REVIEW Stability in fig tree-fig wasp mutualisms: how to be a cooperative fig wasp - Oxford ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Biological Journal of the Linnean Society, 2020, 130, 1–17. With 4 figures.
REVIEW
Stability in fig tree–fig wasp mutualisms: how to be a
cooperative fig wasp
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
DEREK W. DUNN*,
College of Life Sciences, Northwest University, Xian, China
Received 25 October 2019; revised 13 February 2020; accepted for publication 14 February 2020
Fig tree–fig wasp mutualisms are diverse and underpin much biodiversity. The wasps (Agaonidae) are the sole
pollinators of the trees (Ficus). Figs are enclosed inflorescences, each of which contains many small flowers. Female
wasps (foundresses) enter receptive figs to spread pollen and to lay their eggs individually into fig flowers. As they
oviposit, wasps also inject chemicals that transform individual flower ovaries into galls that will feed and house
wasp offspring. For fig tree–fig wasp mutualisms to persist, the trees must set seed; therefore, the wasps have both to
pollinate and to fail to gall all flower ovaries. However, wasps that avoid pollination costs and/or gall all flowers are
predicted to outcompete more cooperative conspecifics, resulting in destabilisation of the mutualism. Here, I review
the literature on why wasps pollinate by focusing on how trees reduce investment to unpollinated figs, resulting
in ‘sanctions’ to wasps that fail to pollinate via reduced production of offspring. I also review the mechanisms that
prevent wasps from galling all flowers, mainly those in monoecious Ficus, that also result in wasps predominantly
galling longer flowers whilst leaving shorter flowers to become seeds. I make suggestions for future work and conclude
by reaffirming why multiple processes promote stability in fig tree–fig wasp mutualisms.
ADDITIONAL KEYWORDS: nursery pollination mutualism – symbiosis.
INTRODUCTION developing successfully only by feeding on some of the
reproductive tissue of the plant (Sakai, 2002; Dufray
Mutualisms are cooperative interactions between
& Anstett, 2003). Mutualist reproductive success is
species, in which each partner receives net benefits
therefore tied directly to resources/services provided
(West et al., 2007). Plant–insect pollination mutualisms
by the other mutualist. This provides opportunities
are widespread and underpin a high proportion
to identify mechanisms that promote the stability of
of global biodiversity (Kiers et al., 2010) and crop
mutualism in a more straightforward manner than in
production (Garibaldi et al., 2013). Understanding how
other, less species-specific systems.
these plant–insect interactions remain stable thus has
Fig tree–fig wasp mutualisms are well-known
profound importance.
nursery systems. Each of the 750+ species of fig
Most insect-pollinated plant species receive pollen
tree (Ficus spp.) is pollinated only by the females of
from several insect species, and most species of
a single (or very few) agaonid wasp species (Cook &
pollinating insect vector pollen for multiple plant
Rasplus, 2003; Herre et al., 2008). Individual wasp
species (Waser & Ollerton, 2006). However, nursery
larvae develop successfully only within a single galled
pollination mutualisms are much more species
specific. These mutualisms consist of a single host flower ovary within the enclosed Ficus inflorescence
plant species being pollinated by only a single or very (syconium, hereafter ‘fig’). For fig tree–fig wasp
few insect species, with the larvae of the insect species mutualisms to persist, trees need to produce both
seeds and wasps. The wasps thus have to pollinate
and also to leave some flowers ungalled to ensure
*Corresponding author. E-mail: dwdunn@btinternet.com seed production. However, foundress wasps should, in
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17 1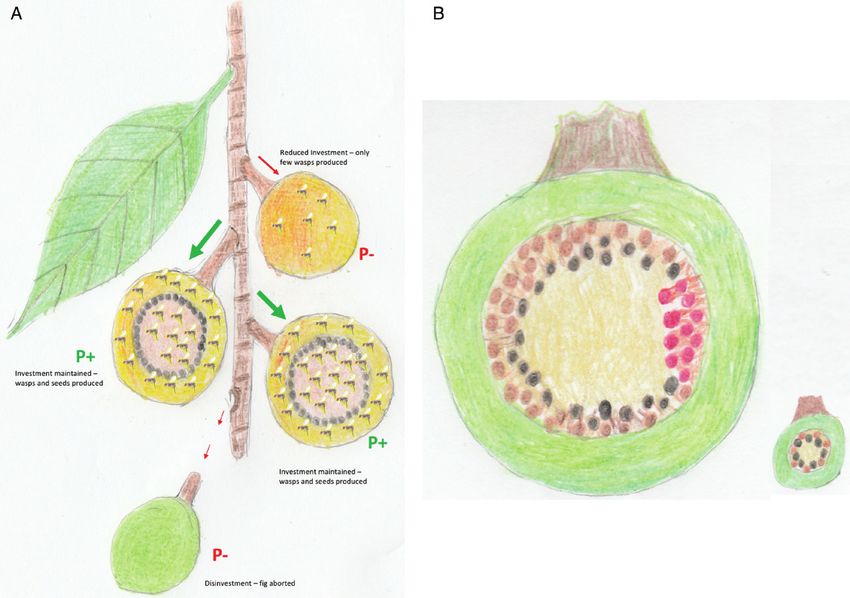
2 D. W. DUNN
theory, do the opposite, because wasps will maximize bract-lined tunnel (ostiole). Once inside the fig cavity
their own short-term fitness by: (1) avoiding costs of (lumen), a foundress will spread pollen onto the flower
pollination; and (2) galling all fig flowers. By doing styles whilst ovipositing (Fig. 1).
so, wasp populations will become dominated by To oviposit, a foundress inserts her ovipositor down
uncooperative individuals, which will destabilize the the style of an individual flower and deposits an egg
mutualism. There is little widespread evidence that into the ovary (Verkerke, 1989; Jansen-Gonzalez et al.,
this has occurred among fig tree–fig wasp mutualisms. 2012; Fig. 1). (In female dioecious figs, the flower
In fact, these mutualisms have radiated and diversified styles are too long for wasps to reach the ovaries.)
on all continents over ≥ 60 Myr (Rønsted et al., 2005) During oviposition, foundresses inject chemicals to
and probably longer (Machado et al., 1996; Xu et al., gall individual ovaries. Each gall supports one larval
2011; Cruaud et al., 2012). Here, I review the literature offspring (Martinson et al., 2014), which eats the
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
on the mechanisms known to promote stability in fig flower endosperm (Jansen-Gonzalez et al., 2012). Adult
tree–fig wasp mutualisms to answer two questions. female pollinating fig wasps are pro-ovigenic (Copland
First, what ensures that the wasps pollinate the trees? et al., 1973; Dunn et al., 2011), short lived (< 48 h;
Second, what prevents the wasps from galling all fig Dunn et al., 2008b; Jevanadam et al., 2013; Sutton
flowers? et al., 2018) and, with few exceptions (e.g. Gibernau
et al., 1996; Moore et al., 2003), die within the first
fig they enter. The lifetime reproductive success of a
foundress is thus almost ubiquitously dependent on a
FIG TREE–FIG WASP BIOLOGY
single bout of oviposition within a single fig (but see
To understand how fig tree–fig wasp mutualisms Moore et al., 2003; Raja et al., 2008; Zhang et al., 2020).
remain stable, general readers need to be familiar
with the reproductive biology of both trees and insects
(see reviews by Janzen, 1979; Weiblen, 1992; Cook & Active and passive pollination
Rasplus, 2003); Herre et al., 2008; Borges, 2015). Approximately 75% of agaonid wasp species are active
pollinators (Kjellberg et al., 2001). Active pollinators
possess pollen-pockets, into which pollen collected from
Monoecy and gynodioecy the relatively few male flowers (Kjellberg et al., 2001)
Fig trees exhibit an approximately equal division into within the natal fig is placed, transported, and then
species that are either monoecious or gynodioecious deposited directly onto the flower styles of another,
(functionally dioecious; Cook & Rasplus, 2003). In receptive fig (Jandér & Herre, 2010). In passive
monoecious species, the pollinating wasps, pollen and pollinators, wasp bodies become covered randomly in
seeds are all produced in the same fig. An individual pollen grains from the many male flowers (Kjellberg
flower ovary can become either a wasp gall or a seed; et al., 2001), which are then transferred indirectly to
therefore, each wasp offspring costs the host tree one the flower styles of another receptive fig (Weiblen,
potential seed (Janzen, 1979). In dioecious species, the 2002; Cook et al., 2004; Jandér & Herre, 2010). Within
pollinating wasps and pollen are produced only in the the figs of most fig tree species, flower styles adhere
figs of ‘male’ trees, with seeds produced only in the together to form a uniform surface, the synstigma.
figs of ‘female’ trees. Gynodioecy has thus effectively When pollen contacts the synstigma, multiple flowers
resolved the conflict between wasp and tree over are thus pollinated (Jousselin & Kjellberg, 2001).
flower fates that occurs in monoecious species (Cook & Passively pollinating fig wasps thus incur no costs of
Rasplus, 2003). The wasps still pollinate and oviposit pollination, and the figs of their associated host tree
in ‘female’ figs but produce no offspring because wasps species, when entered by foundresses, are unlikely
cannot gall the flower ovaries (see below). Female to remain unpollinated. These factors translate to
wasps pollinate female figs because female figs mimic variation in mechanisms promoting pollination in
the chemical attractants produced by conspecific male wasps between passively and actively pollinated host
figs (Soter et al., 2012; Borges, 2015). tree species (Jandér & Herre, 2010; see section below:
Sanctions attributable to non-pollination: variation on
a selective investment theme).
Pollination and oviposition
Figs go through five growth stages (A–E; Galil &
Eisikowich, 1968), three of which (B– D) are associated Adult wasp maturity and dispersal
with the life-cycle of the pollinating wasps (Weiblen, Depending on the species and environmental
2002; Cook & Rasplus, 2003; Fig. 1). When figs become conditions, larvae take from ~3 weeks to several
receptive to pollination (B-stage), female wasps laden months (C-stage) to mature (D.W.D., pers. obs.). Then
with pollen (foundresses) will enter through a narrow, the (D-stage) fig softens, the pollen-laden male flowers
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17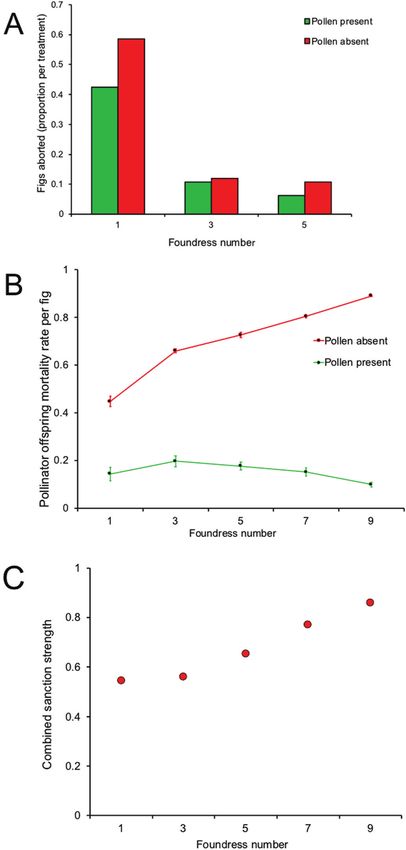
STABILITY IN FIG-WASP MUTUALISMS 3
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
Figure 1. A schematic diagram showing the life-cycle of pollinating fig wasps.
mature, and all of the wasps within emerge from their tree species (Cook & Segar, 2009; Segar et al., 2014;
galls. Adult pollinating fig wasps exhibit a high degree Borges, 2015; Wang et al., 2015a). Non-pollinating fig
of sexual dimorphism. The males are smaller than the wasps can be grouped by size and larval trophic level.
females, are apterous and have much reduced eyes Most species are of similar size to the pollinators,
and mesothoracic legs (Weiblen, 2002). Males emerge oviposit from outside of the fig and can be costly to
from their galls before females (Gu et al., 2014), and either or both mutualists via a reduction in seeds and/
then mate with and release the females from their or pollinator offspring (West & Herre, 1994; West et al.,
galls. Some males will dig an exit tunnel in the fig wall 1996; for reviews, see Weiblen, 2002; Borges, 2015).
and, unless they fall out of this tunnel, in most species Some small NPFW species are parasites (parasitoids
the males do not leave their natal fig. On emergence or kleptoparasitic ‘gall thieves’) of the pollinators; in
from their galls, the females will collect pollen (either several host tree species, the parasitic NPFWs affect the
actively or passively, depending on the species), and flower use patterns of the pollinators, which indirectly
then exit the fig via the tunnels dug by males. The helps to prevent pollinator foundresses from galling all
pollen-laden females will then disperse to another tree fig flowers (Dunn et al., 2008a; Wang et al., 2019; see
that exhibits a crop of receptive figs (Fig. 1). section below: Variation in flower values—the role of
parasitic NPFWs). All parasitic NPFW species cannot
gall fig ovaries and attack their host wasps after the
Non-pollinating fig wasps hosts have oviposited (Martinson et al., 2014; Borges,
In addition to the pollinators, each fig tree species 2015). However, some other small NPFW species can
supports a community of wasp species that all require gall fig ovaries independently and have larvae that
figs to complete their life-cycles (Weiblen, 2002; Cook feed on fig ovary tissue. Some of these ‘small galler’
& Rasplus, 2003; Herre et al., 2008; Borges, 2015). NPFW species oviposit before pollination, which shows
More than 30 of these non-pollinating fig wasp that pollination is not a prerequisite to galling, hence
(NPFW) species can be associated with a single fig to successful wasp reproduction (Borges, 2015).
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17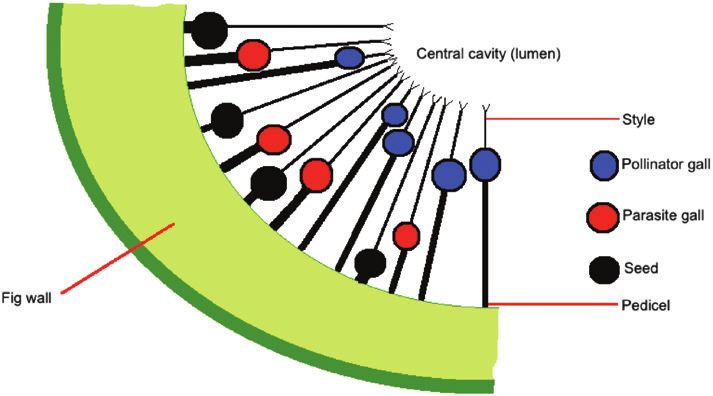
4 D. W. DUNN
Other NPFW species are much larger than the & Herre, 2010; Frederickson, 2013). In other words,
pollinators and have life-cycles that vary in accordance sanctions in fig tree–fig wasp mutualisms are unlikely
with the nutritional requirements of the larvae to have evolved specifically to enforce wasp cooperation
(Weiblen, 2002; Borges, 2015). These NPFWs have (Jandér & Herre, 2010, 2016; Jandér et al., 2012, 2016;
large galls that can reduce the space available within Wang et al., 2014; Zhang et al., 2019). Instead, these
individual figs to pollinator larvae and seeds, but these sanctions are a ‘pre-adaptation’ that evolved to direct
effects are outside the scope of this review. limited resources to those inflorescences that provide
the greatest net benefits to host trees (Jandér & Herre,
2010; Zhang et al., 2019).
In several species of monoecious and male
WHY DO THE WASP SYMBIONTS POLLINATE gynodioecious fig trees, sanctions can result in up to
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
THE TREES? four costs to foundress wasps that fail to pollinate.
First, pollen-free (P−) figs are more likely to abort (figs
Counting the costs and benefits of pollination
drop off of the tree before maturity) than pollinated
In fig tree–fig wasp mutualisms, the rewards provided (P +) figs, resulting in the loss of all wasp offspring
by the host plant to the pollinating insect, namely food (Tarachai et al., 2008; Jandér & Herre, 2010; Jansen-
and housing for the larvae, are accrued by the successful Gonzalez et al., 2012; Wang et al., 2014; Zhang et al.,
dispersal of the adult female insect offspring up to 2019). Second, within non-aborted P− figs, there is an
several months post-pollination (Dufray & Anstett, increased rate of offspring mortality (Jousselin et al.,
2003). Likewise, pollination benefits to the host tree 2003; Jandér & Herre, 2010; Wang et al., 2014; Zhang
are split between those obtained immediately from et al., 2019). Third, within non-aborted P− figs, this rate
pollination, namely seed production (monoecious or of offspring mortality is highest for female offspring
female gynodioecious hosts), and those derived from (Wang et al., 2014; Zhang et al., 2019). Fourth, within
wasp offspring dispersing pollen up to several months non-aborted P− figs, the female offspring are of reduced
later (monoecious and male gynodioecious hosts). All body size (Jandér et al., 2016).
of these processes are contained within individual Increased rates of abortion of P− figs are
figs, which enables fig tree–fig wasp researchers to consistent with trees disinvesting from unpollinated
measure the main costs and benefits of pollination inflorescences (Fig. 2A). This is clearly the costliest
to each mutualist in a more straightforward manner sanction to uncooperative wasps, because all wasp
than in many other plant–pollinator mutualisms. reproductive success is lost (in monoecious and
male gynodioecious figs). However, it is important
to emphasize that in all fig tree species investigated
Sanctions attributable to non-pollination: so far, some P− figs remain non-aborted and some P+
variation on a selective investment theme figs are aborted. Furthermore, monoecious and male
Much recent research on multiple mutualism types, gynodioecious trees will also incur costs of fig abortion,
including fig tree–fig wasp mutualisms, has focused because a tree may still yield some benefits by ‘allowing’
on how hosts promote cooperation in symbiont female wasp offspring to mature and thus disperse
populations by imposing net costs onto uncooperative pollen. Intraspecific variation in rates of fig abortion
symbionts (‘cheaters’ sensu Ghoul et al., 2013; see also probably reflects net benefits to the tree, which are
Jones et al., 2015), processes often termed ‘sanctions’ likely to vary largely but not entirely according to the
(sensu Pellmyr & Huth, 1994; Denison, 2000; Kiers presence of pollen (see Zhang et al., 2019).
et al., 2003, 2011; Weyl et al., 2010; Frederickson, Wasp brood reduction, increased mortality of female
2013). In fig tree–fig wasp mutualisms, sanctions offspring and smaller adult female offspring are all
are likely to select for pollination behaviour in wasps also consistent with selective (reduced) investment
via trees disinvesting completely or partially from to P− figs (Wang et al., 2014; Jandér & Herre, 2016;
unpollinated figs (Jandér & Herre, 2016; Fig. 2A). Jandér et al., 2016; Zhang et al., 2019; Fig. 2A). Agaonid
Given that symbiont wasps interact directly with their larvae feed on the nucellar tissue of normally double-
hosts only within individual figs, this provides trees pollinated flowers (Verkerke, 1989; Jansen-Gonzalez
with the opportunity to target cheaters at the level of et al., 2012). Those larvae developing in P − flowers
the individual fig (but see Jandér et al., 2012; Jandér thus have to feed on the endosperm, which is likely to
& Herre, 2016; Jandér & Steidinger, 2017; Fig. 2A). provide less nutrition. Galling of the ovaries, at least
Angiosperms almost ubiquitously invest preferentially in some fig tree species, is unaffected by pollination
in pollinated inflorescences. The underlying (e.g. Ficus racemosa), but rates of gall failure in P− figs
sanctioning mechanisms to uncooperative symbionts increase significantly (Wang et al., 2014). This suggests
were thus probably present before the evolution and that a lack of pollen results directly in agaonid larval
radiation of fig tree–fig wasp mutualisms (see Jandér mortality after gall formation and provides further
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17STABILITY IN FIG-WASP MUTUALISMS 5
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
Figure 2. A, diagram showing how fig trees invest preferentially in pollinated figs. The attachment of individual figs to
a branch via a peduncle provides trees with opportunities to fine-tune resource investment at the fig level. By reducing
investment to unpollinated figs (P−), trees may alter the cost-to-benefit ratio of supporting figs that will produce no seeds to
their advantage by producing fewer pollen vectors. Alternatively, trees may disinvest completely from P− figs, resulting in
fig abortion. Both scenarios clearly ‘sanction’ uncooperative foundress wasps. B, diagram showing how host tree species that
produce large figs may be able to reduce investment to clusters of flowers galled by foundresses that have failed to pollinate
(here coloured in red), even though other flowers within the same fig have been pollinated by other cooperative foundresses.
This would clearly help to prevent cheater wasps persisting via ‘free-riding’. In host tree species with small figs (inset) this
would be unlikely, owing to fewer foundresses and flowers per fig, resulting in less spatially clustered groups of flowers that
are either pollinated or not.
confirmation that pollination is not a prerequisite exert an additional cost to cheater foundresses (Wang
for galling (Jousselin et al., 2003; see also Jandér & et al., 2014). Small female offspring are less likely than
Herre, 2016). their larger conspecifics to disperse successfully, find
Increased mortality of female offspring (in two another tree that exhibits a crop of receptive figs, and
Asian species; Wang et al., 2014; Zhang et al., 2019) then enter and successfully oviposit within a fig (Liu
and reduced body size of emerged female offspring (in et al., 2011; Jandér et al., 2016). Given that wasp size
four Neotropical species; Jandér et al., 2016) suggest is correlated with egg load (sensu Herre, 1989; D.W.D.,
that the larger female offspring are more sensitive to unpublished observation), adult female wasps from P−
reduced nutrient intake than the males. Fig wasps are figs are also likely to carry reduced egg loads, but this
haplodiploid and actively optimize their offspring sex remains to be tested.
ratios, which are female biased (Cook, 1993; Herre, Rates of fig abortion and offspring mortality within
1985, 1987; Herre et al., 1997; Zhang et al., 2020). High P− figs vary among host tree species (Jandér & Herre,
rates of female offspring mortality will thus constrain 2010; Jansen-Gonzalez et al., 2012; Wang et al., 2014;
the production of optimal offspring sex ratios and Zhang et al., 2019). Importantly, in the two passively
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–176 D. W. DUNN
pollinated host species from which there are published Jandér et al., 2012). Monoecious (and male dioecious)
data, Ficus insipida and Ficus maxima, both costs to trees might still benefit by not sanctioning P − figs,
(experimentally produced) cheater wasps are absent if some wasp offspring vector pollen successfully.
(Jandér & Herre, 2010). This is consistent with figs Therefore, hosts might be accurately deploying
in natural conditions being unlikely to remain pollen mechanisms that (indirectly) sanction cheaters, by
free, owing to foundresses becoming passively covered selectively investing in those inflorescences that will
with pollen within their natal figs. Both F. insipida and yield the hosts net benefits. Such benefits to trees and
F. maxima belong to the Pharmacosycea, a Neotropical wasps might vary in accordance with the number of P+
clade within Ficus that diverged from a clade containing and P− foundresses in a multi-foundress fig. In short,
all other Ficus species ~75 Mya (Cruaud et al., 2012). it might be in the best interest of the tree to ‘tolerate’
All Pharmacosycea species are pollinated passively, the presence of cheaters if the costs of sanctions exceed
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
suggesting that selective investment mechanisms any benefits to the tree.
to unpollinated inflorescences might have been lost Hosts and symbionts in fig tree–fig wasp mutualisms
in the common ancestor of the Pharmacosycea but interact only within individual figs, which in most
retained in the common ancestor of all other Ficus species are attached to a tree branch by a peduncle.
clades (Frederickson et al., 2013). However, the overall This enables trees to manage resource allocation,
picture is more complex, because passive pollination and thus deploy sanctions, at the fig level (Jandér
occurs in several other Ficus clades (Kjellberg et al., et al., 2012; Jandér & Herre, 2016; Fig. 2A). However,
2001; Cook et al., 2004; Cruaud et al., 2012), and an within individual figs, current evidence suggests that
absence of pollen fails to result in costs to wasps in trees do not target the offspring of individual cheater
some non-Pharmacosycea species (e.g. Jousellin & foundresses, i.e. the galled ovaries of unpollinated
Kjellberg, 2001). flowers. This contrasts with the deployment of sanctions
A c r o s s f o u r s y m p a t r i c, a c t i v e l y p o l l i n a t e d in legume–rhizobia mutualisms, in which host plants
monoecious fig tree–fig wasp mutualisms, the natural withhold photosynthate from rhizobia strains that fail
frequency of P− wasps is negatively correlated with to provide nitrogen, whilst maintaining investment to
sanction strength (Jandér & Herre, 2010). Host cooperative strains within the same root mass (Kiers
sanctions might thus determine the proportion of the et al., 2003). For instance, in F. nymphaefolia figs
‘cheater’ phenotype in symbiont populations. However, containing two foundresses, each foundress produces
sanctions were measured only by comparing P+ and on average equal numbers of offspring regardless of
P− single-foundress figs. The strongest sanctions were whether both wasps are P+ or if one is P+ and the other
in Ficus citrifolia, Ficus nymphaeifolia and Ficus P− (Jandér et al., 2012). In F. racemosa, figs entered by
obtusifolia, which on average naturally contain 1.20, a single P− foundress that are subsequently pollinated
2.64 and 1.21 foundresses per fig, respectively (Herre, by seven other ovipositor-excised wasps (that cannot
1989). In all three systems, natural cheater frequency is gall ovaries) produce fewer offspring when pollination
low (< 1%) and can be explained solely by re-occurring is delayed by 24 h than when pollination occurs
mutation for cheating behaviour (Jandér & Steidinger, immediately after the single P− foundress has entered
2017). In the remaining species, Ficus popenoei, which (Wang et al., 2014). Both delayed and immediate
has on average 2.55 foundresses per fig (Herre, 1989), pollination results in more offspring than P− control
sanctions are weaker and natural pollinator cheater figs. Galling remains constant between treatments,
frequency is high (~5%; Jandér & Herre, 2010), meaning but because gall failure shows the reverse trend
that it cannot be explained by mutation alone (Jandér to offspring production, reduced brood sizes can be
& Steidinger, 2017). This suggests that sanctions in attributed directly to increased larval mortality.
F. popenoei are not targeted accurately and that the Delayed pollination also results in rates of fig abortion
natural frequency of cheaters in the population reflects intermediate between immediate pollination and
the point when the relative fitness of cheaters and an absence of pollen. This suggests that reduced
cooperative symbionts is equal; when cheaters become investment to P− F. racemosa figs is initiated by wasp
more common, the net benefits of cheating are thus oviposition but that investment can be reinstated, in
reduced to zero (Jandér & Steidinger, 2017). Although part, by pollination (at least, within 24 h). These host
there are probably additional costs to P− phenotypes sanctions clearly fail to target the offspring of cheaters
(Jandér & Steidinger, 2017), this fails to explain why directly, because in both tree species the cheaters were
F. popenoei hosts fail to sanction single-foundress able to produce full broods when other wasps had
cheaters more effectively and why F. nymphaeifolia pollinated the same fig.
exhibits strong sanctions to single foundresses, even Host sanctions acting at the individual fig level
though this species naturally has multiple foundresses provide an opportunity for P− cheats to reproduce in
per fig (Herre, 1989) and does not sanction individual figs in which at least one other cooperative foundress
cheats in two-foundress figs (i.e. in P+–P− combinations; (P + ) is present. These ‘free-riders’ (Ghoul et al.,
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17STABILITY IN FIG-WASP MUTUALISMS 7
2013) obtain the full benefits of reproducing within
a pollinated fig but will avoid the costs of pollen
collection, transportation and deposition. What then
prevents the spread in wasp populations of cheating
via free-riding in pollinating fig wasp populations?
Among fig tree species, fig size is correlated with
the average number of foundresses (Herre, 1989), with
some tree species with small figs having on average
a number of foundresses less than two (Herre, 1989;
Nedft & Compton, 1996; Derek W. Dunn, unpublished
observations). In addition to having more foundresses,
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
tree species with large figs also have more variable
numbers of foundresses (Herre, 1989; Derek W. Dunn,
unpublished observations). Free-riding is thus unlikely
to outperform cooperation consistently in fig tree–
fig wasp mutualisms in which the host tree species
has small figs, because only few figs contain multiple
foundresses. In single-foundress P− figs, cheaters will be
sanctioned effectively by fig-level reduced investment
by host trees. However, in hosts with large figs, free-
riding opportunities for cheaters will exist, because figs
on average will contain multiple foundresses.
Most studies measuring the effects of the absence of
pollen in fig tree–fig wasp mutualisms have used figs
with only a single P+ or P− foundress (e.g. Jousselin et al.,
2003; Jandér & Herre, 2010; Jansen-Gonzalez et al.,
2012). However, in order to determine whether cheaters
have the potential to outperform cooperative foundresses
via free-riding and to show how sanctions by hosts might
vary according to the number of cheaters, experiments
involving multiple P+ and P− foundresses are required.
To date, such work has mainly been conducted with
F. racemosa, which naturally has large figs (~30 mm in
diameter when mature) containing multiple foundresses
(mean ~4.5; Wang et al., 2014). This has shown that
the sanctioning of cheater wasps is complex. Abortion
rates are highest in P− figs but are negatively correlated
with the number of foundresses; with five or more
foundresses, sanctions cease regardless of the presence
of pollen (Wang et al., 2014; Fig. 3A). In non-aborted
figs, larval mortality rates are positively correlated with
the number of foundresses, but only in P− figs (Fig. 3B),
with overall sanctions (abortion plus increased offspring
Figure 3. Opposing trends in sanctions in Ficus racemosa mortality) being positively correlated with the number
that result in costs to cheater (pollen-free; P−) foundresses, of foundresses (cheaters) per fig (Fig. 3C).
as a function of the number of foundresses introduced These sanctioning patterns are consistent with
experimentally into individual figs. A, fig abortion. The P−
selective investment to inflorescences that result
figs are more likely to abort than P+ figs, but this declines
in net benefits to trees. In tree species whose figs
as the number of foundresses increase. With five or more
tend to contain multiple foundresses, the number of
foundresses, abortion effectively ceases. B, offspring
foundresses tends to be positively correlated with
mortality rates within figs according to the presence of
pollen and number of foundresses. Mortality increases as
the number of foundresses increases, but only for all P− of foundresses; see Jandér & Herre, 2010). Combined
figs. C, combined sanctions (the effects of both abortion sanctions increase as the number of P− foundresses (i.e.
and offspring reduction on P − foundresses compared cheats) per fig increases. Panels are redrawn using data
with foundresses in figs containing the same number presented by Wang et al. (2014).
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–178 D. W. DUNN
both the number of seeds (owing to higher rates of single congeneric cooperative wasp species, with each
pollination) and wasp brood size, i.e. more pollen cheater species being associated with a single host
vectors, within individual figs (Herre, 1989). The fig tree species: Ceratosolen galili–Ficus sycomorus
weakening effect of the number of foundresses on and two undescribed Eupristina species, one with
fig abortion (Fig. 3A) suggests that simply detecting Ficus altissima and the other with Ficus microcarpa.
multiple foundress entry is more cost-effective to Ficus altissima figs often contain foundresses of both
trees to maintain investment in figs than is detecting wasp species (average 1.8 per fig). However, figs with
the presence and/or amount of pollen per se. Ficus a single foundress of either species are common.
racemosa exhibits density-dependent fig receptivity; Importantly, single-foundress Eupristina sp. figs do
to prevent high rates of galling, foundress numbers not have increased rates of abortion (Peng et al., 2008).
are restricted by ostiole closure after initial multi- However, the brood sizes of single cheater Eupristina
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
foundress entry (Wang et al., 2009; see section below: sp. foundresses have been reported as being equal
Why do pollinating wasps not convert all fig flowers (Peng et al., 2008) or ~50% (Peng et al., 2010) those
to wasp galls?). The mechanism responsible may of single cooperative (E. altissima) foundresses. In
thus operate in conjunction with that which affects F. sycomorus, Ceratosolen arabicus (cooperative) and
fig abortion, resulting in the sanctioning patterns C. galili (cheater) foundresses usually occur in figs
found. Alternatively, or additionally, ovipositing wasps in the absence of the other species, even where both
may inject dose-dependent chemicals to prevent fig species are sympatric. The offspring of single C. galili
abortion (Martinson et al., 2014). However, this is foundresses do not incur increased rates of larval
unlikely in F. racemosa because oviposition alone fails mortality, hence reduced brood sizes, nor are they
to affect variation in fig abortion (Wang et al., 2014). subject to increased rates of fig abortion (Compton
Increased larval mortality with increasing numbers et al., 1991).
of cheats (Fig. 3B) suggests that costs of a lack of In the context of host sanctions, it can be deduced
pollen to trees (investing in figs that produce no seeds) that the cheater wasp species of F. altissima and
increasingly outweigh any benefits of pollen vector F. sycomorus are unlikely to be ‘free-riders’, because
production. The positive correlation between combined figs containing single cheater foundresses fail to
sanctions and foundress (cheater) numbers (Fig. 3C), abort, and many wasp offspring can be produced
show that strong reductions in individual foundress (Compton et al., 1991; Peng et al., 2008, 2010). Indeed,
fitness as foundress numbers rise in non-aborted P− in F. sycomorus single C. galili cheater foundresses
figs overrides the opposite trend in patterns of fig produce more offspring than single C. arabicus
abortion (Wang et al., 2014). ‘cooperative’ foundresses (Compton et al., 1991).
Wang et al.’s (2014) results provide a base on The cheater species associated with F. altissima and
which to begin to explain why cheaters (especially F. sycomorus also do not preferentially enter figs that
free-riders) fail to destabilise fig tree–fig wasp already contain at least one cooperative foundress. The
mutualisms. In the Chinese F. racemosa population cheater species of both F. altissima and F. sycomorus
studied, individual P− foundress fitness (offspring per might therefore exhibit traits that enable them to
foundress) only exceeds that of any cooperative P + circumvent host mechanisms that reduce investment
wasps when P− cheaters are single foundresses. This in P− figs, for example via chemicals injected during
is because as foundress numbers increase, foundress oviposition. Alternatively, both host tree species might
fitness is reduced due to intraspecific competition have lost the ability to invest preferentially in P+ figs
for suitable oviposition sites (see below: Why do (sensu Jandér & Herre, 2010; Frederickson, 2013).
pollinating wasps not convert all fig flowers to wasp Further investigations into the effects of pollination
galls?). The P− figs that contain single foundresses and galling by both cooperative and cheater species
have high abortion rates (~60%), but single-foundress within any of these systems are therefore required
figs are relatively rare in the study population (mean (Herre et al., 2008).
number of foundresses is ~4.5; single-foundress figs
~22%; Wang et al., 2014); therefore, cheaters are
unlikely to consistently outperform their cooperative
Host sanctions and suggestions for future
conspecifics (Wang et al., 2014).
work
‘Sanctions’ in male and female gynodioecious figs
Cheater ‘pollinator’ wasp species Most relevant work on gynodioecious Ficus has
There are three known species of agaonid wasp that involved only single-foundress introductions using
fail to pollinate but still gall fig ovaries (Compton ‘male’ trees (e.g. Tarachai et al., 2008). However, the
et al., 1991; Peng et al., 2008, 2010; Wang et al., costs and benefits of pollen absence to host trees and
2015a). Each of these ‘cheater’ species coexists with a wasp symbionts will differ between male and female
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17STABILITY IN FIG-WASP MUTUALISMS 9
trees. P+/P− experiments using both host sexes will pollen-carrying status in determining the variation in
thus be likely to reveal much variation in selective offspring production, and hence the costs vs. benefits
investment, hence the nature of sanctions, but to date of cheating to P− foundresses.
this has been done only with Ficus hispida (Zhang
et al., 2019).
For P− figs, male trees might still benefit by producing Are P− fig wasps obligate or facultative cheats?
some wasps to disperse pollen, but female trees only It is unknown whether cheating behaviour in normally
incur costs, owing to a lack of seed production. In ‘cooperative’ agaonid wasp populations is obligate or
F. hispida, abortion rates of female P− figs are thus facultative (sensu Ghoul et al., 2013). In other words,
much higher than for male P− figs. However, wasps do do cheaters always actively ‘cheat’ and produce
not reproduce in female figs, meaning that cheaters offspring with a propensity to cheat or do some
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
cannot be ‘sanctioned’, even though the mechanism wasps randomly fail to collect pollen? For cheating
responsible for increased rates of abortion does sanction behaviour to respond to selection via host sanctions,
cheaters in male gynodioecious and monoecious figs. it must be at least partly heritable. The presence
Furthermore, the costs of pollen absence to female of both active and passive pollination in agaonids
trees are independent of the number of foundresses, (Kjellberg et al., 2001; Cook et al., 2004) and at least
whereas in male trees the costs may increase as the three instances of a cheater species coexisting with a
number of P− foundresses increases per fig, owing to cooperative, pollinating species (Compton et al., 1991;
the need to support large broods. Given that genes Peng et al., 2008, 2010; Wang et al., 2015a) suggest
contributing to selective investment to inflorescences that this is the case. Experiments to quantify the P+
will be present in both sexes, their expression is likely and P− status of the offspring of naturally occurring
to be a phenotypic compromise. In short, the rates of cheats would enable more accurate estimates of the
P− fig abortion in males are likely to be higher, and in costs and benefits of cheating to both host trees and
females lower, than is optimal for individual males and wasps. These would complement experiments using
females (sensu Arnquist & Rowe, 2002). This can be coexisting cheater and cooperative species in systems
tested comparatively using multiple gynodioecious fig in which cheaters do not incur sanctions (see section
tree–fig wasp mutualisms (Zhang et al., 2019). above: Cheater ‘pollinator’ wasp species).
Multiple introductions of foundresses into large
figs: same number of foundresses, different
proportions of P+ and P−, different order of P+ and WHY DO POLLINATING WASPS NOT
P− entry CONVERT ALL FIG FLOWERS TO
WASP GALLS?
Future experiments should use treatments that have
equal numbers of multiple foundresses that vary in: Across monoecious fig tree species, ~55% of the flowers
(1) their proportions of P+ and P− wasps; and (2) the within individual figs become seeds (Herre, 1989).
order of fig entry according to wasp pollen status. The This is so even in tree species in which figs contain on
host Ficus used should be one that naturally has large average enough wasp eggs to potentially to convert all
figs and multiple foundresses for two reasons. First, flowers into wasp galls, suggesting that mechanisms
within large figs, hosts have more scope to invest are present to restrain wasp galling.
discriminatively in clusters of pollinated flowers than
in smaller figs (Fig. 2B). The introduction of different
numbers of P+ and P− wasps, whilst controlling for total Flower fates within monoecious figs
foundress numbers, will reveal whether host sanctions The flowers within monoecious figs are highly variable
are targeted according to individual foundress in length, which enables optimal packing within the
pollination status, e.g. by genotyping wasp broods usually spherical figs (Ganeshaiah et al., 1995, 1999).
(sensu Jandér et al., 2012). Second, in multi-foundress Most flower ovaries are attached to the inner surface
figs, the first foundress experiences all vacant flowers of the fig wall via a pedicel, although some ovaries may
and will therefore gall those that are most profitable be attached directly to the fig wall. Pedicel lengths and
(see section below: Why do pollinating wasps not style lengths are negatively correlated, i.e. long flowers
convert all fig flowers to wasp galls?). Subsequent nearer the fig lumen have short styles, whereas shorter
foundresses have reduced rates of oviposition, and flowers nearer to or at the fig wall have longer styles
thus reduced reproductive success, owing to: (1) fewer (Jousellin et al., 2004). Flower lengths are normally
profitable flowers; and (2) competition with other distributed, and ovaries are located laterally from the
foundresess (Wang et al., 2009; Dunn et al., 2015). inside surface near the fig wall (‘short flowers’) to near
The order of foundress entry might thus interact with the fig centre (‘long flowers’) (Fig. 4).
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–1710 D. W. DUNN
Across a wide range of fig tree species, within mature evidence that short ovipositors generally prevent wasps
monoecious figs (D-stage; Galil & Eisikowich, 1968) from galling all fig flowers across fig tree–fig wasp
the wasp galls cluster near to the fig centre, with seeds mutualisms. However, further co-evolutionary studies of
at the fig wall (Anstett, 2001; Yu et al., 2004; Dunn flower and ovipositor traits using newer ‘omics’ methods
et al., 2008a, b; Herre et al., 2008; Wang et al., 2019; in genetics might be productive.
Fig. 4), a pattern that clearly results from mechanisms
preventing wasps from galling all flowers. There are
four mechanisms to explain this stratification, each Unbeatable seeds
of which is not mutually exclusive. Importantly, these West & Herre (1994) posited that shorter (long-
may vary among different tree species. styled) flowers have chemical and/or physical barriers
preventing wasp oviposition/galling and are thus
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
‘reserved’ by the tree for seed production. This was
Short ovipositors based on data from six Neotropical fig tree–fig wasp
During oviposition, foundresses inject chemicals from mutualisms showing that pollinating wasps and
their poison gland directly into an ovary, inducing the herbivorous small galler Idarnes NPFWs are in
ovary to form a gall (Martinson et al., 2014). If the direct competition for the same subset of long-styled
longest flower styles exceed the lengths of the wasp flowers, even though most flowers remain unexploited
ovipositors (as in ‘female’ gynodioecious figs), these by any wasps. Furthermore, like most NPFWs (Dunn
flowers cannot be galled. When pollinated, this subset et al., 2008a; Borges, 2015), Idarnes spp. oviposit from
of flowers will thus become seeds. outside of the fig, meaning that fig ovaries that are
Nedft & Compton (1996) tested the ‘short-ovipositor relatively easy to reach, i.e. those nearest the fig wall,
hypothesis’ using 11 African fig tree–fig wasp mutualisms are likely to be avoided (but see section below: Dunn
and found that in B-stage figs (when wasps pollinate et al., 2008a; Al-beidh et al., 2012). Some Idarnes
and oviposit), the ovipositors of nine species could reach wasp species also have very long ovipositors that are
76–100% of the fig flower ovaries. This suggests that inserted through the fig wall, across the lumen and
additional mechanisms to restrict wasp galling are present. down the style of a flower on the opposite side of the fig
Among fig tree–fig wasp mutualisms, style length and to reach an ovary (Elias et al., 2012). The evolution of
wasp ovipositor length are correlated, with evidence of such complex and perhaps costly oviposition strategies
co-evolution of both traits (Weiblen, 2004). As in most are clearly consistent with ‘unbeatable seeds’ if the
mutualisms (Leigh, 2010), host fig trees probably control most accessible fig ovaries are immune from galling.
which of their resources (flowers) are available to their Wang et al. (2013) were the first to test ‘unbeatable
symbionts (wasps). Although wasps undergo multiple seeds’ experimentally, by introducing various numbers
generations per tree generation and can thus respond to of foundresses into F. racemosa figs and giving the
selection more quickly than host trees, the costs of long wasps set times for oviposition. Even when many
ovipositors (e.g. via aerodynamics) might constrain their foundresses were given long enough to gall all fig
evolution (Dunn et al., 2008a). Currently, there is little flowers, wasps still failed to gall short flowers, which
mainly became seeds, supporting ‘unbeatable seeds’.
Martinson et al. (2015) found in F. obtusifolia that
long (short-styled) flowers have higher expression of
genes known to contribute to variation in flavonoids
and carbohydrates than short (long-styled) flowers,
consistent with conditions facilitating galling/larval
feeding. This was suggested as supporting ‘unbeatable
seeds’. However, Martinson et al. (2015) did not show
evidence suggesting that short flowers are immune to
galling. A more in-depth comparative study examining
the chemical defences of flowers of different lengths
and the ability of wasps to sequester plant defence
chemicals, especially involving genomic methods,
would thus be informative (see also section below:
Figure 4. Diagram of one-quarter of a bilaterally sectioned
Optimal ‘foraging’ by foundresses for oviposition sites).
D-stage monoecious fig, showing the spatial segregation
of galls containing pollinator wasps (blue), parasites of
pollinator wasps (either parasitoids or kleptoparasitic
‘gall thieves’; red) and seeds (black). Note the negative
Insufficient eggs
relationship between the lengths of flower pedicels and Pollinating fig wasps often disperse over long distances
styles. See also Dunn et al. (2008a) and Wang et al. (2019). to find a tree with receptive figs (Nason et al., 1998;
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17STABILITY IN FIG-WASP MUTUALISMS 11
Ahmed et al., 2009). There can thus be much variation meaning that fewer flowers are galled per foundress
in the number of foundresses within and among during ‘summer’. When cooler, wasps live for longer,
crops on the same tree (Herre, 1989). If foundresses meaning that fewer foundresses are needed to gall
within individual figs collectively contain fewer eggs the same number of flowers as in summer. This is an
than there are flowers, then this effectively sets the example of host control of its symbiont, a general but
upper limit to how many flowers the wasps can gall. variable mechanism present in mutualisms involving
However, although insufficient eggs might explain diverse taxa (see Leigh, 2010).
why all flowers cannot be galled in some fig tree–fig Although there are currently no published
wasp mutualisms (see below), additional mechanisms comparative studies, ‘insufficient eggs’ may make
are needed in order to explain the spatial segregation a major contribution to stability in fig tree–fig wasp
of wasp galls and seeds in mature figs. mutualisms, especially those that have host tree
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
Explaining wasp gall and seed segregation in the species with large figs. This is because fig size varies
context of ‘insufficient eggs’ relies on a consistent widely among fig tree species (Cook & Rasplus, 2003).
foundress oviposition preference for long flowers. Large figs contain more flowers, have more foundresses
Given that the first foundress enters a fig containing and support larger wasps (with higher egg loads) than
all vacant flowers, short-styled long flowers are galled do smaller figs (Herre, 1989). Owing to tight packing of
first because these allow easier, faster oviposition flowers within the usually spherical figs (Ganeshaiah
than by random oviposition. Subsequent foundresses et al., 1995, 1999), as figs become larger across species
will thus gall ever-shorter (longer-styled) flowers. the increase in flower numbers will be likely to exceed
Even if each foundress lays its complete egg load, if increases in both the number of foundresses and wasp
too few wasp eggs collectively enter a fig than there size. As figs get larger, the difference between the total
are flowers, those flowers that remain ungalled number of wasp eggs entering figs and the number
will probably be the shortest flowers, whose ovaries of flowers within will become increasingly negative.
will mature nearest to the fig wall. Several field ‘Insufficient eggs’ is thus likely to be increasingly
experiments, collectively involving multiple tree and significant as figs and wasps become larger. Moreover,
wasp species, support these predictions (e.g. Jousellin in host tree species with small figs, the difference
et al., 2001; Wang et al., 2013). between the total number of wasp eggs present and
the number of flowers may be positive, with more wasp
Insufficient eggs—restricting foundress entry to eggs present than flowers. Mechanisms restraining
figs: Wasps can gain entry to a fig only through the foundress oviposition rates will thus be more likely to
ostiole; therefore, restricting the time the ostiole occur in species with small figs.
remains open gives trees the potential to manage how
many foundresses, hence the number of wasp eggs, enter Insufficient eggs—keeping pollinators small: Given
individual figs (Anstett et al., 1996). To elaborate, if the that wasp size and fecundity (egg load) are positively
duration for which figs remain receptive to pollinators correlated (Herre, 1989; Dunn et al., 2011), host
is correlated with the number of foundresses, there trees may be able to limit the number of wasp eggs
would be selection for figs to remain receptive for the within their receptive figs, in part, by restricting the
time that allows entry to the number of foundresses average size of the adult wasps. Each wasp has to
that maximizes net benefits to the tree. Although develop from egg to adult within a single galled ovary,
straightforward to describe, empirical testing would be which provides trees with a potential opportunity to
complex, because the optimal number of foundresses constrain wasp gall size. Among the wasp community
will vary among tree species and, probably, within of F. microcarpa at the species level, gall size is
populations of a single species (Herre, 1989; see Wang positively correlated with adult wasp size (Compton
et al., 2009, 2011). et al., 2017). However, to date no study has measured
In F. racemosa, fig receptivity is mediated by the correlation between adult pollinating fig wasp size
seasonal variation in temperature (Wang et al., 2009). and ovary size directly. Furthermore, it is noteworthy
During the subtropical southern Chinese ‘winter’, that because wasps rarely pollinate their natal trees,
when it is relatively cool, fig ostioles close after entry ‘keeping pollinators small’ can only benefit individual
by several foundresses. In summer, ostioles remain trees through inclusive fitness via wasps dispersing
open for longer; therefore, on average, figs contain pollen to other, related trees.
more foundresses in ‘summer’ than in ‘winter’.
Higher temperatures reduce wasp longevity (Wang
et al., 2005; Jevanadam et al., 2013), and interference Optimal ‘foraging’ by foundresses for
competition reduces foundress oviposition rates oviposition sites
(Yu et al., 2004; Wang et al., 2009; see section below: Yu et al. (2004) conceptualized key components of
Optimal ‘foraging’ by foundresses for oviposition sites), both ‘insufficient eggs’ and ‘unbeatable seeds’ to model
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–1712 D. W. DUNN
pollinator galling data using optimal foraging theory pollinator offspring being parasitized decreases from
(Stephens & Krebs, 1987). Briefly, each foundress will > 90% in short flowers near to the fig wall to zero nearest
prefer flowers that confer the highest profitability, to the fig centre (Dunn et al., 2008a). This is because
which is the value of the resource, here the flower some flower ovaries (up to ~25%, depending on the
ovary, divided by the time taken searching for that wasp species) are out of oviposition range to parasitic
resource. The first foundress thus encounters the NPFWs (Al-beidh et al., 2012). Longer (short-styled)
maximal number of highly profitable flowers, but as flowers thus give pollinator offspring enemy-free space
eggs are laid, the profitability of each remaining flower and provide clear evidence of these flowers having the
is reduced owing to the increased time spent searching highest value to foundresses. Moreover, in individual
for the remaining high-value flowers. However, if the F. rubiginosa figs, the abundance of parasitic NPFWs
variation in flower values is high, e.g. if long (short- is positively correlated with the number of seeds (Dunn
Downloaded from https://academic.oup.com/biolinnean/article/130/1/1/5813973 by guest on 16 November 2020
styled) flowers are more valuable than short (long- et al., 2008a), showing that parasites can promote the
styled) flowers, it pays for foundresses still to spend stability of mutualism by helping to prevent symbionts
time searching for vacant long flowers rather than to from fully exploiting host resources (see also Wang
switch to galling short flowers. et al., 2019). Ficus obliqua (Urostigma: Malvanthera)
In figs containing multiple foundresses, as the and F. microcarpa (Urostigma: Conosycea) have
favoured flowers are galled by wasps the number of smaller figs and wasps than F. rubiginosa (Urostigma:
highly profitable flowers becomes depleted relatively Malvanthera), which in turn has smaller figs and wasps
rapidly. Search times therefore increase and oviposition than F. racemosa (Sycomorus) (D.W.D., unpublished
rates are reduced. Owing to short adult lifespans (Dunn observation; Cruaud et al., 2012). Parasitic NPFWs may
et al., 2008b; Wang et al., 2009), some foundresses will thus help to promote stability across a wide range of fig
die before laying their entire egg loads (Kathuria et al., tree–fig wasp mutualisms, because their effects appear
1999; Dunn et al., 2011, 2015). ‘Optimal foraging’ theory to be unconstrained by fig and wasp size or by host tree
can thus account for why figs that contain more wasp phylogeny (Dunn et al., 2008a).
eggs than flowers still produce seeds, and why seeds and Several fig tree species have been translocated
pollinating wasp galls are spatially stratified within figs outside of their natural home ranges. Most also
(Yu et al., 2004; Dunn et al., 2008a, b). have their pollinating wasp species, but they rarely
Empirical investigations of ‘optimal foraging’ fall support any NPFWs (Morgan et al., 2018; Wang et al.,
into two categories: (1) measuring the values of flowers 2019). In translocated F. microcarpa populations in
of different lengths; and (2) measuring the variation in which NPFW parasites of the pollinators are absent,
wasp oviposition patterns as a function of oviposition pollinators have expanded the subset of flowers used
time and/or competition among foundresses. for galling to include some that would otherwise have
become seeds (Wang et al., 2019). Translocated trees
produce fewer seeds per fig than populations in the
Variation in the values of fig flowers of different natural range, which provides further support that
lengths NPFW parasites help to prevent foundress pollinators
Independent of the time taken for foundresses to from fully exploiting fig flowers. Further studies
search for flowers and to lay eggs, flower profitability involving translocated host Ficus species from other
can be affected only by the variation in flower value. clades and/or with larger figs and wasps are required
In turn, the variation in flower value will result from to clarify the general importance of parasitic NPFWs
fitness benefits to offspring varying in accordance with in shaping pollinator flower usage patterns.
flower length.
Variation in flower values—the likelihood of offspring
Variation in flower values—the role of parasitic emergence and flower length: In many Ficus species,
NPFWs: Those NPFWs known to parasitize the within mature (D-stage) figs the flowers near the
pollinators have to insert their long ovipositors from fig centre are less tightly packed than those closer to
outside of the fig in order to reach their hosts within the fig wall (Anstett, 2001; Dunn et al., 2008b). Galls
(Weiblen, 2002; Herre et al., 2008; Borges, 2015). In at containing female pollinator offspring in the tightly
least four fig tree–fig wasp mutualisms, Ficus rubiginosa– packed shorter flowers are thus less likely to be visited
Pleistodontes imperialis, Ficus obliqua–Pleistodontes by the male wasps, resulting in two costs to foundresses
greenwoodii, F. racemosa–Ceratosolen fusciceps (Dunn (Dunn et al., 2008b): (1) female offspring in short
et al., 2008a) and F. microcarpa–Eupristina verticillata flowers are unlikely to be released from their galls; and
(Wang et al., 2019), galls containing NPFW parasites of (2) if female offspring from galls in short flowers are
the pollinators, e.g. Sycoscapter spp. and Philotrypesis released, this occurs after the release of their brood-
spp., lie between the galls containing pollinators and mates in longer flowers, meaning that they are the last
the seeds (Fig. 4). In F. rubiginosa, the likelihood of a female wasp offspring to disperse. Dispersing female
© 2020 The Linnean Society of London, Biological Journal of the Linnean Society, 2020, 130, 1–17You can also read

















































