Revalorization of green tea residues through the production of cellulases by solid-state fermentation using an Aspergillus niger strain.
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Revalorization of green tea residues through the
production of cellulases by solid-state fermentation
using an Aspergillus niger strain.
Salvador Alexis Saldaña ( salvadorsaldana@uadec.edu.mx )
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0002-4039-
7482
Arturo Socrates Palacios Ponce
ESPOL FIMCP: Escuela Superior Politecnica del Litoral Facultad de Ingenieria en Mecanica y Ciencias de la
Produccion
Juan Carlos Contreras Esquivel
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0003-3376-
0674
Raúl Rodríguez Herrera
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0002-6428-
4925
Héctor Arturo Ruiz Leza
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0003-0917-
0324
Juan Alberto Ascacio Valdés
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0001-6595-
863X
José Luis Martínez Hernández
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0002-5158-
7812
Shiburaj Sugathan
University of Kerala https://orcid.org/0000-0001-8185-825X
Cristóbal Noé Aguilar González
Autonomous University of Coahuila: Universidad Autonoma de Coahuila https://orcid.org/0000-0001-5867-
8672
Research Article
Keywords: Cellulases, Solid-state fermentation, Agro-industrial waste, Green tea, β-glucosidase, Aspergillus
niger
Posted Date: May 13th, 2022
Page 1/23DOI: https://doi.org/10.21203/rs.3.rs-1492944/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full
License
Page 2/23Abstract
Green tea is one of the most popular beverages in the world, its industrial production generates huge amounts
of waste, registering 4.8 million tons in 2019, which are a source of environmental pollution. In the present
work, a revalorization of green tea residues (GTR) was explored through cellulase production by solid-state
fermentation (SSF) using a strain of Aspergillus niger 28A. An exploratory analysis of the production of
endocellulases (EC), β-glucosidases (BG), and total cellulase activity (FPUase) was carried out. The effect of
moisture (70-60%), temperature (25-35 °C), and time of fermentation were evaluated. The BG were partially
purified by dialysis and ultracentrifugation, and the optimal pH and temperature for its activity were
determined. It was found that A. niger 28A strain exhibited high titers of EC (65.85 ± 2.18 IU / g), FPUase (5.44 ±
0.11 PFU / g), and BG activities (1016.52 ± 3.58 IU / g). The best production conditions were 34 °C and 96 h for
EC and FPUase activities, and 32 °C, 65 % moisture, and 120 h for BG activity. The BG reached a specific
activity of 496.90 ± 75.04 U / mg after the partial purification process. The optimal temperature and pH for BG
activity from the partially purified extracts were pH 4 and 60 °C. GTR constitutes a suitable substrate for
cellulases production using A. niger 28A strain, reaching high titers of BG, proposing a way for its revalorization
through the generation of products with high added value.
Statement Of Novelty
Green tea is one of the most popular beverages in the world and its industrial production generates enormous
amounts of waste, which are a source of environmental pollution. As far as the authors are aware, this is the
first work that evaluates the fermentation conditions for the production of cellulase enzymes, with a
subsequent partial purification of the β-glucosidase enzymes and determination of the best conditions for the
development of their catalytic activity. This work establishes an alternative for the revalorization of green tea
residues through the generation of high value-added products widely used in the industrial sector.
1. Introduction
Green tea (Camellia sinensis) is a plant of Asian origin [1] currently cultivated in more than 30 countries around
the world [2]. Leaves of this plant are processed through drying treatments to prevent oxidation and are used to
obtain infusions that constitute one of the most popular beverages globally due to its unique flavor and many
beneficial properties that contribute to health [3].
In 2019, a world production of 6.49 million tons of green tea was reported [4], generating 4.8 million tons of
waste [5]. Such wastes are thrown into the environment. In the best case, they are used to elaborate low added-
value products such as fertilizers, animal feed, biodiesel additives, among others [6, 7]. By this reason,
revalorization of tea waste for generation of high added value products is a viable proposal for its use.
On the other hand, enzymes act as biological catalysts allowing the development of specific reactions using
less energy expenditure and shorter times [8]. Cellulases enzymes can degrade cellulose through the hydrolysis
of β 1–4 glycosidic bonds to glucose monomers, and their application stands out in multiple industrial
processes [9], currently constituting the second-largest enzyme by dollar volume [10].
Page 3/23There is information about the use of GTR for the production of cellulases employing SSF, finding that these
residues do not require additional supplementation with mineral enrichment media for the production of these
enzymes using the strain A. niger 28 A [9]. In order to improve the process using GTR for the production of
cellulases, in the present study, different parameters for SSF processes using the strain A. niger 28 A are
analyzed, as well as the partial purification of the BG and determination of the optimal conditions for its
activity.
2. Materials And Methods
2.1 Raw material pretreatment
Green tea was obtained from a local market at the Saltillo, Coahuila; Mexico. The GTR were recovered from the
preparation of infusions at 80 °C for 5 minutes using distilled water and subsequently dried in an oven for 24 h
at 60 °C.
2.2 Reactivation of the strain
The fungal strain used, A. niger 28 A, native to the Western Ghats of India, was provided by the Jawaharlal
Nehru Tropical Botanic Garden & Research Institute (Palode, Thiruvananthapuram, Kerala, India). The strain
was reactivated on PDA agar at 30 °C for 5 days. The spore recovery was carried out using a 1% Tween 80
solution.
2.3.1 Temperature and moisture analysis for cellulase
production
The effect of temperature (°C) and moisture (%) on cellulase production was evaluated using a Hunter & Hunter
exploratory analysis designed through the Statistica 7 program, which generated the treatments shown below:
T1 (25 °C, 70 %), T2 (35 °C, 70%), T3 (25 °C, 60 %) and T4 (35 °C, 60 %). Fermentations were carried out in
triplicate using 250 mL Erlenmeyer flasks with 3 g of GTR and an inoculum of 1X106 spores / g, under the
conditions corresponding to each treatment for 96 h. Extracts were recovered adding 20 mL of 0.05 M citrate
buffer pH 4.8. Then, samples were left stirring at 190 rpm for 10 minutes and recovered by filtration using
Whatman # 1 filter paper. Subsequently, the extracts were centrifuged at 10,000 rpm for 5 minutes. The
supernatants were recovered and stored at -4 °C until use.
2.3.2Evaluation of enzymatic activities
FPUase activity was evaluated using the technique described by Xiao et al. [11]. Briefly, 20 µl of the sample was
mixed with 40 µL of 0.05 M citrate buffer pH 4.8. Whatman # 1 filter paper discs (5.5 mm) were used as the
substrate, then, the reaction was left to incubate for 1 h at 50 °C. After 120 µL of DNS were added and the
sample was boiled for 5 minutes with a subsequent ice bath. Finally, 36 µL of the reaction was mixed with 160
µL of distilled water, and absorbance was measured at 540 nm.
Page 4/23EC activity was evaluated using the technique reported by Ghose (1987) [12] adapted to a microplate [9]. First,
25 µL of the sample was mixed with 25 µL of 2% carboxymethyl cellulose (CMC) (0.05M citrate buffer pH 4.8).
Sample was placed in a water bath for 30 minutes at 50 °C. Then, 150 µL of DNS were added and boiled for 5
min with a subsequent ice bath. After that, 1 mL of distilled water was added, 200 µL were taken, and
absorbance was read at 540 nm.
BG activity was evaluated using the technique reported by Berghem and Pettersson (1974) [13] adapted to
microplate. Briefly, 40 µL of the sample was mixed with 40 µL of a 7.2 mM 4-Nitrophenyl-β-D-glucopyranoside
(PNPG) (0.05 M sodium acetate buffer pH 4.8). After, the sample was placed in a water bath for 10 minutes at
50 °C, and 200 µL of 2% Na2CO3 were added. Subsequently, 200 µL were taken and read on microplate at 410
nm.
2.4 Cellulase enzyme production kinetics
The kinetics were carried out using 250 ml Erlenmeyer flasks with 10 g of GTR at 70% moisture, an inoculum of
1X106 spores/g and allowed to incubate at 30 °C. The experiment ended at 144 h. The extracts were recovered
using the process described above. EC, BG, and FPUase activities were analyzed in each of the evaluated times.
2.5 Enhancement of the incubation temperature to produce EC
and FPUase activities
The optimization temperature was carried out on the EC and FPUase activities production based on the
previous results. Erlenmeyer flasks of 250 ml were used with 10 g of GTR at a humidity of 70% and an
inoculum of 1X106 spores / g. Incubation temperatures from 30 to 38 °C were evaluated. The results were
analyzed using ANOVA, treatment means were compared by a Tukey mean test (p> 0.10), statistical analyzes
were performed using the statistical program InfoStat.
2.6 Influence of the moisture and temperature on the
production of BG activity
From the results obtained in the exploratory analysis and the kinetics of enzyme production, a bifactorial
analysis was carried out for the optimization of BG activity evaluating temperatures of 38, 35, and 32 °C as well
as moisture of 65, 60, and 55% with a fermentation time of 120 h. The results were analyzed using the process
described above in section 2.3.1.
2.7.1 Fermentation in a tray reactor to produce BG
According to the previous stage results, a higher volume fermentation was carried out using a tray reactor. The
fermentation was carried out using 125 g of GTR at a humidity of 60%, inoculum of 1 X 106 spores/ml, 32 °C
and 120 h of incubation. The extracts were recovered using the method described above in section 2.3.1.
Page 5/232.7.2Partial purification of BG
The extracts obtained from the tray fermentation were dialyzed using a Cellu Sep T2 membrane with a pore
size of 6 kDa. Dialysis was performed following the manufacturer's instructions. First, 0.05 M citrate buffer
pH4.8 was used at a ratio of 1:100 for a period of 96 h at 4 °C with continuous stirring. Subsequently, the
dialyzed extracts were ultracentrifuged using Microcon tubes with 100 kDa filters at 14,000 g for 20 minutes at
25 °C following the manufacturer's specifications.
In each of the purification steps, the BG activity was evaluated, and the protein was quantified using the
Bradford method [14]. A Quick Start Bradford protein assay kit (BioRad Laboratories, Hercules, CA, USA) was
used following the manufacturer's instructions. A bovine serum albumin standard (BioRad Laboratories,
Hercules, CA, USA) was used as a standard for protein concentration (0.125 to 2 mg / mL). The results were
used to determine the specific activity.
2.7.3 Influence of temperature and pH on the BG activity of
partially purified enzyme extracts
An analysis on the effect of temperature on BG activity was performed. First, the ultracentrifuged extract was
diluted using 0.05M citrate buffer pH 4.8 in a 1:100 ratio. Subsequently, the BG activity was evaluated by
varying the reaction temperature from 10 to 80 °C.
For the evaluation of the effect of pH on BG activity, the ultracentrifuged extracts was diluted using citrates (pH
3, 4, 5 and 6), phosphates (pH 7 and 8) and Tris-HCL (pH 9) buffers in a 1:100 ratio. Subsequently, the BG
activity was evaluated.
3. Results And Discussion
3.1 Exploratory analysis of the production of cellulase activities
Figure 1 shows results of the exploratory analysis of enzymatic production of A. niger 28A on GTR as support.
In Figure 1 A) it is observed that the T4 treatment showed the highest production of EC activity with 65.85 ±
2.18 IU / g. This result is higher than those reported by various authors presented in Table 1. There is no
significant difference between treatments T4 and T2. On the other hand, in Figure 1 B) it is observed that T2
presented the highest production of FPUase activity with 5.44 ± 0.11 FPU / g and showed a significant
difference concerning the rest of the treatments. This result is slightly lower than that reported by Lodha et al.
(2020) [15], highlighting that these researchers worked with Trichoderma and Penicillium strains, which are
prominent fungi in the production of FPUase activity compared to the Aspergillus genus. Besides, they used a
co-culture technique designed to achieve higher yields than those obtained by pure cultures [16]. On the other
hand, Figure 1C) shows that T4 was the best of the treatments and exhibited a significant difference compared
to the rest, with 1016.52 ± 3.58 IU/g of BG activity. This result is superior to those presented by some authors,
shown in Table 1.
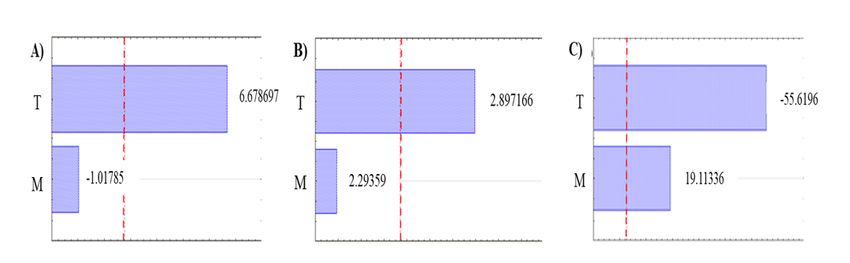
Page 6/23Figure 2 shows the Pareto diagrams of the enzymatic activities evaluated. In the EC (Figure 2 A) and FPUase
activities (Figure 2 B), only the temperature showed significance with a trend towards higher values. On the
other hand, in the BG activity (Figure 2 C), both factors showed significance; the moisture showed a tendency to
the lower values and the temperature to the higher values.
The incubation temperature and initial moisture are some of the few easily controllable factors in SSF
processes, which generally contribute significantly to developing microorganisms and enzyme production [17].
The adequate moisture content in the substrate allows the generation of a layer of water on the particles
surface, facilitating the diffusion and transfer of oxygen and nutrients [18]. Water plays a vital role in the
fermentation processes. It helps the structural maintenance of proteins and amino acids at the molecular level
and the plasma membrane's permeability at the cellular level [19]. In this case, moisture presented a significant
effect only in the production of BG activity allowing its use in the most favorable conditions regarding this
parameter without altering the production titers of EC and FPUase.
Temperature also plays a critical role in SSF on enzymatic production due to the heat produced by metabolic
reactions derived from the difficulty to mix the solids in the medium since most of the substrates used in these
processes have low thermal conductivity [18]. In this study, the temperature had a significant effect on the three
enzymatic activities. It is known that an increase in temperature tends to rise the enzymatic production in
filamentous fungi. Some reports indicate that the optimum temperature for the production of EC by A. niger is
29 °C [18], as well as several studies in which temperatures close to and above 30 °C are used for the
production of cellulases, following the trend described by the exploratory analysis developed in this study [20–
23].
In the case of FPUase activity, results indicated a trend towards higher temperatures, which coincides with that
reported by Akula and Golla (2018) [24], who obtained the highest FPUase activity production at 32 °C working
with a strain of A. niger. In the case of the BG activity, this presented a tendency to high temperatures, similar to
that reported by Saida et al. (2013) [25], who reported the highest BG activity production at 30 °C using an
A.niger strain on sweet sorghum bagasse.
3.2 Cellulase production kinetics.
Figure 3 shows the production kinetics of each of the enzymatic activities evaluated. It is observed that the EC
(Figure 3 A) and the FPUase amounts (Figure 3 B) showed the highest production titer at 96 h (41.95 ± 7.03 IU /
g and 4.87 ± 0.44 IU / g respectively), this coincides with that reported by various authors who indicated that
this was the best time for the production of these activities [22, 23], while the BG (Figure 4C) registered an
increase up to 144 h (1535.17 ± 130 IU / g), which agrees with the investigations carried out by Aliyah et al.
(2017) [26] who reported that the highest production of BG by A. niger using various substrates occurred at 144
h. Figure 3 C) is showed that there is no significant difference between 120 and 144 h for the production of BG
activity; therefore, the incubation period was established at 120 h for subsequent experiments.
This behavior in cellulase production is related to microbial growth phases since fungi tend to produce the
highest titers of EC and FPUase activities during the logarithmic phase, subsequently decreasing during the
stationary phase [22]. Likewise, this behavior is associated with the enzymatic synergy function of cellulases.
Page 7/23EC and exocellulases initiate the breakdown of cellulose for production of cellobiose. The latter inhibits EC and
exocellulase activities through an expression control mechanism known as negative feedback, in which the
final products inhibit the first steps of the metabolic pathways that lead to their production [27]. Subsequently,
the BG are produced to carry out cellobiose degradation into glucose monomers used by the microorganism as
a carbon source [28]. Finally, decrease of enzyme production over time is associated with the lack of nutrients
in the medium and accumulation of toxic wastes that can cause inhibition of fermentation processes and
decrease of microorganisms growth [29].
In the Figure 3 it can be appreciated that there is a slight enzyme production from the beginning of the
fermentation. It is known that transcription of cellulase production genes is regulated by the CreA protein, a
transcriptional repressor of glucose-repressible genes involved in the metabolic pathway, which implies that
there is no significant enzyme production in presence of glucose in the culture medium [30]. However
filamentous fungi have been reported to generate constitutive basal cellulase production. Because cellulose
cannot function as an inducer directly due to its insolubility, this basal production allows degradation of
cellulose, allowing generation of oligosaccharides that act as inducers, promoting a massive production of
cellulases for degradation and use of cellulose [31].
3.3 Analysis of temperature for the production of EC activity
and FPUase activity
Figure 4 shows that EC (4A) and FPUase (4B) activities increase as the incubation temperature rise to 34 °C, at
which these enzymes exhibited their highest activity. In the case of the FPUase activity, this point was
considered as 100% activity for its graphic representation. This result is similar to that reported by [32], who
demonstrated that the best temperature for FPUase activity production by Trichoderma reesei on sugarcane
bagasse was 33 °C. The activity decreased as the temperature approached 40 °C. This phenomenon coincides
with that reported by Akula and Golla (2018) [24], who obtained a decrease in cellulase activity by A. niger as
the temperature approached 40 °C.
It is known that at some points within the SSF, temperature becomes up to 20 °C higher than the incubation
temperature. High temperatures can affect microorganism growth, formation and germination of spores, and
metabolites formation [33]. These temperatures can even denature enzymes and structural proteins by
collapsing the membranous structure [34]. While low temperatures reduce speed of metabolic reactions and
permeability of plasma membrane, reducing exchange of compounds between intracellular and extracellular
environment, thus decreasing transport of nutrients [34]. As a consequence, both situations generate low
enzyme production titers.
3.4 Analysis of the effect of initial moisture and incubation
temperature on the production of β-glucosidase activity.
Effect of temperature and moisture on BG activity is showed in Figure 5. It is appreciated that there is an
interaction between both factors and that the highest enzyme production occurred at 32 °C and 65% moisture,
reaching a titer of 1088.92 ± 54.08 UI / g, higher than those reported by different authors (Table 1). Garcia et al.
Page 8/23(2015) [34] reported that at 65 % moisture, BG production by Lichtheimia ramosa is favored in SSF using wheat
bran as support. Ng et al. (2010) [35] working with a Penicillium citrinum strain in SSF using wheat bran as
support, reported that the best temperature for BG production was at 30 °C.
Moisture is a crucial factor in SSF processes due to its effects on solid particles physical properties [36]. Too
high, moisture content reduces substrate porosity and generates agglomerations, reduces gas volume and its
exchange in the system, decreasing diffusion of oxygen and making difficult to eliminate CO2, while low
contents alter microorganism cellular structure and reduce solubility of nutrients, limiting microorganisms
metabolism and growth and thus secretion of products [19, 36, 37]
The relationship of interaction between temperature and moisture in SSF is crucial. Figure 5 shows that higher
BG production is promoted by high moisture content and low temperatures. The SSF processes present
difficulties in controlling temperature derived from the metabolic heat generated by the microorganism. High
temperatures also promote acceleration of evaporation of the initial water volume, which reduces the amount
of water available in the system, causing the damages mentioned above [17, 33]. Although the strain evaluated
in this trial showed slightly different trends from those reported by other authors, it must be emphasized that, it
is an endemic strain of a region with high climatic fluctuations, so it is natural to appreciate differences in its
behavior.
3.5.1 Fermentation in an aluminum tray reactor for the
production of β-glucosidases
When transferring the experiment from an Elrenmeyer flask to a tray reactor with a larger volume using the best
conditions for the production of BC activity, a decrease in enzyme activity was observed from 1088.93 U / ml ±
54.09 to 659.44 ± 50.39. It is known that SSF processes present these kinds of problems, which makes it
difficult to scale them at an industrial level. This problem is due to, as there is a greater volume of fermentation,
the substrates weight begins to form agglomerations, compacting the material, which hinders gases
transference, dissemination of oxygen, and produces accumulations of CO2. Besides, it reduces elimination of
metabolic heat. As previously mentioned, temperatures reached inside fermentation are higher than those of
incubation [33]. High temperatures can cause protein degradation [38] and promote an increase in the
evaporation rate of water from the system, drastically reducing the initial humidity levels, which can produce
structural alterations of proteins and amino acids at the molecular level and to the plasma membrane at the
cellular level, generating problems of nutrient transfer and waste elimination [19, 36]. These factors produce
stress and difficulties for appropriate microbial metabolism, which is reflected in compounds secretion. To
reduce or eliminate these limitations, bioreactors have been designed with attachments that allow the
regulation of crucial parameters in SSF processes, such as aeration and mixing systems that control oxygen
dissemination and content, and humidification chambers allow the amount of water to be regulated in the
desired parameters for the appropriate microbial metabolism and product secretion [17, 39, 40].
3.5.2 Partial purification of β-glucosidase enzymes
Page 9/23Pre-purification steps of the BG enzymes contained in the crude extracts is showed in Table 2. It is observed
that protein titer was concentrated. Besides, the enzymatic activity showed an increase of almost 25 times
when comparing crude extract with extract in its last phase of partial purification. A production greater than
that obtained by other authors was obtained, such as Karami et al. (2020) [29], who reported 164.001 ± 1.393 U
/ mg when pre-purifying A. niger BG by anion exchange chromatography and cation exchange
chromatography; and Narasimha et al. (2016) [41] who reported 60.6 IU / mg using ammonium sulfate
precipitation techniques and size exclusion chromatography for the purification of BG from A. niger.
It can be seen that the specific activity, as well as the purification factor, showed only a slight increase through
the partial purification steps. This effect is attributed to the fact that it can be observed that the crude extract
has a high specific activity of BG and that a large number of proteins contained in the crude extract have a high
molecular weight greater than that of the filters used for partial purification. In each of the partial purification
steps, small molecules were removed, and the liquid content was decreased, resulting in a high concentration
of BG with a low purification index.
The high specific amount presented by the crude extracts makes it attractive to be used directly in degradation
processes of lignocellulosic material that do not require purified enzymes, which could considerably reduce
processing costs.
3.5.3 Effect of temperature on the BG activity of partially
purified enzyme extracts
BG activity exhibited by partially purified extracts at different temperatures is showed in Figure 8. It is observed
that the enzymatic activity increases as temperature increases to its optimum value at 60 °C, reaching 952.12 ±
46.79 IU / mL, after which it begins to decrease, losing almost completely when reaching 80 °C.
The partially purified extracts presented their highest BG activity at 60 °C, which is similar to that published by
Ali et al. (2016) [42] and Zhao et al. (2013) [43], who reported that the optimum temperature for BG activity was
60 °C working with recombinant enzymes from A. niger cloned and expressed in Pichia pastoris strains. This
result is similar to that presented by other authors in the literature shown in Table 3.
Speed of enzymatic reactions increases as temperature increases due to the rise in the molecules kinetic
energy within the interval in which the enzyme remains stable [44]. The decrease in enzyme activity with an
excessive increase in temperature is generally associated with denaturation of the enzyme active sites in
response to change in environmental temperature [45]. However, the equilibrium model of enzymatic behavior
explains existence of an intermediate stage between irreversible inactivation and denaturation produced by
change of temperature. This transition occurs in shorter periods with imperceptible structural changes that
produce conformational changes in the enzymes active site. In addition to the conformational changes, there
are other factors generated by increase of temperature that affects the enzymatic activity, such as solubility of
gases, affinity for the substrate, development of competition reactions, and pH modification [44]
The pKa values of amino acid side chains are sensitive to temperature changes; basic residues such as lysine
and histidine (common in and around active sites) and terminal amino-n groups undergo large changes in their
Page 10/23charges because of temperature changes. Since ionizable residues from amino acid side chains interact with
charges on adjacent ionized residues, neighboring peptide dipoles, polar residues, and bound water, changes in
temperature have the potential to change charge and its distribution in the active site [46–48].
Enzymes are generally sensitive to extreme temperature conditions, so it is convenient to identify the best
conditions to increase performance without damaging them. The enzymatic hydrolysis reactions developed in
some industries such as sugar, food, textiles, among others, are usually carried out under moderate operating
conditions [26], for which the optimum temperature obtained in this work for BG activity is a desirable outcome.
3.5.4 Effect of pH on the β-glucosidase activity of partially
purified enzyme extracts
Effect of pH on the BG activity of partially purified enzyme extracts is showed in Figure 9. It can be seen that
the highest activity occurs at pH 4, reaching 701.86 ± 2.85 IU / mL and that this begins to decrease as the pH
value increases. This result coincides with that reported by different authors in the literature shown in Table 4.
These results are associated with the acidic conditions in which fungi develop naturally [49]. The results show
that A. niger 28A BGs are highly acidophilic, highlighting their aptitude to be used as biocatalysts in industrial
processes that use extreme acidic conditions such as the fruit juice industry (pH 3.5- 4) [29].
The concentration of hydronium ions in the medium strongly influences the enzymatic activity; these affect
proteins ionization degree, including those found in the active site, influencing the three-dimensional structure
of the enzyme and its affinity for the substrate [44]. It is known that changes in pH induce conformational
changes of the active sites derived from changes in the bonds charges, generating enzymatic inactivation [46].
When the enzymes are at a pH very far from their optimal value, the secondary and tertiary structures are
altered by the protonation and deprotonation of aspartic, glutamic, lysine, arginine, and histidine residues,
mainly causing their unfolding or denaturation in an irreversible way [44].
4. Conclusions
The A. niger 28A strain exhibited high titers of EC, FPUase, and BG enzymes, being the latter in which it
performed more favorably. It was determined that the best conditions for cellulase production were 34°C and
96 h of incubation for EC and (FPUase) activity and 32°C, 65% humidity, and 120 h of incubation for β-
glucosidase activity. The crude extracts produced by A. niger 28 A under the best conditions for the production
of BG activity have high specific activity. Dialysis and ultracentrifugation are methods that allow a high
concentration of BG. The partially purified BG produced by A. niger 28 A showed their optimal activity at pH 4
and 60°C favoring their use in industrial processes developed under acidic conditions such as the fruit juice
industry. In this way, the present work presents an alternative for the revalorization of green tea residues
through the production of cellulase enzyme, which constitutes a high value-added product widely used in the
industrial sector.
Declarations
Acknowledgement
Page 11/23The authors thank the financial support given by the National Council of Science and Technology (CONACYT-
Mexico) through the project FONCICYT-CONACYT-SRE-C0013-2015-03-266614, which was implemented within
a framework of bilateral cooperation between Mexico and India. Author Salvador A. Saldaña Mendoza thanks
CONACYT-Mexico as well as the Autonomous University of Coahuila for the financial support for his MSc
studies.
Funding
This work was funding by the National Council of Science and Technology (CONACYT-Mexico) through the
project FONCICYT-CONACYT-SRE-C0013-2015-03-266614.The funding source had no participation in the
development of the research, writing of the article or in making the decision to submit this work for publication
Competing Interests
Authors declare have no conflicts of interest
Author Contributions
Salvador A. Saldaña-Mendoza: Conceptualization, Methodology, Investigation Writing - Original Draft. Arturo S.
Palacios-Ponce:: Review & Editing. Juan C. Contreras-Esquivel: Resources, Review & Editing. Raúl Rodríguez-
Herrera: Resources, Review & Editing. Héctor A. Ruiz: Resources, Review & Editing. Alberto Ascacio-
Valdés: Resources. José L. Martínez-Hernandez: Review & Editing. Shiburaj Sughatan: Review & Editing. C.N.
Aguilar: Project administration, Funding acquisition, Supervision.
Data availability
All data generated or analyzed during this study are included this published article.
References
1. Moromi, H., Martínez, E.: Efecto del té verde en la formación de la placa bacteriana por Streptococcus
mutans. Odontol. Sanmarquina. 9, 23–24 (2006)
2. Oketch-Rabah, H.A., Roe, A.L., Rider, C. V., Bonkovsky, H.L., Giancaspro, G.I., Navarro, V., Paine, M.F., Betz,
J.M., Marles, R.J., Casper, S., Gurley, B., Jordan, S.A., He, K., Kapoor, M.P., Rao, T.P., Sherker, A.H., Fontana,
R.J., Rossi, S., Vuppalanchi, R., Seeff, L.B., Stolz, A., Ahmad, J., Koh, C., Serrano, J., Low Dog, T., Ko, R.:
United States Pharmacopeia (USP) comprehensive review of the hepatotoxicity of green tea extracts.
Toxicol. Reports. 7, 386–402 (2020). https://doi.org/10.1016/j.toxrep.2020.02.008
3. Zhang, L., Ku, K.M.: Biomarkers-based classification between green teas and decaffeinated green teas
using gas chromatography mass spectrometer coupled with in-tube extraction (ITEX). Food Chem. 271,
450–456 (2019). https://doi.org/10.1016/j.foodchem.2018.07.137
4. Food and Agriculture Organization of the United Nations: FAOSTAT,
http://www.fao.org/faostat/es/#data/QC/visualize
5. Gao, P., Ogata, Y.: CHAMU: An effective approach for improving the recycling of tea waste. IOP Conf. Ser.
Mater. Sci. Eng. 711, (2020). https://doi.org/10.1088/1757-899X/711/1/012024
Page 12/236. Xingfei, L., Shunshun, P., Wenji, Z., Lingli, S., Qiuhua, L., Ruohong, C., Shili, S.: Properties of ACE inhibitory
peptide prepared from protein in green tea residue and evaluation of its anti-hypertensive activity. Process
Biochem. (2020). https://doi.org/10.1016/j.procbio.2020.01.021
7. Bharti, R., Singh, B.: Green tea (Camellia assamica) extract as an antioxidant additive to enhance the
oxidation stability of biodiesel synthesized from waste cooking oil. Fuel. 262, 116658 (2020).
https://doi.org/10.1016/j.fuel.2019.116658
8. Liu, S.: Enzymes. In: Bioprocess Engineering. pp. 229–290. Elsevier (2020)
9. Saldaña-Mendoza, S.A., Ascacio-Valdés, J.A., Palacios-Ponce, A.S., Contreras-Esquivel, J.C., Rodríguez-
Herrera, R., Ruiz, H.A., Martínez-Hernandez, J.L., Sugathan, S., Aguilar, C.N.: Use of wastes from the tea and
coffee industries for the production of cellulases using fungi isolated from the Western Ghats of India.
Syst. Microbiol. Biomanufacturing. (2020). https://doi.org/10.1007/s43393-020-00001-z
10. Patel, A.K., Singhania, R.R., Sim, S.J., Pandey, A.: Thermostable cellulases: Current status and perspectives.
Bioresour. Technol. 279, 385–392 (2019). https://doi.org/10.1016/j.biortech.2019.01.049
11. Xiao, Z., Storms, R., Tsang, A.: Microplate-based filter paper assay to measure total cellulase activity.
Biotechnol. Bioeng. 88, 832–837 (2004). https://doi.org/10.1002/bit.20286
12. Ghose, T.K.: Measurement of cellulase activities. Pure Appl. Chem. 59, 257 – 268 (1987)
13. Berghem, L.E.R., Pettersson, L.G.: The Mechanism of Enzymatic Cellulose Degradation: Isolation and Some
Properties of a β‐Glucosidase from Trichoderma viride. Eur. J. Biochem. 46, 295–305 (1974).
https://doi.org/10.1111/j.1432-1033.1974.tb03621.x
14. Bradford, M.M.: A rapid and sensitive method for the quantitation of microgram quantities of portein
utilizing the pcinciple of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
https://doi.org/10.1016/j.cj.2017.04.003
15. Lodha, A., Pawar, S., Rathod, V.: Optimised cellulase production from fungal co-culture of Trichoderma
reesei NCIM 1186 and Penicillium citrinum NCIM 768 under solid state fermentation. J. Environ. Chem.
Eng. 8, 103958 (2020). https://doi.org/10.1016/j.jece.2020.103958
16. de Oliveira Rodrigues, P., Gurgel, L.V.A., Pasquini, D., Badotti, F., Góes-Neto, A., Baffi, M.A.: Lignocellulose-
degrading enzymes production by solid-state fermentation through fungal consortium among
Ascomycetes and Basidiomycetes. Renew. Energy. 145, 2683–2693 (2020).
https://doi.org/10.1016/j.renene.2019.08.041
17. Mitchell, D.A., Sugai-Guérios, M.H., Krieger, N.: Solid-State Fermentation. In: Reference Module in Chemistry,
Molecular Sciences and Chemical Engineering. pp. 1–9. Elsevier Inc. (2019)
18. Santos, T.C. dos, Abreu Filho, G., Brito, A.R. de, Pires, A.J.V., Bonomo, R.C.F., Franco, M.: Production and
Characterization of Cellulolytic Enzymes By Aspergillus Niger and Rhizopus Sp. By Solid State
Fermentation of Prickly Pear. Rev. Caatinga. 29, 222–233 (2016). https://doi.org/10.1590/1983-
21252016v29n126rc
19. Gervais, P., Molin, P.: The role of water in solid-state fermentation. Biochem. Eng. J. 13, 85–101 (2003).
https://doi.org/10.1016/S1369-703X(02)00122-5
20. Xue, D. sheng, Zeng, X., Lin, D., Yao, S.: Ethanol tolerant endoglucanase from Aspergillus niger isolated
from wine fermentation cellar. Biocatal. Agric. Biotechnol. 15, 19–24 (2018).
https://doi.org/10.1016/j.bcab.2018.04.016
Page 13/2321. Boggione, M.J., Allasia, M.B., Aguilar, C.N., Farruggia, B.: Valorization of corn cob for the obtention and
purification of endoglucanase produced by SSF. Process Biochem. 88, 106–112 (2020).
https://doi.org/10.1016/j.procbio.2019.09.026
22. Sohail, M., Siddiqi, R., Ahmad, A., Khan, S.A.: Cellulase production from Aspergillus niger MS82: effect of
temperature and pH. N. Biotechnol. 25, 437–441 (2009). https://doi.org/10.1016/j.nbt.2009.02.002
23. Kang, S.W., Park, Y.S., Lee, J.S., Hong, S.I., Kim, S.W.: Production of cellulases and hemicellulases by
Aspergillus niger KK2 from lignocellulosic biomass. Bioresour. Technol. 91, 153–156 (2004).
https://doi.org/10.1016/S0960-8524(03)00172-X
24. Akula, S., Golla, N.: Optimization of Cellulase Production by Aspergillus niger Isolated from Forest Soil.
Open Biotechnol. J. 12, 256–269 (2018). https://doi.org/10.2174/1874070701812010256
25. Saida, L., Oberoi, H.S., Narasu, M.L.: Studies on Cellulase Production by Solid state Fermentation using
Sweet Sorghum bagasse Abstract :, (2013)
26. Aliyah, A., Alamsyah, G., Ramadhani, R., Hermansyah, H.: Production of α-Amylase and β-Glucosidase from
Aspergillus Niger by solid state fermentation method on biomass waste substrates from rice husk,
bagasse and corn cob. Energy Procedia. 136, 418–423 (2017).
https://doi.org/10.1016/j.egypro.2017.10.269
27. Dos Santos, T.C., Gomes, D.P.P., Bonomo, R.C.F., Franco, M.: Optimisation of solid state fermentation of
potato peel for the production of cellulolytic enzymes. Food Chem. 133, 1299–1304 (2012).
https://doi.org/10.1016/j.foodchem.2011.11.115
28. Behera, B.C., Sethi, B.K., Mishra, R.R., Dutta, S.K., Thatoi, H.N.: Microbial cellulases – Diversity &
biotechnology with reference to mangrove environment : A review. J. Genet. Eng. Biotechnol. 15, 197–210
(2017). https://doi.org/10.1016/j.jgeb.2016.12.001
29. Karami, F., Ghorbani, M., Sadeghi Mahoonak, A., Khodarahmi, R.: Fast, inexpensive purification of β-
glucosidase from Aspergillus niger and improved catalytic/physicochemical properties upon the enzyme
immobilization: Possible broad prospects for industrial applications. Lwt. 118, 108770 (2020).
https://doi.org/10.1016/j.lwt.2019.108770
30. Rajoka, M.I., Akhtar, M.W., Hanif, A., Khalid, A.M.: Production and characterization of a highly active
cellobiase from Aspergillus niger grown in solid state fermentation. World J. Microbiol. Biotechnol. 22,
991–998 (2006). https://doi.org/10.1007/s11274-006-9146-0
31. Suto, M., Tomita, F.: Induction and catabolite repression mechanisms of cellulase in fungi. J. Biosci.
Bioeng. 92, 305–311 (2001). https://doi.org/10.1016/S1389-1723(01)80231-0
32. Mekala, N.K., Singhania, R.R., Sukumaran, R.K., Pandey, A.: Cellulase production under solid-State
fermentation by trichoderma reesei RUT C30: Statistical optimization of process parameters. Appl.
Biochem. Biotechnol. 151, 122–131 (2008). https://doi.org/10.1007/s12010-008-8156-9
33. Pandey, A.: Solid-state fermentation. Biochem. Eng. J. 13, 81–84 (2003). https://doi.org/10.1016/S1369-
703X(02)00121-3
34. Garcia, N.F.L., da Silva Santos, F.R., Gonçalves, F.A., da Paz, M.F., Fonseca, G.G., Leite, R.S.R.: Production of
β-glucosidase on solid-state fermentation by Lichtheimia ramosa in agroindustrial residues:
Characterization and catalytic properties of the enzymatic extract. Electron. J. Biotechnol. 18, 314–319
(2015). https://doi.org/10.1016/j.ejbt.2015.05.007
Page 14/2335. Ng, I.S., Li, C.W., Chan, S.P., Chir, J.L., Chen, P.T., Tong, C.G., Yu, S.M., Ho, T.H.D.: High-level production of a
thermoacidophilic β-glucosidase from Penicillium citrinum YS40-5 by solid-state fermentation with rice
bran. Bioresour. Technol. 101, 1310–1317 (2010). https://doi.org/10.1016/j.biortech.2009.08.049
36. Noor El-Deen, A.M., Shata, H.M.A.H., Farid, M.A.F.: Improvement of β-glucosidase production by co-culture
of Aspergillus niger and A. oryzae under solid state fermentation through feeding process. Ann. Microbiol.
64, 627–637 (2014). https://doi.org/10.1007/s13213-013-0696-8
37. Pandey, A., Ashakumary, L., Selvakumar, P., Vijayalakshmi, K.S.: Influence of water activity on growth and
activity of Aspergillus niger for glycoamylase production in solid-state fermentation. World J Microbiol
Biotechnol. 10, 485–486 (1994). https://doi.org/10.1007/BF00144481
38. Singhania, R.R., Patel, A.K., Soccol, C.R., Pandey, A.: Recent advances in solid-state fermentation. Biochem.
Eng. J. 44, 13–18 (2009). https://doi.org/10.1016/j.bej.2008.10.019
39. Soccol, C.R., Costa, E.S.F. da, Letti, L.A.J., Karp, S.G., Woiciechowski, A.L., Vandenberghe, L.P. de S.: Recent
developments and innovations in solid state fermentation. Biotechnol. Res. Innov. 1, 52–71 (2017).
https://doi.org/10.1016/j.biori.2017.01.002
40. Arora, S., Rani, R., Ghosh, S.: Bioreactors in solid state fermentation technology: Design, applications and
engineering aspects. J. Biotechnol. 269, 16–34 (2018). https://doi.org/10.1016/j.jbiotec.2018.01.010
41. Narasimha, G., Sridevi, A., Ramanjaneyulu, G., Rajasekhar Reddy, B.: Purification and Characterization of β-
Glucosidase from Aspergillus Niger. Int. J. Food Prop. 19, 652–661 (2016).
https://doi.org/10.1080/10942912.2015.1023398
42. Ali, N., Xue, Y., Gan, L., Liu, J., Long, M.: Purification, characterization, gene cloning and sequencing of a
new β-glucosidase from Aspergillus niger BE-2. Appl. Biochem. Microbiol. 52, 564–571 (2016).
https://doi.org/10.1134/S0003683816050045
43. Zhao, L., Zhou, T., Li, X., Fan, S., You, L.: Expression and characterization of GH3 β-Glucosidase from
Aspergillus niger NL-1 with high specific activity, glucose inhibition and solvent tolerance. Microbiol.
(Russian Fed. 82, 356–363 (2013). https://doi.org/10.1134/S0026261713030181
44. Quirasco Brauch, M., López-Mungía Canales, A.: Enzimas. In: Quintanar Duarte, E. (ed.) Química de los
alimentos. pp. 301–362. PEARSON EDUCACIÓN, México (2006)
45. Ezeilo, U.R., Lee, C.T., Huyop, F., Zakaria, I.I., Wahab, R.A.: Raw oil palm frond leaves as cost-effective
substrate for cellulase and xylanase productions by Trichoderma asperellum UC1 under solid-state
fermentation. J. Environ. Manage. 243, 206–217 (2019). https://doi.org/10.1016/j.jenvman.2019.04.113
46. Daniel, R.M., Danson, M.J.: Temperature and the catalytic activity of enzymes: A fresh understanding.
FEBS Lett. 587, 2738–2743 (2013). https://doi.org/10.1016/j.febslet.2013.06.027
47. Peterson, M.E., Eisenthal, R., Danson, M.J., Spence, A., Daniel, R.M.: A new intrinsic thermal parameter for
enzymes reveals true temperature optima. J. Biol. Chem. 279, 20717–20722 (2004).
https://doi.org/10.1074/jbc.M309143200
48. Danson, M.J., Hough, D.T., Russell, R.J.M., Taylor, G.L., Pearl, L.: Enzyme thermostability and thermoactivity.
Protein Eng. 9, 629–630 (1996). https://doi.org/10.1093/protein/9.8.629
49. Ang, S.K., Shaza, E.M., Adibah, Y.A., Suraini, A.A., Madihah, M.S.: Production of cellulases and xylanase by
Aspergillus fumigatus SK1 using untreated oil palm trunk through solid state fermentation. Process
Biochem. 48, 1293–1302 (2013). https://doi.org/10.1016/j.procbio.2013.06.019
Page 15/2350. Wattanakitjanukul, N., Sukkasem, C., Chiersilp, B., Boonsawang, P.: Use of Palm Empty Fruit Bunches for
the Production of Ligninolytic Enzymes by Xylaria sp. in Solid State Fermentation. Waste and Biomass
Valorization. 11, 3953–3964 (2020). https://doi.org/10.1007/s12649-019-00710-0
51. Shruthi, K., Yadav, P.S., Prasad, B.V.S., Chandra, M.S.: Cellulase production by Aspergillus unguis in solid
state fermentation. J. For. Res. 30, 205–212 (2019). https://doi.org/10.1007/s11676-018-0619-4
52. Xu, X., Lin, M., Zang, Q., Shi, S.: Solid state bioconversion of lignocellulosic residues by Inonotus obliquus
for production of cellulolytic enzymes and saccharification. Bioresour. Technol. 247, 88–95 (2018).
https://doi.org/10.1016/j.biortech.2017.08.192
53. Dias, L.M., dos Santos, B. V., Albuquerque, C.J.B., Baeta, B.E.L., Pasquini, D., Baffi, M.A.: Biomass sorghum
as a novel substrate in solid-state fermentation for the production of hemicellulases and cellulases by
Aspergillus niger and A. fumigatus. J. Appl. Microbiol. 124, 708–718 (2018).
https://doi.org/10.1111/jam.13672
54. Leite, P., Salgado, J.M., Venâncio, A., Domínguez, J.M., Belo, I.: Ultrasounds pretreatment of olive pomace to
improve xylanase and cellulase production by solid-state fermentation. Bioresour. Technol. 214, 737–746
(2016). https://doi.org/10.1016/j.biortech.2016.05.028
55. Crognale, S., Liuzzi, F., D’Annibale, A., de Bari, I., Petruccioli, M.: Cynara cardunculus a novel substrate for
solid-state production of Aspergillus tubingensis cellulases and sugar hydrolysates. Biomass and
Bioenergy. 127, 105276 (2019). https://doi.org/10.1016/j.biombioe.2019.105276
56. Prajapati, B.P., Kumar Suryawanshi, R., Agrawal, S., Ghosh, M., Kango, N.: Characterization of cellulase
from Aspergillus tubingensis NKBP-55 for generation of fermentable sugars from agricultural residues.
Bioresour. Technol. 250, 733–740 (2018). https://doi.org/10.1016/j.biortech.2017.11.099
57. das Neves, C.A., de Menezes, L.H.S., Soares, G.A., dos Santos Reis, N., Tavares, I.M.C., Franco, M., de
Oliveira, J.R.: Production and biochemical characterization of halotolerant β-glucosidase by Penicillium
roqueforti ATCC 10110 grown in forage palm under solid-state fermentation. Biomass Convers. Biorefinery.
(2020). https://doi.org/10.1007/s13399-020-00930-8
58. Nishida, V.S., de Oliveira, R.F., Brugnari, T., Correa, R.C.G., Peralta, R.A., Castoldi, R., de Souza, C.G.M., Bracht,
A., Peralta, R.M.: Immobilization of Aspergillus awamori β-glucosidase on commercial gelatin: An
inexpensive and efficient process. Int. J. Biol. Macromol. 111, 1206–1213 (2018).
https://doi.org/10.1016/j.ijbiomac.2018.01.146
59. Xie, Y., Gao, Y., Chen, Z.: Purification and characterization of an extracellular β-glucosidase with high
transglucosylation activity and stability from Aspergillus niger No. 5.1. Appl. Biochem. Biotechnol. 119,
229–240 (2004). https://doi.org/10.1007/s12010-004-0004-y
60. Watanabe, A., Suzuki, M., Ujiie, S., Gomi, K.: Purification and enzymatic characterization of a novel β-1,6-
glucosidase from Aspergillus oryzae. J. Biosci. Bioeng. 121, 259–264 (2016).
https://doi.org/10.1016/j.jbiosc.2015.07.011
61. Krogh, K.B.R.M., Harris, P. V., Olsen, C.L., Johansen, K.S., Hojer-Pedersen, J., Borjesson, J., Olsson, L.:
Characterization and kinetic analysis of a thermostable GH3 β-glucosidase from Penicillium brasilianum.
Appl. Microbiol. Biotechnol. 86, 143–154 (2010). https://doi.org/10.1007/s00253-009-2181-7
62. Leite, R.S.R., Gomes, E., da Silva, R.: Characterization and comparison of thermostability of purified β-
glucosidases from a mesophilic Aureobasidium pullulans and a thermophilic Thermoascus aurantiacus.
Page 16/23Process Biochem. 42, 1101–1106 (2007). https://doi.org/10.1016/j.procbio.2007.05.003
Tables
Table 1. Production of cellulases by solid-state fermentation using different substrates
Page 17/23Substrate Strain Enzyme Activity Reference
Palm empty fruit bunches Xylaria sp EC 9.93 UI [50]
/g
Groundnut fodder A. unguis EC 1,1 UI / [51]
g
Wheat bran Inonotus obliquus EC 27.15 [52]
UI / g
Biomass sorghum A. niger SCBM1 EC 41.47 [53]
UI / g
Exhausted olive pomace A. niger EC 38 UI / [54]
g
Sugar cane bagasse Co-culture de A. niger NRC 7A y A. oryzae EC 1.38 UI [36]
NRRL 447 /g
Wheat bran Co-culture Trichoderma reesei NCIM FPUase 6.71 [15]
1186 and Penicillium citrinum NCIM 768 FPU /g
Palm empty fruit bunches Xylaria sp FPUase 0.49 [50]
FPU/g
Groundnut fodder Aspergillus unguis FPUase 5.9 [51]
FPU /
g
Cynara cardunculus A. tubingensis NRRL4700 FPUase 4 [55]
FPU/g
Copra meal A. tubingensis NKBP-55 FPUase 3.8 [56]
FPU /
g
Wheat bran Inonotus obliquus FPUase 3.16 [52]
FPU/g
Sugar cane bagasse Co-culture de A. niger NRC 7A y A. oryzae FPUase 1.0 [36]
NRRL 447 FPU /
g
Forage palm (Nopalea Penicillium roqueforti ATCC 10110 BG 935.07 [57]
UI / g
cochenillifera)
Palm empty fruit bunches Xylaria sp BG 21.5 UI [50]
/g
Cynara cardunculus A. tubingensis NRRL4700 BG 25 UI / [55]
g
A mixture of pineapple A. awamori BG 820 UI [58]
crown leaves and wheat /g
bran
Biomass sorghum A. niger SCBM1 BG 54.9 UI [53]
/g
Wheat bran Inonotus obliquus BG 2.53 UI [52]
Page 18/23/g
Corn cob A. niger BG 286.05 [26]
UI / g
Sugar cane bagasse Co-culture de A. niger NRC 7A y A. oryzae BG 1289 [36]
NRRL 447 UI / g
(EC)endocellulase activity, (FPUase) total cellulase activity and (BG) β-glucosidase activity.
Table 2. Partial purification steps of A. niger 28A β-glucosidases produced using GTR as support in SSF
processes
Volume Protein Protein Enzymatic Total Specific Yield Purifica-
activity activity activity tion
mL mg/mL mg % factor
U/mL U U/mg
Crude extract 20.00 0.22 ±0 4.33 ± 76.93 ± 1538.58 355.00 100.00 1.00
01 0.34 5.88 ± 65.47 ± 44.53
Dialyzed 10.00 0.17 ± 1.67 ± 60.36 ± 603.57 361.95 39.23 1.02
0.00 0.00 7.79 ± 95.36 ± 57.18
Ultracentrifuged 0.25 1.27 ± 0.32 ± 632.22 ± 158.05 496.90 10.27 1.40
0.02 0.01 12.81 ± 27.04 ± 75.04
Total activity =Enzymatic activity (U/mL) * Volume (mL)
Specific activity= Total activity (U) / Protein (mg)
Yield = (Total activity of the purification step (U) / Total activity of crude extract (U)) *100
Purification factor = Specific activity of purification step (U/mg) / Specific activity of crude extract (U/mg)
Table 3. Optimal temperature of β-glucosidase activity reported by different authors
Optimal temperature °C Strain Reference
70 A. tubingensis [56]
30 A. niger [26]
65 Lichtheimia ramosa [34]
70 Penicillium citrinum [35]
60 niger [59]
Page 19/23Table 4. Optimal pH for β-glucosidase activity reported by different authors
Optimal pH Strain Reference
4.5 A. oryzae [60]
5.5 L. ramosa [34]
4.8 Penicillium brasilianum (recombinant strain) [61]
4 Pichia pastoris (recombinant strain) [43]
4 Aspergillus fumigatus [49]
4.0-4.5 Aureobasidium pullulans [62]
4 Thermoascus aurantiacus
Figures
Figure 1
Exploratory analysis of the influence of temperature (°C) and moisture (%) on the enzymatic production of
Aspergillus niger 28A on GTR as support. A) Endocellulase activity, B) Total cellulase activity C) β-glucosidase
activity (T1 25 °C, 70%; T2 35 °C, 70%; T3 25 °C, 60%; T4 35 °C, 60%). The standard deviations are represented
by vertical lines in each of the bars and the statistical differences by letters.
Page 20/23Figure 2
Pareto diagram, exploratory analysis, production of enzymatic activities by Aspergillus niger 28A using GTR as
support. A) EC, B) FPUase, C) BG activities (T-temperature, M-moisture). The reference line indicates which
elements are statistically significant at p> 0.05.
Page 21/23Figure 3
Kinetics of enzyme activity production by A. niger 28A using GTR as support. A) EC, B) FPUase, C) BG activities.
The standard deviations are represented by vertical lines at each of the kinetic points and the statistical
differences by letters.
Figure 4
Effect of incubation temperature on the production of A) EC activity and B) FPUase activity by SSF using the A.
niger 28A strain on GTR as support. The standard deviations are represented by vertical lines in each of the
bars and the statistical differences by letters.
Figure 5
Page 22/23Bifactorial analysis of the effect of moisture and temperature on the production of β-glucosidase activity by A.
niger 28A on GTR as support. Moisture of the treatments is presented in (%) and incubation temperature in (°C).
The standard deviations are represented by vertical lines in each of the bars and the statistical differences by
letters.
Figure 6
A) Optimum temperature evaluation for the β-glucosidase activity of partially purified extracts of A. niger 28A
produced by SSF using GTR as support. B) Optimum pH evaluation for the β-glucosidase activity of partially
purified extracts of A. niger 28A produced by SSF using GTR as support The standard deviations are
represented by vertical lines in each of the bars and the statistical differences by letters.
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
GA.png
Page 23/23You can also read

















































