Physiology and regulation of biological rhythms in laboratory animals: an overview
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Laboratory Animals (1989) 23, 107- 125 107
Physiology and regulation of biological rhythms in laboratory
animals: an overview
F. WOLLNIK•
Northwestern University, Department of Neurobiology and Physiology, Evanston, Illinois, USA
Summary cycles per second (e.g. neuronal activity, heart
Biological rhythms have been observed in rate) up to one cycle per year {e.g. seasonal
practically all groups of laboratory mammals rhythms). Although many biological rhythms are
and at every level of physi.ological and be- related to external environmental cycles, such as
havioural organization. Biological rhythms are the daily light-dark cycle or seasonal changes in
classified according to their period as ultradian day length and temperature, most of them persist
( < 24 h), circadian (- 24 h), infradian ( > 24 h), in the laboratory, even under highly standardized
and seasonal or circannual rhythms ( -1 year). conditions. Knowledge of biological rhythms and
This review outlines what is known about the their consideration in experimental design can
neurobiology of biological rhythms in mammals help to reduce the variability of quantitative
and describes the hierarchical order in which measurements in laboratory animals.
ultradian, circadian and infradian rhythms are Over the past 30 years research has been
related to each other. The article does not focused mainly on biological rhythms that are
attempt to catalogue every physiological variable synchronized to the environmental 24-h cycle,
showing rhythmical fluctuations in laboratory and investigators have tried to determine whether
mammals. Rather, it focuses on the basic these rhythms reflect only a passive reaction to
concepts of circadian rhythms and recent advances periodic stimulations from the environment
made in our understanding of the physiology of {exogenous rhythms) or whether they are an
the internal clock controlling circadian and other intrinsic property of the organism {endogenous
biological rhythms. rhythms). Most of the daily fluctuations in
behavioural and physiological variables were
Keywords: Ultradian; Circadian; lnfradian; found to be truly endogenous since they persist
Seasonal, and Circannual Rhythms; Photo- even when no external time cues are present.
periodism However, without a synchronizing 'zeitgeber'
(time giver) the rhythm will slightly deviate from
the 24-h period and eventually 'free-run' with an
Many characteristics of laboratory animals vary intrinsic natural period. Halberg (1959) intro-
more or less regularly with specific periods. duced the name 'circadian' rhythms {circa diem,
These temporal variations are called biological L. ::; about one day) for these endogenous
rhythms and can be observed not only in the rhythms, and the remaining spectrum of biological
whole organism but also in single organs, tissues rhythms has hence been divided into 'ultradian'
or even isolated cells. Biological rhythms cover rhythms with periods shorter than 24 h and
a wide range of frequencies, from one or several 'infradian' rhythms with periods longer than 24 h
(Halberg eta/., 1965). Some seasonal rhythms,
*Present address: Dr Franziska Wollnik, Universitiil Konstanz, as well, have been found to persist under
Fakultiit fiir Biologie, Postfach 55 60, 0-7750 Konstanz I, constant laboratory conditions. Because their
FRO
period is approximately 1 year they are classified
Received 3 March 1988,· accepted 8 July 1988 as 'circannual' rhythms.108 Wollnik
Circadian rhythms
Circadian rhythms are ubiquitous in animals. FREE-RUN U~ER
They have been documented for practically every . CONSTANT DARK !DOl
laboratory species and at every level of organ-
10 -==.. -r=24,5 hr
ization within an organism, from physiological -: CONSTANT LIGHT (LL)
to biochemical and behavioural processes. In ·T =2S,Ohr
mammals, birds, and several invertebrates in-
vestigators have actually been able to find the ENTRAINMENT TO A
ZEITGEBER T=24,0 hr
location of a circadian clock and have discovered
some of the underlying physiological mechanisms
that generate circadian rhythms. PHASE SHIFT
OF ZEITGEBER
=---
Functional properties
To investigate the mechanisms generating cir-
-=
.=ENTRAINMENT TO A
cadian rhythms one could, in principle, study any
= ZEITGEBER T:23.2Shr
behavioural or physiological variable. However, 60
some phenomena have more precise rhythms POSTENTRAINMENT
sm E FREE-RUN
than others and, thus, are more likely to give
reproducible results. Commonly investigated
100 ~
= e :PULSf
.:::::::::::S I IIUUS
':'•
==
-=---
-;:PHASE ADVANCE
circadian rhythms are those of wheel-running
activity or overall locomotor activity of small 110 ~G~E
laboratory animals such as hamsters, rats and
rruce. Spontaneous locomotor activity not only -
- = STIH IA.US =- PHASE DELAY
120
represents the overall physiological status of an
animal (Aschoff, 1962) but it is also a convenient
feature to study, because it can be continuously 0 21. 46
measured for many days using automatic re-
Fig. 1. Idealized dahl from a hypothetical experiment designed
cordin_g equipment with little disturbance to the to show period cllanges of a free-running rhythm caused by
animal. internal (age, hormones) or external Oight, temperature)
Figure 1 shows results of some standard stimuli, entrainment by a strong zeltgeber, and phase shifts
in response to a single pulse stimulus. The data are presented
experimental procedures used to determine as a double plot, a format commonly used for the recordings
functional properties of circadian rhythms. It is of locomotor activity, where the horizontal axis covers a 48-h
based on an idealized experiment originally period and successive days arc plotted from top to bottom.
The black horizontal bars represent the physiological variable
presented by Enright (1981). The data are under investigation, for example wheel-running activity or
presented in a 'double plot' , a format which is food intake. A vertical sequence of these bars represents a
commonly used for recordings of locomotor rhythm with a period of 24 h. A diagonal sequence represents
a free-running rhythm with a period longer or shorter than
activity but could also be applied to other 24 h. The upper portion of the record shows a free-running
behavioural or physiological variables. The first rhythm with periods of r=24·S h and r=lS h. Such a
part illustrates a free-running circadian rhythm lengthening of the free-running period can be observed, for
example, after transferring the animal from continuous dark
after elirrunation of all external time cues. In such (DO) to constant light conditions (LL) o r after changing the
an experiment, the animal would be kept in a hormonal status of the animal. The middle portion shows
soundproof chamber under conditions of con- entrainment to a zeitgeber with a period of T =24 ·0 h (i.e.
a light-dark cycle}, entrainment to a phase shift of the
stant illumination or darkness and constant zeitgeber, and entrainment to a zeitgeber with a period of
temperature, with food and water available ad T =23 · 25 h. The lower portion of the reco.r d shows the effect
libitum. The period of the free-running rhythm of single-pulse stimuli on the circadian rhythm. The first
stimulus caused a phase advance, the second a phase delay.
(r) depends on intrinsic factors, such as the The times at which the phase-shifting stimuli were given are
physiological status of the individual animal indicated by asterisks. (Adapted from Enright, 1981.)Biological rhythms in laboratory animals 109
(e.g. age, hormone levels), and its genetic
background (e.g. diurnal or nocturnal species),
and on external factors, such as environmental 2
conditions (e.g. light intensity, temperature)
and the experimental history of the animal
(e.g. breeding conditions, previous experiments)
(reviewed in: Aschoff, 1979a,b; Pittendrigh ~ or-,-~~~~~----~------~~
& Daan, 1976a). Nonetheless, the internal .,
Cl>
clock is remarkably stable (Pittendrigh & (/)
~ -1
Calderola, 1973). With the exception of light n.
intensity, these factors have only small effects
-2
on the average free-running period. Changes in
light intensity have a dramatic effect on the
free-running period. Under constant darkness 0 6 12 18 24
(DO) T tends to be longer than 24 h in many Circadian time (hours)
diurnal mammals and shorter than 24 h in
nocturnal species. With an increase in light Fig. 2. Phase-response curve (PRC) illustrating the phase-
shifting effe,ct of 60-min light pulses on the wheel-running
intensity under constant light (LL) T lengthens activity rhythm of golden hamsters. The timing of the light
in both diurnal and nocturnal mammals (Aschoff, pulses is expressed as circadian time (CT) relative to activity
1979a,b). onset ( = CT 12). Phase advances are plotted as positive phase
shifts, phase delays as negative phase shifts. Each point
In the presence of a 24-h environmental cycle, represents the mean phase shift of 6 animals; vertical bars
for example a light-dark (LD) regime, the free- indicate ±SD (from Takahashi & Zatz, 1982; ©Copyright
running rhythm adjusts to this zeitgeber and 1982 by the AAAS).
exhibits a period of exactly 24 h. This adjustment
is called ' synchronization' or •entrainment' and
is represented in the second part of Fig. 1. To circadian clock can be entrained by a zeitgeber
verify the entrainment one usually demonstrates is limited. In mammals this range is relatively
that the rhythm follows a phase shift, i.e. a single narrow (20-28 h; Aschoff & Pohl, 1978) and
displacement of the zeitgeber along the time axis. depends on the strength (amplitude) of the
The development of a steady-state (i.e. stable) zeitgeber as well as the strength of the endogenous
phase relationship between the external zeitgeber circadian system of the organism. If the 'range
and the internal rhythm usually requires several of entrainment' is exceeded, the circadian rhythm
cycles. After termination of entrainment the free- _ will free-run with a period close to that observed
run must always start at a reproducible J:!hase rela- under constant conditions. Even then, the
tive to that seen during prior entrainment signals of the zeitgeber can still modulate the
(Enright, 1981). The strongest zeitgeber for most free-running pattern by periodically entraining
animals is the light-dark cycle. Other environ- the rhythm, a phenomenon called 'relative
mental factors that can entrain circadian rhythms coordination'.
are temperature cycles (Aschoff & Tokura, 1986; It should be emphasized that measurements of
Tokura & Aschoff, 1983), food availability circadian rhythms such as locomotor activity or
(Boulos & Terman, 1980), and social cues, which hormonal profiles do not directly represent the
in primates and humans seem to be as important state of the internal pacemaker system, since the
for the entrainment of circadian rhythms as the pacemaker signal can be subject to many internal
light-dark cycle (Wever, 1982). and external modifications. The genuine status
Although entrainment of the free-running of the pacemaker system is described only by the
rhythm is not restricted to periods of exactly period length under free-running conditions
24 h, the range of periods to which the internal and the steady-state phase under entrainment110 Wollnik (Pittendrigh, 1981). In Fig. I, the last part of the Herkenham, 1981) thus providing clues as to data represents a common experiment used to which neural mechanisms might be involved in examine the effects of light or other stimuli, such the generation and entrainment of circadian as hormones, neurotransmitters, and drugs, on rhythms. the circadian clock. If an animal that has been kept in constant darkness is exposed to a light Neuronal organization of biological pulse of 1-60 min, the timing of the activity onset rhythms in mammals will be permanently modified, so that its free- The location of the circadian clock in mammals running rhythm will show a 'phase-shift'. was first discovered by Richter (1967). After Depending on the time of day at which it is removing various endocrine glands and lesioning applied the light pulse may induce an advance specific parts of the rat brain he concluded that or a delay of the activity onset or have no effect a circadian clock must be located near the at all. The time of the light pulse is usually given 'ventral median nucleus' of the hypothalamus. in 'circadian time' relative to the activity onset. Further studies in the J970s determined the In nocturnal species the activity onset is defined primary location more specifically in the supra- as circadian time 12 (CT 12); the animal is active chiasmatic nuclei (SCN) of the hypothalamus during its 'subjective night' (CT 12-24) and is (Moore & Eichler, 1972; Stephan & Zucker, inactive during its 'subjective day' (CT 0-12). 1972a). The magnitude and direction of the phase shift The SCN are bilaterally symmetrical nuclei is plotted in a 'phase response curve' (PRC) as located near the third ventricle and directly above a function of the circadian time at which the the optic chiasm (Fig. 3a). The SCN of rodents stimulus was given. PRCs to light pulses have the have an oval shape, those of cats and monkeys same general shape as shown in Fig. 2 for both are more irregularly shaped (Lydic el a/., 1982). nocturnal and diurnal animals (reviewed in Daan In rats, each nucleus is about 300 J.tffi in & Pittendrigh, 1976). If the light pulse is given transverse diameter and 600 ~-tm in length and during the late subjective night (CT 18- 24) the contains about 8000 neurons organized in rhythm will be advanced; if it is given during the recognizable subpopulations (Van den Pol, early subjective night (CT 12-15) the rhythm will 1980). So far, two specific substances, the be delayed. If the light pulse is given during the peptides vasopressin and vasoactive intestinal subjective day (CT 0-12), little or no phase shifts polypeptide (VIP), have been identified as being occur. The PRC also predicts the repeated daily produced by SCN neurons, although they are adjustments necessary to synchronize the endo- only found in less than half of all neurons genous free-running period (r) to the period (D (Sofroniew & Weindl, 1982). Other neuro- of the external light-dark cycle. Consequently, peptides and neurotransmitters, such as somato- a phase delaying light pulse causes a temporary statin, serotonin, corticotropin-releasing factor, lengthening of the period, and a phase advancing avian pancreatic polypeptide, neuropeptide Y, light pulse causes a temporary shortening of the and gamma-aminobutryc acid (GABA) have period. Under conditions of entrainment the been found in afferent nerve fibres to the SCN daily phase shift A ¢ is equal to the difference (Card & Moore, 1982; Groos, 1982; Sofroniew between the entraining period T and the free- & Weindl, 1982; Van den Pol & Tsujimoto, running period r: t:. ¢ = T- r (Daan & Pitten- 1985). drigh, 1976; Pittendrigh, 1981). The PRC The primary input into the SCN is the approach has been used in mammals to describe retinohypothalamic tract (RHT) through which how light or administration of various drugs and the entraining signals of the external light-dark neurotransmitters can alter the phase and period cycle reach the internal clock (Hendrickson et a/., of the circadian clock (Turek, 1985, 1987; Zatz, 1972; Moore & Lenn, 1972). The RHT is 1979; Zatz & Brownstein, 1979, 1981; Zatz & common to all mammals studied so far. It leaves
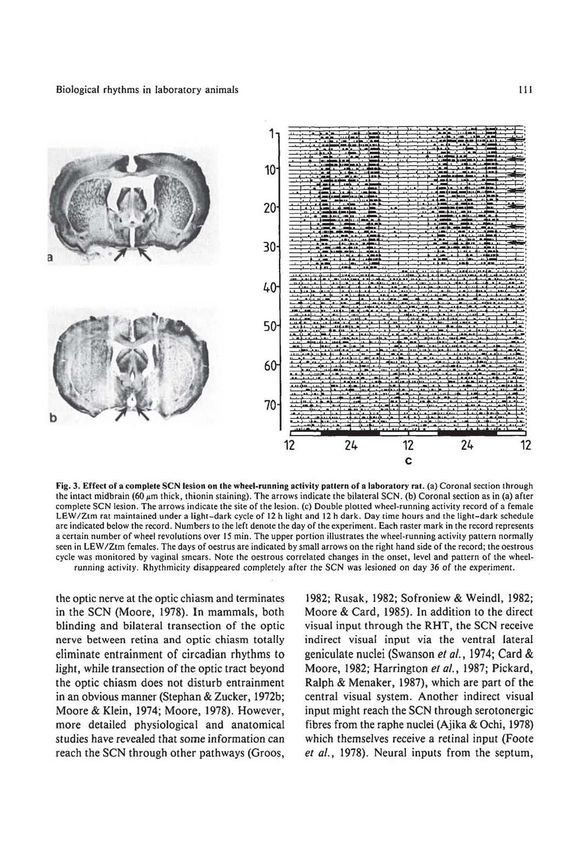
Biological rhythms in laboratory animals Il l
1
10
20
30
4
50
6
70
12 24 12 24 12
c
Fig. 3. Effect of a ~:o mplete SCN lesion on the wheel-ru nning activity pattern of a laboratory rat. (a) Coronal section through
the intact midbrain (60 J.Lm thick, thionin staining). The arrows indicate the bilateral SCN. (b) Coronal section as in (a) after
complete SCN lesi on. The arrows indicate the site of the lesion. (c) Double ploLted wheel-running activity record of a female
LEW/Ztm rat maintained under a light-dark cycle of 12 h light and 12 h dark. Day time hours and the light-dark schedule
are indicated below the record. Numbers to the left denote the day o f the experiment. Each raster mark in the record represents
a certain number of wheel revolutions over 15 min. The upper portion illustrates the wheel-running activity pattern normally
seen in LEW /Ztm females. The days of oestrus are indicated by small arrows on the right hand side of the record; the oestrous
cycle was monitored by vaginal smears. Note the oestrous correlated changes in the onset, level and pattern of the wheel-
running activity. Rhythmicity disappeared completely after the SCN was Jesioned on day 36 of the expe~iment.
the optic nerve at the optic chiasm and terminates 1982; Rusak, 1982; Sofroniew & Weindl, 1982;
in the SCN (Moore, 1978). In mammals, both Moore & Card, 1985). In addition to the direct
blinding and bilateral transection of the optic visual input through the RHT, the SCN receive
nerve between retina and optic chiasm totally indirect visual input via the ventral lateral
eliminate ent rainment of circadian rhythms to geniculate nuclei (Swanson eta/. , 1974; Card &
Jjght, while transection of the optic tract beyond Moore, 1982; Harrington eta/., 1987; Pickard ,
the optic chiasm does not disturb entrainment RaJph & Menaker , 1987), which are part of the
in an obvious manner (Stephan & Zucker , 1972b; central visual system. Another indirect visual
Moore & Klein, 1974; Moore, 1978). However, input might reach the SCN through serotonergic
more detailed physiological and anatomical fibres from the raphe nuclei (Ajika & Ochi, 1978)
studies have reveaJed that some information can which themselves receive a retinal input (Foote
reach the SCN through other pathways (Groos, er a/., 1978). Neural inputs from the septum,112 Wollnik
hypothalamus, thalamus, hippocampus and 1979). This indicates that circadian rhythms in
midbrain as well as commissural connections neuronal activity originate in the SCN. The
between the bilateral SCN have also been neural circadian rhythms in the SCN showed
demonstrated (Pickard, 1982). Efferent characteristic properties (e.g. a free-running
projections of the SCN terminate in other period) expected of an endogenous circadian
hypothalamic nuclei (paraventricular nucleus, osciUator (Inouye & Kawamura, 1982; Inouye,
dorsomedial hypothalamic nucleus), in the 1984). The SCN seem to maintain a
thalamus (paraventricular thalamic nucleus) and neural or hormonal circadian rhythm even
in the midbrain (Swanson & Cowan, 1975; Berk in vitro (Green & Gillette, 1982; Shibata et
& Finkelstein, 1981; Stephan et a/., 1981; a/., 1984; Earnest & Sladek, 1987). Using
Sofroniew & Weindl, 1982). 2-deoxyglucose autoradiography, researchers
The following experiments indicate that the have also found that in a wide range of
SCN play an important role in the normal species glucose consumption follows a cir-
expression of circadian rhythms: cadian rhythm in the SCN (Flood & Gibbs,
(I) SCN lesions: A variety of behavioural and 1982; Schwartz, Reppert, Eagan & Moore-
physiological circadian rhythms, including Ede, 1983). SCN metabolic activity is high
locomotor activity, feeding, drinking, body during the subjective day and low during the
temperature, synthesis and/or secretion of subjective night in both nocturnal and
several hormones, and the sleep-wake cycle, diurnal animals (Schwartz eta!., 1983). In
can be severely disrupted or completely rats the circadian rhythm of glucose uptake
abolished by complete bilateral SCN lesions in the SCN can be detected as early as several
or isolation of the SCN from other neural days before birth, prior to the arrival of any
structures (reviewed in: Moore, 1978, 1983; retinal projections (Reppert & Schwartz,
Rosenwasser & Adler, 1986; Rusak & 1983, 1984).
Zucker, 1979; Turek, 19B5). An example is (3) Electrical and chemical stimulations: In
given in Fig. 3c which illustrates the effect hamsters and rats, short electrical stimul-
of complete SCN lesions on the wheel- ation of the SCN has a similar phase-shifting
running activity pattern of laboratory rats. effect on the circadian rhythm as light
Follow-up studies on rats whose SCN had (Rusak & Groos, 1982). Timed injections of
been lesioned immediately after birth have pharmacological agonists or antagonists of
shown that the function of the SCN were not neurotransmitters found in SCN cells and
taken over by other parts of the brain axons have been shown to mimic or block
(Mosko & Moore, 1978). SCN lesions also the phase-shifting effect of light (reviewed in:
disrupt oestrous cyclicity and photoperiodic Turek, 1985, 1987; Rosenwasser & Adler,
time measurement, two components of the 1986). For example, photic stimulation in-
reproductive process that depend upon the creases the concentration of acetylcholine in
circadian system for temporal information the SCN (Murakami et a/. 1984).
(Rusak & Morin, 1965; Stetson & Watson- Intracerebroventricular injections of car-
Whitmyre, 1976; Turek eta/., 1980). bachol, an acetylcholine agonist, mimic
(2) Demonstration of intrinsic oscillations: Cir- the phase shifting effect of light (Zatz &
cadian rhythms in multi-unit neuronal activity Herkenham, 1"981; Earnest & Turek, 1983),
have been recorded in the SCN and several whereas mecamylamine, a cholinergic antag-
other regions of the rat brain. After surgical onist, blocks the phase-shifting effect of light
isolation of the SCN within a 'hypothalamic (Keefe et a/., J987). This suggests that
island'. circadian firing patterns persist in the acetylcholine may be involved in the trans-
SCN but can no longer be recorded in other mission of light-dark information to and
parts of the brain (Inouye & Kawamura, within the circadian clock.Biological rhythms in laboratory animals 113
(4) SeN-Transplantation: Only recently has the exposing the animals to long-term bright light
pacemaker function of the SCN been demon- conditions or SCN lesions (Rusak, 1977; Earnest
strated in the most direct way. By trans- & Turek, 1982). In other species splitting has also
planting fetal neural tissue containing the been observed after long-term exposure to
anlagen for the SCN into arrhythmic SCN- constant dark (Hoffman, 1971), administration
lesioned hamsters and rats circadian of pharmacological substances (Wirz-Justice,
rhythmicity of locomotor activity has been Groos & Wehr, 1982), and manipulations of the
restored (Kawamura & Nithonmatsu, 1985; endocrine system by gonadectomy and hormone
Aguilar-Roblero e/ a/., 1986; Lehman el at., replacement {Morin, 1980; Morin & Cummings,
1987). It remains to be shown whether 1982). ln hamsters, splitting disappears rapidly,
pacemaker properties such as phase and when the animals are transferred into constant
period of the donor can also be transferred. darkness (Earnest & Turek, 1982). The splitting
This would demonstrate that the trans- phenomenon has led to the assumption that at
planted tissue actually generates the restored least two mutually coupled oscillators underlie
rhythm rather than simply having a permis- the circadian locomotor activity rhythm (Pitten-
sive effect. The specificity of transplanting drigh & Daan, 1976b). Because of the splitting
SCN tissue has been questioned lately after phenomenon and the fact that even the un-
it was observed that circadian rhythms could disturbed, non-split activity rhythms of many
also be restored by transplanting fetal mammalian species display quite complex pat-
cortical tissue into the lesion site (Garcia- terns made up of two or more activity components
Hernandez et a/., 1987). (Aschoff, 1962), it is widely accepted today that
The circadian system of mammals is thought the circadian clock of mammals is a multi-
to consist of multiple circadian oscillators which oscillatory system.
are coupled to each other in both a hierarchical Although multiple circadian oscillators are
and non-hierarchical (mutual) manner (reviewed assumed to lie in the SCN, there is also evidence
in: Pittendrigh, 1960, 1974; Rusak, 1979; Rosen- for the presence of oscillators outside the SCN.
wasser & Adler, 1986). While these oscillators For example, not all circadian rhythms disappear
normally are synchronized in order to generate after complete SCN lesions. In hamsters, both
a single pacemaker signal, certain conditions can circadian and ultradian rhythms in wheel-
cause their desynchronization. running activity have been reported to persist in
The most prominent example of desynchroniz- individual animals (Rusak, 1977). In rats,
ation is 'splitting', the dissociation of the periodic feeding can entrain food-anticipatory
circadian rhythm into two or sometimes even rhythms in a number of physiological parameters
more distinct components (reviewed in: Pitten- (reviewed in: Boulos & Terman, 1980; Aschoff
drigh, 1974; Pittendrigh & Daan, 1976b; Turek et a/., 1983), and those rhythms persist after
eta/., 1982; Rosenwasser & Adler, 1986). A SCN lesions showing the same characteristics as
characteristic feature of splitting is the temporary before (Krieger et a/., 1977; Stephan et at.,
free-run of the two components with di fferent 1979a,b; Stephan, 1982, 1984). These results
periods. The two components often resynchronize indicate that the food entrainable pacemaker is
after reaching a 180° phase-relation and sub- functionally and anatomically independent from
sequently free-run with identical periods. Splitting the 'classical', i.e. light entrainable, circadian
has been reported for a number of mammalian system within the SCN (Gibbs, 1979; Boulos et
species, including hamsters (Earnest & Turek, a/., 1980). The search for the food entrainable
1982; Turek et at., 1982), rats (Boulos & Terman, pacemaker has focused on the ventromedial
1979; Cheung & McCormack, 1983), tree shrews nucleus of the hypothalamus because destruction
(Hoffman, 1971) and monkeys (FuJJer et at., of this nucleus can abolish the food-anticipatory
1979). In hamsters it has been evoked by rhythms (Krieger, 1980; Inouye, 1982). Recently,114 Wollnik
the circadian locomotor actlV!ty rhythm of Hegmann, 1980) as well as in the circadian phase
completely arrhythmic SCN lesioned rats could and period of body temperature (Connolly &
be restored by chronic administration of meth- Lynch, 1981, 1983). Furthermore, a positive
amphetamine in the drinking water (Honma et genetic correlation was found between the phase
a/., 1987). Since the rhythm was obviously not and the period of the circadian rhythms of food
generated by the SCN nor entrained by the light- intake and water consumption, which suggests
dark cycle, one m'\Jst assume that the meth- that both parameters are regulated by a common
amphetamine activated an endogenous oscillatory physiological mechanism and would respond to
mechanism outside the SCN that is probably natural selection as a single circadian complex
identical with the food-anticipatory pacemaker under common gene control (Possidente &
system. Hegmann, 1980). In hamsters, strain differences
have been demonstrated for the free-running
Genetic determination period, the circadian phase of activity onset and
A number of characteristic properties of circadian splitting (Hotz & Turek, 1987). Recently, a
rhythms have been shown to be genetically spontaneous hamster mutant has been isolated
determined. To investigate the genetic back- that has a remarkably short free-running period
ground of circadian rhythms in mammals of only 20 h (Ralph & Menaker, 1987). In rats,
quantitative genetic methods of variance and strain differences have been demonstrated for the
covariance analysis (Hegmann & Possidente, free-running period and the daily pattern of
1981) have been applied to inbred strains of small spontaneous locomotor activity (Buttner &
laboratory animals like mice, hamsters and rats Wollnik, 1984). Two inbred strains of rats,
(Table l). In mice, there are significant dif- BH/Ztm and LEW /Ztm, showed an unusual
ferences between strains in the free-running bimodal and trimodal, respectively, activity
period of wheel-running activity (Ebihara eta/., pattern. A classical genetic analysis revealed that
1978) and food and water intake (Possidente & the trimodal activity pattern of the LEW /Ztm
strain has a recessive single-gene mode of
inheritance (Wollnik eta/., 1987).
Table 1. Inbred strains of laboratory mammals used for
genetic analyses of biological rhythms Another approach to investigate the genetic
background of circadian rhythms is to study
Mice BALB/CBYJ Possidente & Hegmann (1980) strains with known mutations in the visual or
BALB/clbg Connolly & Lynch (1981)
C3H/HeJ Possidente & Hegmann (1980) central nervous system. Mutations which in early
C3H/21bg Connolly & Lynch (1981, 1983) development modify retinal pigment and visual
C57BL/6J Connolly & Lynch (1981, 1983) pathways also modify developmental and func-
Ebihara el a/. (1978)
C57BL/10Sn Ebihara el a/. (1978) tional properties of circadian rhythms. For
Possident e & Hegmann (1980) example, the free-running circadian period of
C58/ J Possidente & Hegmann (1980) both albino and pinkeye-dilute mice is shorter
DBA/IBG Connolly & Lynch (1981)
DBA/IJ Possidente & Hcgmann (1980) than that of pigmented mice (Possidente et a/.,
DDK Ebihara eta/. (1978) 1982). Mice of an anophthalmic strain (ZRDCT-
MOL-A Ebihara eta/. (1978) An) with varying degrees of hypogenesis of the
Rats ACI / Ztm J Biiuner & Wollnik (1984) mediobasal hypothalamus and SCN that showed
LEW/Ztm Wollnik et at. (1987)
a partially or completely arrhythmic wheel-
AS/Ztm ] running activity pattern had less than one third
BH/Ztm Biittner & Wollnik (1984)
l
BS/Ztm of the typical number of cells in the SCN
Golden CB (Scheuch et al., 1982). A strain of laboratory rats
hamsters with a genetic inability to produce vasopressin
LHC
LSH Hotz & Turek (1987)
MHA (Long Evans, Brattleboro strain) showed no
PD4 obvious alterations or deficits in their circadianBiological rhythms in laboratory animals 115
rhymicity (Peterson el a/., 1980) indicating that is often necessary to use elaborate recording and
the circadian clock does not involve the vaso- analysing techniques, such as periodogram and
pressin neurons of the SCN. Inbred strains of power spectrum analyses (Van Cauter, 1981;
laboratory animals with genetically fuced modifi- Wollnik & Dohler, 1986).
cations of the circadian system would provide a Since many ultradian rhythms are related to
good test case for SCN transplantation exper- metabolic processes (e.g. food intake, hormone
iments, since the transfer of characteristic secretion, urinary excretion) and sometimes even
properties of a clock mutant could easily be show an allometric correlation between period and
verified. body weight (Daan & Slopsema, 1978; Gerkema
& Daan, 1985), several authors have suggested
Ultradian rhythms that digestive functions are involved in the
Ultradian rhythms are defined as biological generation of ultradian rhythms (reviewed in:
rhythms with a period significantly shorter than Aschoff, 1962; Daan & Aschoff, 1981). However,
24 h, ranging from 1 to 12 h (Halberg et a/., it is now well established that their most likely
1965). Typical examples of ultradian rhythms are site of generation is the nervous system (reviewed
the repetition of rapid eye movements (REM) in Schulz & Lavie, 1985). Two general models
every 90 min during sleep in humans and some for the generation of ultradian rhythms have
mammalian species and ultradian rhythms in been proposed. The first assumes the existence
physiological parameters associated with REM of an ultradian pacemaker system that is
sleep (reviewed in Schulz & Lavie, 1985). Other anatomically and functionally independent from
examples are episodic secretion patterns of the circadian system. It was developed by
various hormones, such as luteinizing hormone Kleitman (1961, 1982) and is based on data on
(peaks every 0· 5-3 h), growth hormone (peaks the 90-min sleep-wake cycle in humans. This
every 3 h), and corticosterone (reviewed in Van independent ultradian pacemaker system can
Cauter & Honinckx, 1985), and short-term either be a periodic oscillatory system or a
behavioural rhythms in locomotor activity or homeostatic renewal process (Gerkema & Daan,
food intake of small mammals (Gerkema & 1985). The second model views ultradian rhythms
Daan, 1985; Honma & Honma, 1985a). Ultradian as a result of desynchronization of several
rhythms are more difficult to investigate than circadian oscillators that are coupled to each
circadian rhythms for the following reasons. other in different phase relations (Pittendrigh,
First, ultradian rhythms include a wide range of 1974). Considering the broad variety of ultradian
rhythmic phenomena which do not necessarily rhythms it is not surprising that results have been
share common properties. For example, some reported in support of both models.
ultradian rhythms are true periodic processes For example, an independent ultradian oscil-
with a consistent period, whereas others are lator appears to generate the feeding and
rather episodic with variable time lags between locomotor activity rhythms in the common vole
single events (Aschoff & Gerkema, 1985). (Daan & Slopsema, 1978; Daan & Aschoff,
Second, unlike circadian rhythms, which have 1981). In this species a total lesion of the SCN
evolved in adaptation to the 24-h fluctuations of completely abolished the circadian rhythm but
the environment, ultradian rhythms do not did not affect the ultradian rhythmicity (Gerkema
correspond to any known physical cycle in the & Daan, 1985). Similarly, complete SCN lesions
environment, and a general functional significant in rats did not disrupt the pulsatile pattern of LH
like that of circadian rhythms has not been release (Soper & Weick, 1980) suggesting that
demonstrated yet (Gerkema & Daan, 1985). ultradian and circadian rhythms in hormone
Third, in order to detect ultradian rhythms in release are generated by different neural centres.
raw data and to discriminate truly rhythmic In both cases, investigators identified the arcuate
phenomena from random stochastic processes it nucleus (ARC) of the hypothalamus as a possible116 Wollnik candidate for the ultradian pacemaker system ultradian rhythms in the adult LEW /Ztm rats (Gerkema & Daan, 1985; Knobil & Hotchkiss, depends on the hormonal status of the animals; 1985). The same neuronal site has also been high serum levels of oestrogen suppress ultradian suggested as the location of a pacemaker for rhythms in both male and female rats (Woilnik ultradian rhythms in growth hormone release & Dohler, 1986). Since similar effects of hor- in rats (Eikelboom & Tannenbaum, 1983). mones on the coupling between circadian oscil- However, even if ultradian rhythms are gen- lators have been demonstrated for hamsters erated by an independent ultradian pacemaker (Morin, 1980) and mice (Daan et a/., 1975), they can still be modulated by the circadian ultradian rhythms in locomotor activity may be system. Such a modifying influence of the the result of the combination of several circadian circadian system has been found in the pulsatile oscillators coupled to each other in various phase patterns of hormone release (reviewed in Turek relations. & Van Cauter, 1988) and the behavioural The diversity of ultradian rhythms is also ultradian rhythms of voles (Gerkema & Daan, apparent in the domain of their functional 1985). relevance. For example, the normal reproductive Behavioural activity patterns of small labor- function of humans and primates depends on a atory mammals that show short-term fluctuations specific ultradian timing of the release of are also often referred to as ultradian rhythms gonadotropin-releasing hormone by the hypo- (reviewed in: Aschoff, 1962; Daan & Aschoff, thalamus and luteinizing hormone by the pituitary 1981). In laboratory rats, behavioural ultradian (Knobil & Hotchkiss, 1985). In voles, the rhythms appear prior to circadian rhythms ultradian feeding and activity rhythms are during early developmental stages (Honma & essential for providing optimal timing of food Honma, l985b). They are also prominent during intake and digestive pauses as well as inter- aging (Albers eta/., l981a) and under long-term individual social synchrony among conspecifics exposure to constant light (Albers eta/., 1981a; in order to reduce the predator risk for the Honma & Honma, I985a) when the circadian individual animal (Daan & Slopsema, 1978; rhythmicity fades or disappears. These findings Daan & Aschoff, 1981). have even led to the suggestion that ultradian Ultradian rhythms often have periods of 3, 4, rhythms may be the underlying time pattern from 4·8, or 6 h. Since these periods are subharmonics which circadian rhythms are then derived by of the circadian period, one must verify that frequency and ampUtude modulation (Honma & they are clearly detectable in the raw data and Honma, 1985a,b). not mere artifacts of the statistical method The following observations on behavioural used for data analysis. The predominance of ultradian rhythms in laboratory rats, however, integer ratios of the circadian period can be do not support the existence of an independent interpreted both as a result of the desynchroniz- ultradian pacemaker. First, ultradian activity ation of a multi-oscillatory circadian pacemaker rhythms in adult laboratory rats have so far been system and as a sign for independent ultra- described only for particular animals (Stephan, dian oscillators that have evolved during the 1983) or specific inbred strains, e.g. LEW /Ztm evolutionary process because they could maintain (Buttner & Wollnik, 1984), and may thus not be a constant phase relationship with the circadian characteristic for the normal temporal organiz- rhythm (Broughton, 1985). Organisms with ultra- ation of rat behaviour. Second, since complete dian rhythms of 3, 4, 4 · 8 or 6 h periods should SCN lesions in the inbred strain LEW /Ztm have significant advantages over organisms with abolished both ultradian and circadian rhythms ultradian rhythms of, for example, I· 7, 5 or 7 h of wheel-running activity, the SCN is apparently periods, since the latter would never stay in phase involved in the control of both rhythms (Wollnik with each other and the major environmental & Turek, 1987). Third, the expression of periodicities.
Biological rhythms in laboratory animals 117
Although there is no doubt today that many phase. On the days following ovulation, activity
behavioural and physiological events follow an starts later and shows a bi- or trimodal pattern.
ultradian temporal pattern, no other class of By looking at continuous recordings of locomotor
biological rhythms is characterized by such a activity one can easily predict the day of
diversity of rhythmic phenomena, generating ovulation. Changes in the onset, intensity and
mechanisms, and biological functions. There- pattern of wheel-running activity are almost as
fore, caution is warranted when results from reliable a marker of the oestrous cycle as vaginal
observations of one ultradian phenomenon are smears (Sridaran & McCormack, 1977).
being generalized to other ultradian rhythms. The oestrous cycle is an endogenous rhythm
that depends on various components of the
lnfradian rhythms neuroendocrine system located in the ventro-
Infradian rhythms are defined as biological medial arcuate region and the preoptic supra-
rhythms with a period significantly longer than chiasmatic area of the hypothalamus (reviewed
1 day, but less than 1 year (Halbert eta/., 1965). in Mahesh, 1985). A striking feature of the
In mammals, many infradian rhythms are related oestrous cycle is the circadian timing of many
to functional changes in the ovary and can oestrous related events. For example, ovulation
therefore be observed only in females of spon- and the LH surge on the day of proestrus
taneous ovulatory species. Oestrous is induced maintain a constant phase relation with the
by changes in the levels of the pituitary hormones external light-dark cycle and follow a phase shift
luteinizing hormone (LH) and follicle-stimulating of this zeitgeber (Alleva et al., 1968; Stetson &
hormone (FSH) (reviewed in Mahesh, 1985) and Gibson, 1977; Gallo, 1981). Even in the absence
occurs at regular intervals characteristic for each of external time cues, the oestrous cycle is closely
species (mice, rats, hamsters: once every 4-5 associated with the circadian system. For example,
days, guineapigs, sheep, goats: once every 13-20 the oestrous cycle of hamsters and rats free-runs
days). The resulting physiological changes in the with a period 4 or (in some rats) 5 times that of
ovaries include fluctuations of oestrogen and the circadian activity rhythm (Fitzgerald &
progesterone secretion which in turn influence Zucker, 1976; Albers et a/., 198lb). Female
behavioural parameters such as spontaneous hamsters whose activity rhythm was split after
locomotor activity, food intake, aggression and exposure to constant light show two LH surges,
sexual behaviour (reviewed in Campbell & each occurring 0-4 h before the onset of one
Turek, 1981}. activity component (Swann & Turek, 1985).
The period of behavioural heat on or around The circadian timing of the oestrous cycle
the day of ovulation is characterized by an seems to be controlled by the SCN. Bilateral
increased level of activity (rats: Albers et at., lesions of the SCN not only abolish the
1981 b, mice: Guttman et a!., 1975, hamsters: preovulatory LH surge and thus impede ovulation
Finkelstein eta/., 1978). In hamsters and rats, but also cause a state of persistent oestrous in
activity starts earlier on the day of oestrous hamsters and rats (Stetson & Watson-Whitmyre,
resulting in a 'scalloping' of the activity onset in 1976). Since the preovulatory LH surge is
intervals of 4 or 5 days (Albers et a/., 1981 b; normally triggered by an increased secretion of
Morin et a/., 1977; Sridaran & McCormack, gonadotropin releasing hormone (GnRH) from
1977). Furthermore, the activity pattern under- the mediobasal hypothalamus (Levine & Ramirez,
goes complex changes during the oestrous cycle 1982) the SCN must somehow be coupled to
(Takahashi & Menaker, 1980). Figure 3c shows an those GnRH neurons. However, it is not clear
example of oestrous-correlated modulation of yet which pathways and neurotransmitters are
wheel-running activity in a laboratory rat. On the involved in the transmission of the circadian
day of ovulation activity starts earlier and retains timing signal to the mediobasal hypothalamus
a constant high level throughout the activity (reviewed in: Turek & Van Cauter, 1988).118 Wollnik
Except those infradian rhythms associated manipulated using different light- dark cycles
with the oestrous cycle only few others have been (reviewed in: Hoffmann, 1981; Keefe & Turek,
reported. For example, in dormice kept under 1985). Much of our understanding of the
various laboratory conditions infradian rhythms neuroendocrine events involved in the photo-
with a period of about 2 months have been periodic control of reproduction is derived from
demonstrated for several physiological functions, studies on three mammalian species: the golden
such as body weight, food and water intake, and hamster and the Djungarian hamster (both long-
weight of liver, testis, and adrenal and sal vary day breeders) and the sheep (a short-day
gland (Mrosovsky eta/., 1980). Another example breeder). In sheep, long photoperiods (more than
is the weekly, or circaseptadian, rhythm of 13 h light per day) induce reproductive quiescence,
enzyme activity in the rat pineal (Vollrath, 1982). whereas short photoperiods (less than 12 h light
Since this weekly rhythm was only found in per day) stimulate gonadal reactivation and
animals that had been kept under normal reproduction; in hamsters the effect is just the
laboratory conditions and had not been demon- opposite. However, the photoperiod induced
strated to free-run with a period of about 7 days, changes are not permanent. In hamsters, for
it is likely that it is an example of an exogenous examples, the short-day induced gonadal quies-
rhythm induced by periodic events in the cence persists only for about 18 weeks. After 18
laboratory environment, such as work schedules weeks gonadal reactivation may occur even if the
of animal care personnel, heater operation, street animal remains exposed to short days (Turek eta/.,
noise, etc. 1975). Before gonadal quiescence can be induced
by short photoperiods again, the hamsters would
Seasonal and circannual rhythms have to be kept under long day conditions for
Many biological activities and reproductive several weeks (Stetson et a/., 1977).
functio~s are restricted by environmental con- Photoperiodic animals can measure the length
ditions such as temperature, day length, or food of day very precisely. In early work on the
availability to a time of the year when they are mechanisms of photoperiodic time measurement
most likely to be successful. In addition, many it was assumed that animals determine the total
behavioural, physiological and morphological duration of light and darkness by some sort of
processes undergo annual fluctuations that are ' hourglass' mechanism or interval timer (Farner
directly or indirectly related to seasonal variations et al.,, 1953). Since then several experiments using
in the environment (reviewed in Gwinner, 1981). unu~uallight-dark cycles ('resonance', 'T' and
These seasonal rhythms may be referred to as 'night interrupt' experiments) have revealed, that
'circannual' rhythms only if it is has been animals measure the length of day by comparing
demonstrated that they persist with a period the external light-dark cycle against an endo-
of about 1 year for at least two cycles under genous circadian rhythm of light sensitivity
constant environmental conditions. Annual (reviewed in Keefe & Turek, 1985). In those
rhythms of body weight or reproductive func- experiments light was presented only for short
tions that can be observed in many mammals periods under otherwise constant darkness. The
under natural conditions have been confirmed animal interpreted as either long or short days
as a circannual rhythm only in a few species, such only those light pulses that were applied during
as ground squirrels (Pengelty & Asmundson, a certain phase of the circadian photosensitivity
1974). rhythm.
In many seasonal breeding animals, seasonal An important part of the photoperiodic time
changes in reproductive functions are not ob- measurement seems to be the circadian system
served under constant laboratory conditions. in the SCN. Bilateral lesions of the SCN abolish
Their reproductive state is a direct result of the photoperiod induced changes in the reproductive
day length, or 'photoperiod', and thus can be status; lesioned animals retain their gonadalBiological rhythms in laboratory animals 119
function regardless of the light-dark cycle two reasons. First, they allow an optimal timing
(hamsters: Rusak & Morin, 1976; Stetson & of biological activities within the context of
Watson-Whitmyre, 1976; ewes: Domanski eta/., physical environmental cycles. Circadian and
1980). The neural information about day length circannual rhythms serve as biological clocks that
is transferred to the endocrine hypothalamic- enable the organism to anticipate periodic events
pituitary-gonadal axis via t he pineal organ in its environment so that it can initiate slow
(Reiter, 1980a,b). Using anatomical and physio- processes on time. Second, biological rhythms
logical techniques the following pathway between provide an internal framework for the temporal
the SCN and the pineal organ has been discovered organization of physiology and behaviour thus
(reviewed in Keefe & Turek, 1985). SCN efferents contributing to the maintenance of a balanced
project to regions of the paraventricular nuclei internal milieu.
of the hypothalamus (Klein et a/., 1983; Biological rhythms are classified according to
Pickard & Turek, 1983; Lehman et at., 1984; their period as circadian, ultradian, infradian,
Inouye & Turek, 1986). The paraventricular and seasonal or circannual rhythms. In recent
nuclei project to the intermediolateral nuclei of years, researchers have discovered some of the
the spinal cord (Swanson & Kuypers, 1980) which underlying pacemaker systems and physiological
innervate the superior cervical ganglia (Rando et mechanisms involved in the generation, entrain-
at., 1981). A beta-adrenergic input from the ment and coupling of biological rhythms .
superior cervical ganglia controls the activity of Circadian rhythms are generated by a multi-
pineal N-acetyltransferase, thereby regulating the oscillator system located in the SCN and are
melatonin synthesis in the pineal organ (Zatz, synchronized to the external24 h light-dark cycle.
1981; Klein, 1985). In a number of photoperiodic Ultradian rhythms can be generated by a variety
species, pinealectomy prevents the short-day of different neural pacemaker systems and are
induced gonadal regression (reviewed in Hoffman, characterized by the diversity of their rhythmic
1981). phenomena. Many infradian rhythms are the
The SCN regulates the rhythmic timing of result of changes in gonadal hormone levels
production of melatonin so that melatonin levels during the oestrous cycle and can only be
are high at night and low during the day observed in female animals. Many annual
(Goldman, 1983; Goldman & Darrow, 1983). rhythms are correlated with changes in the
Recent studies on both Djungarian hamsters and reproductive functions over the year. Photo-
sheep suggest that the duration of high night-time periodic changes of reproductive functions are
levels in circulating melatonin determines whether controlled through complex interactions between
the photoperiod will be interpreted as a long or the SCN, the pineal, and the hypothalamic-
a short day (Carter & Goldman, 1983a,b; pituitary-gonadal axis. Frequency and amplitude
Bittman & Karsch, 1984). It is not known yet of ultradian rhythms are often modulated on a
how melatonin alters hypothalamic-pituitary circadian time scale. The circadian system must
functions and how the signal of melatonin be considered the most important biological
duration is transformed into a change in clock since it affects the regulation of all other
neuroendocrine gonadal activity. biological rhythms. Many key events within the
oestrous cycle are timed by the circadian system,
Conclusion and the photoperiodic control of seasonal
Biological rhythms have been observed in rhythms involves the circadian clock for
practically all groups of laboratory animals and measuring day length.
at every level of physiological and behavioural Most, if not all, physiological systems are
organization even under standardized laboratory affected by biological rhythms, and laboratory
conditions. Biological rhythms are important for animal science should take this into account in
the health and wellbeing of the organism for designing experimental schedules. For example,120 Wollnik
contradictory results from different laboratories of methodological or other constraints, exper-
might be due simply to differences in the daily imenters should at least provide information
schedules in which experiments are being per- about their experimental time schedule and the
formed. Different findings on the periodicity of light-dark cycle of their animals in order to allow
ultradian rhythms, such as episodic hormonal others to reproduce or compare their results.
fluctuations, might be due to different sampling
intervals. Such pitfalls can be avoided by Acknowledgments
scanning the unknown rhythmicity of the in- I am grateful to Professors Fred W Turek and
vestigated variable with repeated experiments Joe S Takahashi for their continuing en-
performed at several time points in the cycle of couragement and helpful advice. My thanks also
a rhythm in order to determine characteristic go to Andreas Herrmann, Amy Jetton, Janet E
phase points, such as the daily peak or low of Joy, Jill J Milette, RickS Smith, and Beth E F
a circadian rhythm. These should serve as Wee for their suggestions and comments on an
reference points when selecting the times to earlier draft of this manuscript. Preparation
perform the real experiments. If this chrono- of this article was supported by Deutsche
biological approach cannot be taken because Forschungsgemeinschaft (11 02-WO 354/2-1).
References
Aguilar-Roblero R, Garia-Hernandez F, Aguilar R, Aschoff J & Tokura H (1986) Circadian activity rhythms in
Arankowsky-Sandoval G & Drucker-Colin R (1986) Supra- squirrel monkeys: entrainment by temperature cycles.
chiasmatic nucleus transplants function as an endogenous Journal of Biological Rhythms 1, 91-99
oscillator only in constant darkness. Neuroscience Lerters Berk ML & Finkelstein JA (1981) An autoradiographic
69, 47-52 determination of the efferent projections of the suprachias-
Ajika K & Ochi J (1978) Serotonergic projections to the matic nucleus of the hypothalamus. Brain Research 226, 1-13
suprachiasmatic nucleus and the median eminence of the Bittman EL & Karsch FJ (1984) Nightly duration of pineal
rat: identification by fluorescence and electron microscope. melatonin secretion determines the reproductive response
Journal of Anatomy 127, 563-576 to inhibitory day length in the ewe. Biology of Reproduction
Albers HE, Gerall AA & Axelson JF {198la) Circadian 30, 558-593
rhythm dissociation in the rat: effects of long-term constant Boulos Z, Rosenwasser A & Terman M (1980) Feeding
illumination. Neuroscience Lerters 25, 89-94 schedules and the circadian organization of behavior in
Albers HE, Gerall AA & Axelson JF (198lb) Effect of the rat. Behavioral Brain Research 1, 39-65
reproductive state on circadian periodicity in the rat. Boulos Z & Terman M (1979) Splitting of circadian rhythms
Physiology & Behavior 26, 21-25 in the rat. Journal ofComparative Physiology A 134,75-83
Alleva JJ, Waleski MY, Alleva FR & Umberger EJ (1968) Boulos Z & Terman M (1980) Food availability and daily
Synchronizing effect of photoperiodicity on ovulation in biological rhythms . Neuroscience & Biobehavioral Reviews
hamsters. Endocrinology 82, 1227-1235 4, 119-131
Aschoff J (1962) Spontane lokomotorische Aktivitii.t. In Broughton RJ (1985) Three central issues concerning
Handbuch der Zoologie (eds JG Helmcke, H von Lengerken ultradian rhythms. In Ultradian Rhythms in Behavior and
& D Starck), vol. 8, pp. 1-76. Berlin: Walter de Gruyter Physiology (eds H Schulz & P Lavie), pp. 217-233. Berlin:
Aschoff J (1979a) Circadian rhythms: influences of internal Springer Verlag
and external factors on the period measured in constant Buttner D & Wollnik F (1984) Strain-differentiated circadian
conditions. Zeitschrift fur Tierpsychologie 49, 225-249 and ultradian rhythms in locomotor activity of the
Aschoff J (1979b) Circadian rhythms: general features and laboratory rat. Behavior Genetics 14, 137-152
endocrinological aspects. In Endocrine Rhythms (ed D Campbell CS & Turek FW (1981) Cyclic function of the
Krieger), pp. l-61, New York: Raven Press mammalian ovary. In Handbook of Behavioral Neuro·
Aschoff J & Gerkema MP (1985) On diversity and uniformity biology, Vol. 4: Biological Rhythms (ed. J Aschoff),
of ultradian rhythms. In Ultradian Rhythms in Behavior pp. 523-545. New York: Plenum Press
and Physiology (eds H Schulz & P Lavie), pp. 321-334. Card JP & Moore RY (1982) Ventral lateral geniculate nucleus
Berlin: Springer Verlag efferents to the rat suprachiasmatic nucleus exhibit avian
Aschoff J, von Goetz C & Honma Kl (1983) Restricted pancreatic polypeptide-like immunoreactivity. Journal of
feeding in rats: effect of varying feeding cycles. Zeitschrift Comparative Neurology 206, 390-396
fiir Tierpsychologie 63, 91-111 Carter DS & Goldman BD (1983a) Antigonadal effects of
Aschoff J & Pohl H (1978) Phase relations between a timed melatonin infusion in pinealectomized male Djun-
circadian rhythm and its zeitgebers within the range of garian hamsters (Phodopus sungorus sungorus): Duration
entrainment . Naturwissenschaften 65, 80-84 is the critical parameter. Endocrinology 113, 1261-1267Biological rhythms in laboratory animals 121
Carter OS & Goldman BD (1983b) Progonadal role o f the Foote WE, Tabor-Pierce E & Edwards L (1978) Evidence
pineal in the Djungarian hamster (Phodopus sungorus for a retinal projection to the midbrain raphe of the cat.
sungorus): Mediation by melatonin. Endocrinology 113, Brain Research 156, 135-140
1268-1273 Fuller CA, Sulzman FM & Moore-Ede MC (1979) Circadian
Cheung PW & McCormack CE (1983) Splitting of the loco- control of thermoregulation in the squirrel monkey, Saimiri
motor activity rhythm in rats by exposure to continuous sciureus. American Journal of Physiology 236, RI53-RI61
light. American Journal of Physiology 244, R573-R576 Gallo RV (1981) Pulsatile release during the ovulatory LH surge
Connolly MS & Lynch CB (1981) Circadian variation of strain on proestrus in the rat. Biology of Reproduction 24, 100-104
differences in body temperature and activity in mice. Garcia-Hernandez F, Aguilar-Roblero R & Drucker-CoUn
Physiology & Behavior 27, 1045- 1049 R (1987) Transplantation of the fetal occipital cortex to
Connolly MS & Lynch CB (1983) Classical genetic analysis the third ventricle of SCN-Iesioncd rats induces a diurnal
of circadian body temperature rhyt hms in mice. Behavior rhythm in drinking behavior. Brain Research 418, 193-197
Genetics 13, 491-500 Gcrkema MP & Daan S (1985) Ultradian rhythms in behavior:
Daan S & Aschoff J (1981) Shon-term rhythms in activity. The case of the common vole (Microtus arvatis). In
In Handbook of Behavioral Neurobiology, Vol. 4: Bio- Ultradian Rhythms in Physiology and Behavior (eds H
logical Rhythms (ed. J. Aschoft), pp. 491-522. New York: Schulz & P Lavie), pp. 11-31. Berlin: Springer Verlag
Plenum Press Gibbs FP (1979) Fixed interval feeding does not entrain the
Daan S, Damassa D, Pittendrigh CS & SmithER (1975) An circadian pacemaker in blind rats. American Journal of
effect of castration and testosterone replacement on a Physiology 236, R249-R2S3
circadian pacemaker in mice (Mus musculus). Proceedings Goldman BD (1983) The physiology of melatonin in
oftheNationalAcademyofSciences, USA 12,3744-3747 mammals. In Pineal Research Reviews (ed RJ Reiter),
Daan S & Pittendrigh CS (1976) A functional analysis of vol. I, pp. 145-182. New York: Alan R. Liss
circadian pacemakers in nocturnal rodents. II. The Goldman BD & Darrow JM (1983) The pineal gland and
variability of phase response curves. Journal of Com- mammalian photoperiodism. Neuroendocrinology 37,
parative Physiology A 106, 253-266 386-396
Daan S & Slopsema S (1978) Short-term rhythms in foraging Green OJ & Gillette R (1982) Circadian rhythm of firing rate
behavior of the common vole, MicrOtus arvalis. Journal recorded from single cells in the rat suprachiasmatic brain
of Comparative Physiology A 127, 215-227 slice. Brain Research 245, 198-200
Domanski E, Pnekop F & Polkowska 1 (1980) Hypothalamic Groos GA (1982) The neurophysiology of the mammalian
centres involved in the control of gonadotropin secretion. suprachiasmatic nucleus and its visual afferents. In
Journal of Reproduction and Fertility 58, 493-499 Vertebrate Circadian Rhythms: Structure and Physiology
Earnest DJ & Sladek CD (1987) Circadian vasopressin release (eds J Aschoff, S Daan & GA Groos), pp. 96-105. Berlin:
from perfused rat suprachiasmatic cxplants i.n vitro: effects Springer Verlag
of acute stimulation. Brain Research 422, 398-402 Guttman R, Lieblich I & Gross R (1975) Behavioral correlates
Earnest OJ & Turek FW (1982) Splitting of the circadian of estrous cycle stages in laboratory mice. Behavioral
rhythm of activity in hamsters: Effects of exposure to Biology 13, 127-132
constant darkness and subsequent re-exposure to constant Gwinner E (1981) C ircannual systems. In Handbook of
light. Journal of Comparative Physiology A 145,405-411 Behavioral Neurobiology, Vol. 4: Biological Rhythms (ed
Earnest OJ & Turek FW (1983) Ro:Je for acetylcholine in J Aschoff), pp. 391-410. New York: Plenum Press
mediatingeffectsofllghton reproduction. Science219, 77-79 Halberg F (1959) Physiological 24-hour periodicity: general
Ebihara S, Tsuji K & Kondo K (1978) Strain differences of and procedural considerations with reference to the
the mouse's free-running circadian rhythm in continuous adrenal cycle. Zeitschrift fiir Vitamin-. Horman- und
darkness. Physiology Behavior 20, 795-799 Fermentforschung 10, 225-296
Enright JT (1981) Methodology. In Handbook of Behavioral Halberg F, Engeli M, Hamburger C & Hillman VD (1965)
Neurobiology, Vol. 4: Biological Rhythms (ed. J. Aschoff), Spectral resolution of low-frequency, small-amplitude
pp. 11-19. New York: Plenum Press rhythms in excreted 17-ketosteroids: probably androgen-
Eikelboom R & Tannenbaum GS (1983) Effects of obesity induced circaseptan desynchronization. Acta Endocrinologica
inducing ventromedial hypothalamic lesions on pulsatile 103, 1-54
growth hormone and insulin secretion: evidence for the Harrington ME, Nance DM & Rusak B (1987) Double-
existence of a growth hormone releasing factor. Endo- labeling of neuropeptidc Y-immunoreactive neurons which
crinology 112, 212-219 project from the geniculate to the suprachiasmatic nuclei .
Farner DS, Mewaldt LR & Irving SO (1953) The roles of Brain Research 410, 275-282
darkness and light in the activation of avian gonads. Hegmann JP & Possidente B ( 1981) Estimating genetic
Science 118, 351-352 correlations from inbred strains. Behavior Genelics 11,
Finkelstein JS, Baum FR & Campbell CS (1978) Entrainment 103-114
of the female hamster to reversed. photoperiod: Role of Hendrickson AE, Wagoner N & Cowan WM {1972)
the pineal. Physiology & Behavior 21, 105-111 An autoradiographic and electron microscopic study
Fitzgerald KM & Zucker I (1976) Circadian organization of of the retina-hypothalamic connections. Zeitschrift fiir
the estrous cycle of the golden hamster. Proceedings of Zel/forschung 135, 1-26
the National Academy of Sciences, USA 13, 2923-2927 Hoffmann K (1971) Splitting of the circadian rhythm as a
Flood DG & Gibbs FP (1982) Species differences in function of tight intensity. In Biochronometry (ed. M
[ 14C]2-deoxyglucose uptake by suprachiasmatic nuclei. Menaker), pp. 134-150. Washington DC: National Academy
Brain Research 232, 200-205 of SciencesYou can also read

















































