald, H. Wald, K. Beider, M. Abraham, E. Galun, A.
J. C. Jones, T. M. Tumpey, J. M. Katz, L. A. Kelley, J. Gauldie, and Nagler, and A. Peled. 2010. IFN-g treatment at early stages of influ-
S. Schultz-Cherry. 2010. Transforming growth factor-b: activation enza virus infection protects mice from death in a NK cell-depen-
by neuraminidase and role in highly pathogenic H5N1 influenza dent manner. J. Interferon Cytokine Res. 30: 439--449.
pathogenesis. PLoS Pathog. 6: e1001136. 33. Turner, S. J., E. Olivas, A. Gutierrez, G. Diaz, and P. C. Doherty.
25. Gratz, N., L. N. Loh, B. Mann, G. Gao, R. Carter, J. Rosch, and E. I. 2007. Disregulated influenza A virus-specific CD81 T cell homeo-
Tuomanen. 2017. Pneumococcal neuraminidase activates TGF-b sig- stasis in the absence of IFN-g signaling. J. Immunol. 178: 7616--7622.
nalling. Microbiology (Reading). 163: 1198--1207. 34. Bot, A., S. Bot, and C. A. Bona. 1998. Protective role of gamma interferon
26. Sanjabi, S., S. A. Oh, and M. O. Li. 2017. Regulation of the immune during the recall response to influenza virus. J. Virol. 72: 6637--6645.
response by TGF-b: from conception to autoimmunity and infec- 35. Graham, M. B., D. K. Dalton, D. Giltinan, V. L. Braciale, T. A. Stew-
tion. Cold Spring Harb. Perspect. Biol. 9: a022236. art, and T. J. Braciale. 1993. Response to influenza infection in mice
27. Flavell, R. A., S. Sanjabi, S. H. Wrzesinski, and P. Licona-Lim on. with a targeted disruption in the interferon gamma gene. J. Exp.
2010. The polarization of immune cells in the tumour environment Med. 178: 1725--1732.
by TGFbeta. Nat. Rev. Immunol. 10: 554--567. 36. Nguyen, H. H., F. W. van Ginkel, H. L. Vu, M. J. Novak, J. R.
28. Batlle, E., and J. Massague. 2019. Transforming growth factor-b sig- McGhee, and J. Mestecky. 2000. Gamma interferon is not required
naling in immunity and cancer. Immunity. 50: 924--940. for mucosal cytotoxic T-lymphocyte responses or heterosubtypic
29. Denney, L., W. Branchett, L. G. Gregory, R. A. Oliver, and C. M. immunity to influenza A virus infection in mice. J. Virol. 74: 5495--
Lloyd. 2018. Epithelial-derived TGF-b1 acts as a pro-viral factor in the 5501.
Downloaded from http://www.immunohorizons.org/ by guest on January 14, 2022
lung during influenza A infection. Mucosal Immunol. 11: 523--535. 37. Liu, B., L. Bao, L. Wang, F. Li, M. Wen, H. Li, W. Deng, X. Zhang, B.
30. Joklik, W. K. 1990. Interferons. In Virology, 2nd Ed., B. N. Fields, Cao. 2019. Anti-IFN-g therapy alleviates acute lung injury induced
and D. N. Knipe, eds. Raven Press, New York, p. 383--410. by severe influenza A (H1N1) pdm09 infection in mice. J. Microbiol.
31. Karupiah, G., J.-H. Chen, S. Mahalingam, C. F. Nathan, and J. D. Immunol. Infect. S1684-1182: 30438--30439.
MacMicking. 1998. Rapid interferon g-dependent clearance of influ- 38. Sun, K., and D. W. Metzger. 2008. Inhibition of pulmonary antibac-
enza A virus and protection from consolidating pneumonitis in nitric terial defense by interferon-gamma during recovery from influenza
oxide synthase 2-deficient mice. J. Exp. Med. 188: 1541--1546. infection. Nat. Med. 14: 558--564.
https://doi.org/10.4049/immunohorizons.2100020You can also read

















































