Occurrence of depredation by common bottlenose dolphins (Tursiops truncatus) on reef fish captured and released by rod and reel fisheries in the ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Occurrence of depredation by
common bottlenose dolphins (Tursiops truncatus)
on reef fish captured and released by rod and reel fisheries
in the northeastern Gulf of Mexico
by
Corie E. Grewal
Advisor: Dr. Andrew J. Read
Prepared for: Jessica Powell, National Marine Fisheries Service
April 30, 2021
Masters project submitted in partial fulfillment of the requirements for the Master of
Environmental Management degree in the Nicholas School of the Environment of Duke
University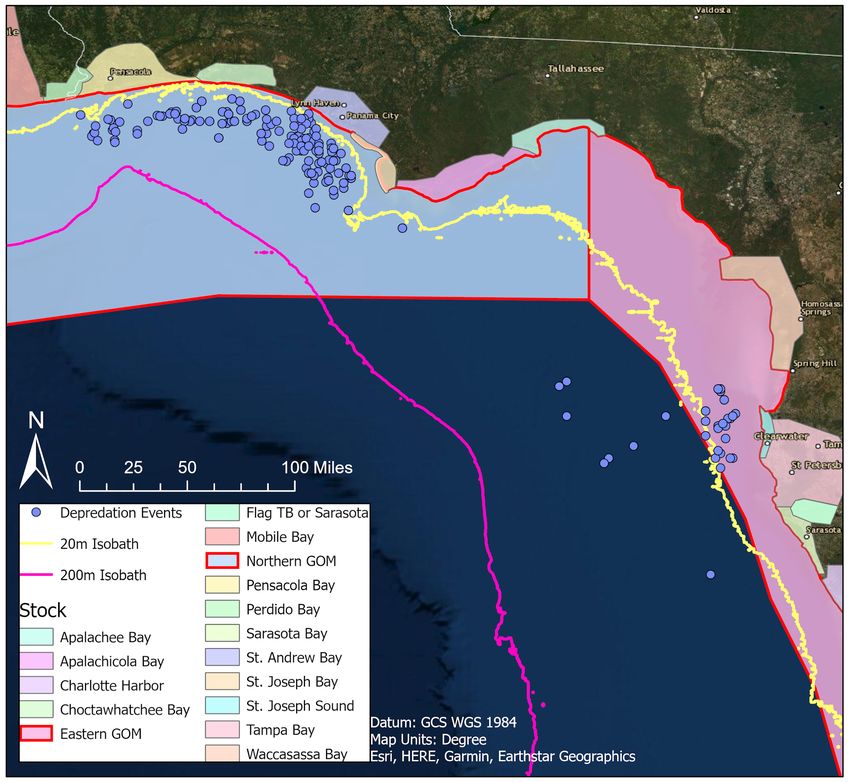
Executive Summary
In the Gulf of Mexico, common bottlenose dolphins (Tursiops truncatus) have been
observed depredating fish from rod and reel anglers. Depredation is defined as the removal of
captured fish or bait by a predator, in this case bottlenose dolphins. Depredation can cause
serious injury and mortality for dolphins who may become entangled or hooked in fishing gear
and also increases the costs of fishing for anglers by forcing them to replace gear and bait, as
well as the loss of catch to dolphins. Protected species managers in the Gulf are receiving an
increasing number of reports about depredation from anglers and are hoping to gain more insight
into how such interactions can be reduced.
To better understand the nature and extent of these interactions, I examined potential
factors that could influence the probability of depredation and the spatial distribution and
frequency of such interactions. I analyzed data collected from the Florida Fish and Wildlife
Conservation Commission (FWC) fisheries-dependent monitoring at-sea observer program to
determine which factors played a role in rod and reel dolphin depredation of rod and reel
fisheries between 2009 and 2020. Every observation in the dataset represented a fish caught by a
rod and reel angler, and depredation was recorded in the dataset after fish were discarded and
observed being consumed by a dolphin. I chose potential predictive factors based on their known
or hypothesized influence on dolphin behavior and used a generalized linear model (GLM) to
examine relationships between instances of depredation and these predictive factors. The
variables that I examined included year, depth, geographic location binned by latitude and
longitude into fishing zones, vessel type, fishing mode, the number of anglers fishing, taxonomic
family of captured fish, fish fork length and whether or not the fish was vented. To reduce
dependence between individual observations that may have taken place on the same trip or at the
same location, I also created and included a lag variable.
The most parsimonious model was the best fit to the data, and included year, the number
of anglers fishing, two geographic areas surrounding Panama City and Destin, Florida, and the
fish families Lutjanidae (snapper family), Carangidae (jack family), Serranidae (grouper and sea
bass family), and a binned group of other species that were individually captured less frequently.
All of these variables were significant predictive factors in the model.
The results suggest that food provisioning in the Florida panhandle is a driver of
depredation and that the incidence of depredation has increased over time. A spatial analysis of
i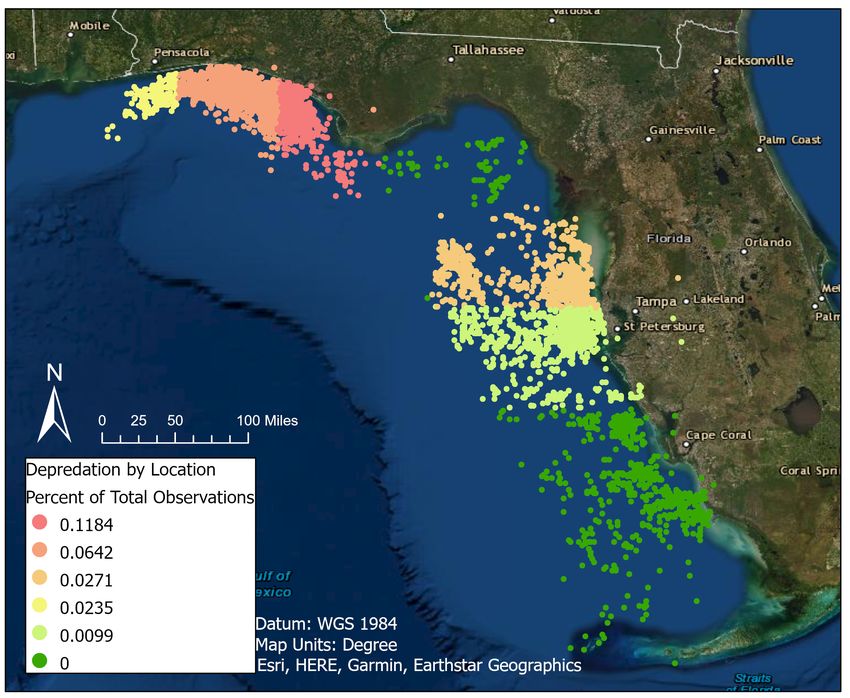
the location of depredation events indicates that the northern and eastern coastal stocks, which
occur over the near-shore shelf off the Florida gulf coast, have become conditioned to exhibit a
range of low cost foraging techniques, such as scavenging, begging, and depredation. Snappers
were observed in 78% of depredation events and this family was the most significant predictive
factor in the model, revealing a regional prey preference for snapper by dolphins of the northern
coastal stock.
To deter dolphins from depredation, a number of mitigation techniques such as acoustic
deterrents, gear modifications, and avoidance are possible, but each technique has advantages
and disadvantages that must be considered. Acoustic devices have been shown to be effective in
deterring some other marine mammal species, but are likely to draw bottlenose dolphins towards
fishing activities, acting as a “dinner bell”. Gear modifications and avoidance may create
additional economic burdens for anglers, but are more promising and deserve further study.
The analysis presented here can help inform management and mitigation of bottlenose
dolphin depredation in the Gulf of Mexico and provides a baseline for future scientific study.
This study was the first to analyze the factors influencing the incidence of bottlenose dolphin
depredation using data from fishery observers. The use of previously existing data reduced the
time and funding necessary for this study, but also created limitations. The dataset was created
for fishery managers and needed to be intensively edited for use with this depredation case study.
Furthermore, the relatively small number of depredation events compared with the total number
of observations, and the manner in which depredation events were recorded complicated this
assessment. Future research should employ experimental designs that are designed to address
depredation events.
ii
Table of Contents
Introduction………………………………………………………………………………………..1
Materials and Methods……………………………………………………………………………5
Data Source……………………………………………………………………………… 5
Data Preparation………………………………………………………………………….8
Modeling………………………………………………………………………………....10
Model Selection and Analysis …………………………………………………………....10
Spatial Analysis…………………………………………………………………………..10
Results…………………………………………………………………………………………....11
Discussion………………………………………………………………………………………..15
Acknowledgements……………………………………………………………………………....24
References………………………………………………………………………………………..25
iiiIntroduction
Over the past few decades, depredation by common bottlenose dolphins (Tursiops
truncatus) has emerged as a complex conflict for marine resource management. In rod and reel
angling (private and for-hire) fisheries, depredation events can consist of a dolphin taking a
target fish off a hook as the angler is attempting to reel it in, but can also include taking bait off
the line or scavenging to collect fish that are thrown back after being caught (Tixier et al. 2020,
Zollett and Read 2006). Depredation has negative consequences for bottlenose dolphins because
dolphins that become conditioned to depredate have a higher chance of injury or mortality
(Christiansen et al. 2016). It can also lead to bycatch because depredating dolphins are more
likely to become entangled or ingest fishing gear (Read 2008, Wallace 1985). Bycatch is non-
target catch that becomes trapped in gear during fishing (Read 2008). Bycaught animals may be
difficult to free from gear, causing serious injury or death to the individual (Read 2008, Wells
and Scott 1994). Dolphins can also be injured or killed due to vessel strikes when depredating
(Wells and Scott 1997). Depredation causes additional economic burdens for private and for-hire
anglers who lose out on profitable catch or need to purchase extra bait or replace gear to account
for losses to marine mammals (Read 2008, Tixier et al. 2020). Frustration stemming from
interactions with dolphins can lead to harassment and retaliation efforts (Department of Justice
2006, Department of Justice 2007, NMFS 2016, Read 2005, Zollet and Read 2006). The Gulf of
Mexico has been referred to as a hotbed for retaliation against depredating bottlenose dolphins
and several documented observations of retaliation have been observed (Department of Justice
2006, Department of Justice 2007, NMFS 2016, Vail 2016, Wursig 2017).
In the Gulf of Mexico, there have been numerous anecdotal reports of bottlenose dolphin
depredation events in rod and reel (also known as hook and line) fisheries. In Florida, there have
been several documented cases of dolphin depredation in various locations such as Destin,
1Panama City, and Sarasota Bay, and some private and for-hire anglers report increased
depredation over time (Cunningham-Smith et al. 2006, Powell et al. 2018, Powell and Wells
2011, Samuels and Bejder 2004, Shippee et al. 2017, Zollet and Read 2006). There are several
potential reasons for depredation, which likely interact with one another in the Gulf. One reason
is overfishing, which has depleted fish stocks sufficiently to create direct competition between
fisheries and dolphin populations (DeMaster et al. 2001, Read 2008, Reichmont et al. 2018). In
the Gulf of Mexico, stable reef fish populations lead to healthy ecosystem functions such as
balanced predator-prey interactions (Coleman and Koenig 2010). However, the effects of climate
change and other anthropogenic disturbances further exacerbate this balance (Coleman and
Koenig 2010). In the northeastern Gulf, competition is also driven by declines in fish populations
after periodic harmful algal blooms, increased fish predation by the invasive lionfish, and the
2010 BP Deepwater Horizon Oil Spill (Green et al. 2012, Lewis et al. 2020, Powell and Wells
2011). Another potential reason for depredation by dolphins in Florida is intentional feeding.
Higher incidence of food provisioning escalates depredation because conditioned dolphins learn
that humans offer the potential opportunity for a meal (Powell et al. 2018, Samuels and Bejder
2004). This conditioned behavior may be passed on to social associates and younger individuals
through cultural transmission (Christiansen et al. 2016, Herzing 2005, Wells 2003).
While bottlenose dolphins are not threatened or endangered under the Endangered
Species Act, they are protected under the Marine Mammal Protection Act (MMPA) of 1972 and
require conservation consideration in the United States. Bycatch, feeding, and intentional harm
are deemed “takes” under the MMPA and are thus prohibited except under specific permits
issued by protected species managers (MMPA 1972). Permits for takes are based on level of
potential biological removal (PBR), calculated based on estimated population size, reproduction
2rate, and a recovery factor (MMPA 1972). Take associated with bycatch is hard to avoid due to
its unintentional nature, but other activities like feeding and retaliation are intentional and
avoidable behaviors that constitute harassment and are always considered illegal (MMPA 1972).
By reducing illegal feeding, depredation interactions will also likely be reduced and angler
frustration with dolphins and associated occurrence of retaliation may also be slowed or halted.
To more thoroughly understand the effects of depredation on bottlenose dolphin
populations in the Gulf, it is important to study the systems that create opportunities for
depredation to take place (Read 2008, Tixier et al. 2020). While there have been prior studies of
bottlenose dolphin depredation on rod and reel fisheries in Florida and Gulf of Mexico waters,
they often did not incorporate fisheries data, while many others focused on crab pot, gillnet, and
troll fisheries (Noke and Odell 2002, Powell and Wells 2011, Reichmont et al. 2018, Shippee et
al. 2017, Zollett and Read 2006). Despite these examples, there is little understanding of the
impact of dolphin depredation on rod and reel private and for-hire fisheries.
Rod and reel fisheries can be more complicated for data collection because there are often
several different relevant fishing modes that fall under this category (National Research Council
Ocean Studies Board 2006, Powers and Anson 2016). For instance, individuals can participate in
rod and reel fishing off of a private vessel, or a group of people can take part in a fishing charter
that is owned and operated by another party. Individuals can also fish off of a headboat that is
operated similarly to a fishing charter. Despite differences in fishing mode, all of these
approaches use the same types of rod and reel gear and likely target the same species, making
them all potential opportunities for depredation. However, surveys interested in understanding
depredation may only target boat captains or may only be surveying anglers at limited access
3points, missing important and relevant information from anglers (National Research Council
Ocean Studies Board 2006, Powers and Anson 2016).
While depredation has been studied in other fisheries, none have closely examined trends
in depredation for rod and reel private and for-hire anglers. Recreational fishing, and rod and reel
angling in particular, is important in the Gulf of Mexico, accounting for 64% of total landings
and exerting particular influence on high-value species like red snapper and red drum (Coleman
et al. 2004, Powers and Anson 2016). Using a dataset collected by the fisheries-dependent
monitoring at-sea observer program operated and maintained by the Florida Fish and Wildlife
Conservation Commission (FWC), the aim of this study is to determine whether bottlenose
dolphin depredation has increased over the last decade, where these interactions are most often
happening, which dolphin stocks are depredating most, and understand factors related to
bottlenose dolphin depredation events with rod and reel anglers. To do this, I modeled the
relationships between depredation events and several independent variables that fell into three
categories. The first two categories of independent variables I was interested in were geographic
location and behavioral characteristics of anglers and vessels. The third category examined the
relationships between depredation events and the fish being depredated. Year was also included
to determine whether depredation has been increasing over time. The dataset includes
information about whether depredation occurred in each observation, which provides the
capability to determine which relevant factors influence depredation. I also conducted a spatial
analysis to examine the geographic data visually and attempt to determine which dolphin stocks
were depredating. Creating context on the specifics of depredation, such as whether it is
increasing, where it is happening, what fish are most often targeted, which dolphin stocks are
4depredating, and what techniques anglers are using while on-board is important for future
research on potential mitigation strategies for depredation.
Materials and Methods
Data Source
The dataset used was collected as part of the FWC fisheries-dependent monitoring at-sea
observer program, which consisted of observations recorded by FWC fisheries observers on rod
and reel fishing trips between 2009 and 2020. Observations run along the Florida gulf coast,
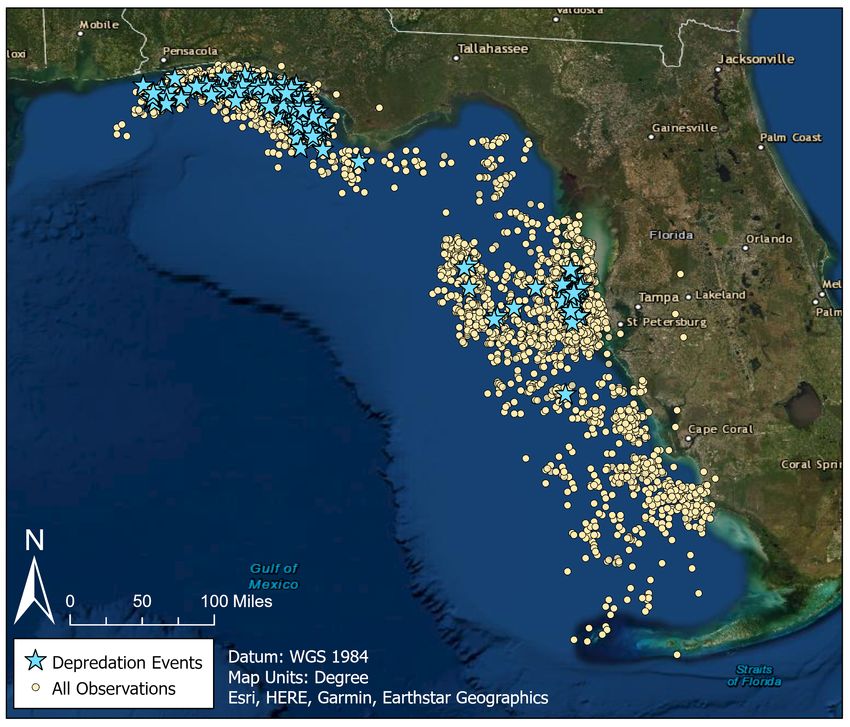
although some are located in Alabama waters (Figure 1). Each observation represented one fish
caught by an angler. For each observation, there were >100 associated variables recorded by the
observer to provide context for the conditions that gave rise to the fish being caught and released
or kept by the angler. Observations were identified by a reference number that was created using
the date of the trip, a trip number for that date, and a station number that the vessel was fishing
at. Stations were unique to the trip and were organized in chronological order from the beginning
of the trip (the first station) to the end of the trip (the last station). At each station, another
variable identified the chronological order in which fish were caught in. Although the dataset is
extremely comprehensive, there are instances where an observer did not record all fish caught
during the trip, particularly on larger vessels where there may have been visual impairments or
too many anglers for the observer to keep a constant record.
Events that were considered depredation were recorded as a factor under a variable that
included several values describing potential actions that could happen to the fish following
release. This variable was only recorded upon discard, so it was only possible to analyze
depredation events that happened following release, although it is important to note that
depredation is most often reported by anglers as removal of captured fish still on the line prior to
5bringing on-board the vessel. Additionally, this variable did not specify the species of marine
mammal involved in the interaction. However, spotted dolphins and bottlenose dolphins are the
only two marine mammal species that can be found on the continental shelf in the Gulf of
Mexico, and of those two species, bottlenose is the only one associated with depredation,
scavenging, and begging behavior (Balmer et al. 2016, Samuels and Bejder 2004, Powell and
Wells 2011, Powell et al. 2018, Tixier 2020). To use this dataset, I assumed that all depredating
marine mammals included in the dataset were bottlenose dolphins because all depredation events
occurred on the continental shelf (Figure 1).
Figure 1. Map of the gulf coast of Florida indicating all observations in the FWC fisheries dependent
monitoring at-sea observer program dataset, with depredation events highlighted as stars. The continental
shelf is indicated in lighter blue.
6Independent variables used in the final models were chosen based on initial examinations
of the dataset and their potential to influence dolphin behavior. Many of the variables deemed
potentially important for this study were factors rather than continuous variables. I chose to use
water depth and location based on Florida Fish and Wildlife’s fishing zone area codes to
examine the relationship between geographic location and depredation events. The zone codes
were designated based on latitude and longitude and provided one variable that integrated these
two factors together (Figure 2). The second group of variables included the vessel type, the
fishing mode, and the number of anglers fishing. The vessel types included charter, headboat,
multi-day trips, and research trips. Multi-day and research trips took place on-board charter and
headboat vessels, but were categorized separately and made up a very small portion of the
dataset. Fishing modes included anchored fishing, drift fishing, trolling, and holding, also known
as idling. The variables included regarding the characteristics of depredated fish included the
taxonomic family of fish, fish fork length (length of the fish from front of face to middle of the
tail) and whether the fish was vented before release. The use of bait was originally included as a
variable, but was later eliminated from consideration because it had a very low amount of
complete observations, which both reduced the model’s ability to accurately predict its
occurrence and severely truncated the dataset. Another variable that was ultimately discarded
was whether the fish was alive or dead on release, but all depredation events were on fish that
were alive. Year was included into the model to determine whether depredation was increasing
over time.
7Figure 2. Florida Fish and Wildlife Conservation Commission fishing area code map. Provided by FWC.
Data Preparation
All data manipulation and analysis was completed using R 3.6.1 software (R Core Team,
2019). To isolate the ‘preyed upon by a marine mammal’ value of the actions post-release
variable, a new binary depredation variable was created, where 1 referred to all instances where
the ‘preyed upon by marine mammal’ value was found, and all other observations were 0 (Figure
1).
Fish species were truncated to a taxonomic family level to reduce the number of levels
for the variable. Initially, there were 230 individual fish species originally recorded in the
dataset, including reef fishes, demersal fishes, large pelagics, elasmobranchs, and cephalopods
(Table 1). Once these observations were reduced to the family level, there were still 65 families,
many with very few observations. Thus, due to the size of the dataset, only families with >1,000
observations in the complete dataset were included as separate factors, and all other families
were included under one ‘other’ factor (Table 1). Five families were separated with >1,000
observations and 55 families were combined into the ‘other’ category (Table 1). Although this
8removed some families from individual consideration in the model, I felt that these divisions
were more representative of the most commonly captured fish and could accurately display the
variability I wanted to address.
Fish Family Species Included
Gray triggerfish, ocean triggerfish, queen triggerfish
Balistidae
African Pompano, almaco jack, amberjacks, Atlantic bumper, Atlantic moonfish, banded
Carangidae
rudderfish, bar jack, bigeye scad, blue runner, crevalle jack, greater amberjack, horse-eye jack,
jacks and pompanos*, leatherjacket, lesser amberjack, mackerel scad, permit, rainbow runner,
round scad, rough scad, yellow jack
Haemulidae
Bluestriped grunt, cottonwick, French grunt, margate, tomtate, white grunt, pigfish, porkfish
Lutjanidae Cubera snapper, dog snapper, gray snapper, lane snapper, mutton snapper, red snapper,
schoolmaster, yellowtail snapper, vermillion snapper
Bank sea bass, black grouper, black sea bass, butter hamlet, coney, creole-fish, gag, goliath
Serranidae grouper, graysby, red grouper, red hind, rock hind, rock sea bass, sand perch, scamp,
soapfishes*, Spanish flag, yellowmouth grouper, tattler, whitespotted soapfish, yellowedge
grouper
Atlantic bonito, Atlantic croaker, Atlantic mackerel, Atlantic sharpnose shark, Atlantic
spadefish, Atlantic thread herring, banded jawfish, bandtail searobin, bandtail puffer, bank
butterflyfish, bank cusk-eel, baitfish, barracudas*, batfishes*, Bermuda chub, bigeyes*, bighead
searobin, blackbar drum, blackbelly rosefish, blackedge moray, blackfin tuna, blackline tilefish,
blacknose shark, blacktip shark, blackwing searobin, blowfishes*, bluefishes*, blueline tilefish,
blue parrotfish, bonnethead, bull shark, cero, chub mackerel, clearnose skate, cobia, checkered
puffer, common moray, common puffer, conger eel, cubbyu, doctorfish, dolphinfish, dusky
damselfish, dusky flounder, dusky shark, filefishes*, finetooth shark, Florida smoothhound,
flounders*, gafftopsail catfish, great barracuda, grass porgy, gray angelfish, green moray,
guaguanche, gulf flounder, gulf toadfish, hammerhead shark, hardhead catfish, high-hat,
hogfishes*, inshore lizardfish, jawfishes*, jolthead porgy, king mackerel, knobbed porgy,
lefteye flounder, ladyfish, lemon shark, leopard toadfish, lionfish, little tunny, littlehead porgy,
Other lizardfishes*, loggerhead sea turtle, longspine squirrelfish, mackerels*, morays*, Northern
sennet, Northern puffer, nurse shark, ocellated frogfish, oceanic puffer, ocellated flounder,
ocellated moray, octopus*, orange filefish, oyster toadfish, pearly razorfish, pinfishes*,
planehead filefish, porgies*, puddingwife, red drum, red mullet, red porgy, redband parrotfish,
remora, requiem shark, reticulate moray, sailfish, sandbar shark, sanddabs, sand diver, sand
seatrout, sand tilefish, saucereye porgy, scaled sardine, scorpionfishes*, scrawled cowfish,
scrawled filefish, sea robins*, sharks*, sharksuckers*, sharpnose lizardfish, sheepshead, short
bigeye, silky shark, slippery dick, silver seatrout, smooth dogfish, smooth puffer, snakefish,
Southern flounder, Southern sennet, Southern stingray, Southern puffer, Spanish hogfish,
Spanish mackerel, Spanish sardine, spiny dogfish, spiny lobster, spinycheek scorpionfish,
spinner shark, spot, spotfin hogfish, spottail pinfish, spotted moray, spotted scorpionfish,
spotted seatrout, squirrelfishes*, stingrays*, striped burrfish, tiger shark, tilefishes*, toadfishes*,
tripletail, turtles*, unicorn filefish, wahoo, white marlin, whitebone porgy, whitefin sharksucker,
yellowcheek wrasse
Table 1. Fish species included in truncated taxonomic family groups. Species denoted with an
asterisk were only identified at the genus level by the at-sea observer.
9Many of the depredation events happened on the same trip or at the same station, so that
those observations were not independent. To account for this issue, a binary lag variable was
created where 0 was used for all observations before the depredation event on a trip and 1
included the depredation event and all subsequent observations on the trip. The lag variable was
incorporated as an independent variable in all models.
Modeling
I developed a generalized linear model (GLM) to examine the potential influence of
independent variables listed above on the occurrence of depredation. I used binomial regression
to portray the proportional distribution of depredation events to all other observations. The
dataset consisted of 333,912 observations with 110,575 complete observations ultimately
included in modeling. This subset of the data included 269 depredation events. The original
model included all independent variables but was parsed to find the model with the best fit.
Model Selection and Analysis
Model summaries, Akaike’s Information Criterion (AIC) and a chi-squared analysis of
variance (ANOVA) were all methods used in tandem to determine the model with the best fit to
the data. I used the Variance Inflation Factor (VIF) to test for collinearity and the ratio of
deviance and residual degrees of freedom to assess dispersion. I verified the goodness-of-fit of
the selected model using r.squaredGLMM, which calculates the amount of variation that can be
described by the variables. Coefficients were transformed using inv.logit to determine the
probabilities of the effects of each predictor variable on depredation events.
Spatial Analysis
A spatial analysis was conducted using ESRI’s ArcGIS Pro software (ESRI Inc. 2021).
Points were generated to designate all observations using the latitude and longitude coordinates
recorded in the fishery observer dataset. Spatial data for Gulf of Mexico bottlenose dolphin
10stocks was provided by NMFS, and was used to determine which stocks were depredating based
on where depredation happened in the fishery observer dataset. An examination of the
frequencies of depredation events was also conducted in each fishing zone to understand where
depredation hot spots occurred.
Results
After examining the summaries of parsed models with various combinations of the
predictors, I determined that the most parsimonious model had the best fit (ANOVA: pof all depredated fish (Table 2, Table 3). In comparison, the Carangidae family, made up of
jacks, pompanos, and scad, made up only 0.01% of all observations and 4% of depredated fish,
but was also a significant factor in the model (Table 2, Table 3).
Table 2. Results of the GLM.
Serranidae, or grouper and sea bass species, made up 0.03% of all observations but 14% of
depredated fish (Table 3). The ‘other’ category only had 0.004% depredation out of all
observations, but made up 1.49% of all observations, so although the frequency of depredation
was low, the variable was still significant (Table 2, Table 3). While these families were
depredated, the frequency of depredation on Lutjanidae species is much higher (Table 3).
12Table 3. Table indicating frequency of depredation events on each fish family within the context of the
entire dataset of observations.
More recent years since 2015 were significant in the model, with the exception of 2016
(Table 2, Figure 3). 2012 had a high number of depredation events and the largest frequency of
depredation events out of all observations, but it was not significant (Figure 3, Table 2). This
may also be attributed to the large number of observations that year, which was more than any
other year in the dataset.
Figure 3. Frequency of depredation occurrences out of all observations per year.
13The spatial analysis informed where depredation events are occurring and which dolphins
are depredating. Using spatial data relaying the boundaries of dolphin stocks along the gulf coast,
I was able to determine that bottlenose dolphins from the northern coastal stock are those most
often depredating in the waters off Panama City and Destin, Florida (Figure 4). Bottlenose
dolphins from the eastern coastal stock are depredating near Clearwater, Florida (Figure 4). The
coastal stocks are most often found between coastline beaches and the 20 meter isobath, whereas
bay stocks are found in coastal bays and estuaries (Balmer et al. 2019, Figure 4). The bay stocks
rarely leave those enclosed ranges and thus are less likely to be depredating out on the
continental shelf (Balmer et al. 2019, Figure 4). While the locations in the panhandle were both
significant in the model, the location off the coast of Clearwater was not, but the
Figure 4. Map of bottlenose dolphin stocks and depredation events in the northeastern Gulf of Mexico.
Bathymetry data developed by Scripps Institute of Oceanography at UCSD.
14frequency of observations that had depredation events in the Clearwater fishing zone was much
lower than the frequencies found in panhandle fishing zones (Table 2, Figure 5).
Figure 5. Percent of observations that include depredation in each geographic fishing zone designated by
FWC in the northeastern Gulf of Mexico.
Discussion
The variables that were significant in the model point to a combination of foraging theory
and prey preference as reasons for increased depredation. The probability of a depredation event
occurring was high in the Panama City and Destin areas, where there has already been a high
percentage of depredation events between 2009 and 2020 (Table 2, Figure 5). Thus, our data
15backs up the evidence observed in Panama City and Destin that conditioned dolphins with more
access to human interactions are more likely to depredate. Since the 1980s, there is ample
evidence of food provisioning in the northeastern gulf to high numbers of wild dolphins, who
become conditioned to initiate begging and scavenging when vessels and people are nearby
(NMFS 1994, Powell et al. 2018, Samuels and Bejder 2004). In these cases, conditioned
dolphins engaged with humans between 73-77% of the time when a human stimulus was within
50 meters (Powell et al. 2018).
Feeding dolphins to encourage them to participate in swim-with programs is another
widely practiced means for food provisioning in the Panama City and Destin areas, and also
increases opportunities for dolphins to become conditioned to foraging for food provided by
humans (Powell et al. 2018, Samuels and Bejder 2004, S. Horstman 2021, personal
communication). Although securing food provisions the dolphin with energy, prey search and
capture require both time and energy as inputs. To maximize fitness, dolphins adopt foraging
tactics that provide the most energy from prey at the lowest cost in terms of conserving time and
energy (Torres and Read 2009, Weiss 2006). Traits like high plasticity and knowledge transfer
between conspecifics make it easier for bottlenose dolphins to find and exploit foraging tactics
based around human behaviors, such as food provisioning and fishing, for their own benefit
(Torres and Read 2009, Weiss 2006). One study on bottlenose dolphins in Florida observed that
individuals that specialized in a foraging tactic would focus on using only that tactic as much as
possible, and even limit their spatial distribution to facilitate the use of their specialized foraging
technique (Torres and Read, 2009). Powell et al. (2018) witnessed two conditioned dolphins who
were known beggars directly attempting to depredate from recreational anglers in Panama City,
16indicating that these foraging tactics all fall under the same specialization skill set for bottlenose
dolphins.
There are three coastal stocks of bottlenose dolphins that stay within the bounds of the
continental shelf break in the Gulf of Mexico, as well as 31 smaller stocks that primarily reside
in bay, sound, and estuary (BSE) areas with more restricted ranges than the coastal stocks
(Balmer et al. 2019). Seven BSE stocks and one coastal stock can be found in the Florida
panhandle alone (Figure 4, Balmer et al. 2016, NMFS 2016). There is minimal spatial overlap
between BSE and coastal stocks in the Panama City and Destin areas, the groups are rarely
sighted together, and stocks have significant genetic differentiation (Balmer et al. 2016, Balmer
et al. 2019, Sellas et al. 2005). The coastal stock can often be found swimming along shore, and
is more often the stock targeted for food provisioning and swim-with programs by beach goers
and tourism ventures, although individuals from both stocks have been documented being
provisioned (Figure 4, Balmer et al. 2016). Animals in the same stock are more likely to exhibit
social transmission, so the dolphins that learn to forage by means of human interaction near the
beach are able to spread that knowledge to others and ultimately increase depredation at
nearshore reefs on the continental shelf (Herzing 2005, Wells 2003). In 2016, the northern
coastal stock was estimated to be around ~ 7000 individuals, while the St. Andrews Bay stock
found in the bays around Panama City is estimated at ~200 individuals (Balmer et al. 2019).
Both stocks are experiencing mortality and serious injury at rates higher than their PBR (NMFS
2016, NOAA 2019). Behavior resulting from provisioned dolphins by both dolphins and anglers
include depredation and retaliation. Thus, one or more of the mitigation practices outlined below
need to be implemented in the Panama City and Destin areas immediately in order to eliminate
17the occurrence of these behaviors, reducing incidence of depredation events for anglers and
serious injury and mortality for dolphins.
I determined that for each unit increase in the number of anglers, the probability of a
depredation event occurring also increased (Table 2). More anglers fishing in one area are more
likely to be noticed and depredated by conditioned dolphins foraging for food, particularly since
bottlenose dolphins have been documented begging and scavenging around fishing boats once
they realize that there are opportunities for food (Samuels and Bejder 2004, Shippee et al. 2017,
Powell et al. 2018). Conditioned dolphins have also been documented repeatedly returning to
areas where anglers normally fish to forage, or restricting their range to only include those areas
(Cantor et al. 2018). Thus, in Panama City and Destin where dolphins have already become
closely associated with humans, it is likely that they have learned to follow boats to known
fishing sites and end up staying in the area. Another study also reported bottlenose dolphins
responding to fish vocalizations by turning towards the sound and increasing echolocation
frequency, which would also be more likely around boats with many anglers at a large fishing
ground (Gannon et al. 2005). Ultimately, it is understandable that this variable is significant
while others examining angler behavior are not. Vessel type and fishing mode are more human-
centric variables, whereas number of anglers more directly considers dolphin sensation and
behavior.
The Lutjanidae family, which includes snapper and schoolmaster species, were
depredated in 78% of all depredation events, indicating a high overall level of depredation in
comparison to all other families (Table 3). This could be attributed to dolphin foraging theory,
but may also indicate a prey preference for bottlenose dolphins in the Gulf. There is evidence
that marine mammals target prey based on quality rather than focusing solely on quantity
18because the metabolic cost of living is correlated with diet quality (Spitz et al. 2012).
Additionally, bottlenose dolphin prey preference for soniferous fishes (such as species found in
the Lutjanid, Serranid and Carangid families) has been demonstrated in Florida (McCabe et al.
2010). Consistency in the diet of bottlenose dolphins residing in a restricted range has been
demonstrated in Florida, indicating that populations do exhibit prey preference, and prey species
recorded in the study examining bottlenose dolphin stomach contents included snapper species
(Dunshea et al. 2013). Additionally, anglers in the field have observed bottlenose dolphins
exhibiting prey preference for snapper species (J. Powell 2021, personal communication). Thus,
there is a high likelihood that bottlenose dolphins off the Gulf coast of Florida do prefer those
species and target them specifically as prey.
The frequency of snapper depredated is important to consider for Gulf of Mexico anglers,
who target red snapper under strictly managed regulations such as shortened catch seasons and
required discards of fish smaller than the legal size limit. After discarding fish below the legal
limit and watching them be depredated by dolphins, anglers may become more frustrated with
the imposed regulations and thus report depredation more frequently. This has also been
suggested by Shippee et al. (2017) as a reason for increased complaints. This increase in
complaints is justified given our results, which show that over time, depredation is increasing.
While the frequency of depredation events stayed fairly even over time except for a large number
in 2012, 2015, 2017, 2018 and 2019 were all significant to the model, indicating that there was a
higher probability of depredation happening in those years (Figure 3, Table 2). There was not
enough data from 2020 recorded at the launch of this study to determine a continuation of the
trend. Depredation is part of a harmful cycle of human interactions with dolphins, and since
anglers are struggling to deal with increasing events and their effects on economically important
19catch, it is imperative that changes are implemented sooner rather than later to reduce this
particular trend.
Several studies have focused on venting as a means for mitigating barotrauma in reef fish
that may be impaired after being caught (Drumhiller et al. 2014, Pulver 2017). While venting has
demonstrated improvements in recovery for fish and has been thought to assist them in evading
predatory marine mammals, it was not significant in this study and was not included in the final
model (Drumhiller et al. 2014). However, using barotrauma mitigation tactics as a preventative
measure against depredation may be useful for anglers. There is evidence that anglers prefer
other barotrauma mitigation methods, such as descending devices, and that these may be better at
preventing depredation from dolphins as well as other large predators such as sharks because the
behavior of the fish on a descending device is more fluid and natural, rather than erratic (Ayala
2020, Curtis et al. 2019, Shippee et al. 2017). However, there is also anecdotal evidence that
bottlenose dolphins can learn to depredate fish off descending devices in short periods of time (J.
Powell 2021, personal communication). This is not surprising given the high plasticity and
preference for low cost foraging behavior that bottlenose dolphins are known for, particularly in
uncommon habitats and across varying prey sources (Torres and Read 2009, Weiss 2006).
Other potential mitigation techniques for reducing the occurrence of dolphin depredation
events include the use of deterrents such as acoustic pingers, gear modifications, and avoidance.
Deterrent devices and gear modifications are often used as mitigation techniques for marine
mammal bycatch, but have also been studied in the context of reducing depredation (Dawson et
al. 2013, Hamer et al. 2015, McPherson 2011, Tixier et al. 2020, Zollett and Read 2006). While
both methods have demonstrated some success at deterring marine mammals from depredating, a
comparison of the two in a review of depredation mitigation techniques showed that gear
20modifications have emerged as the more promising method, particularly for foraging bottlenose
dolphins, where conclusions on pinger usage have been mixed (Dawson et al. 2013, Hamilton
and Baker 2019, Tixier et al. 2020). While some groups have seen small reductions in
depredation when pingers are used, a complete elimination of depredation using this method has
never been described, and there are no examples of long term pinger use in any fishery (Dawson
et al. 2013). In fact, the use of pingers for bottlenose dolphins has been linked to the “dinner bell
effect” whereby bottlenose dolphins become habituated to the sound and use it to determine
where to depredate, despite gear type, due to their characteristic food motivation (Cox et al.
2004, Dawson et al. 2013). However, a potential gear modification for rod and reel use was
created and tested on the Florida king mackerel commercial troll fishery in 2006, and was
successful at deterring bottlenose dolphins from depredating without reducing catch (Zollett and
Read 2006). Further studies using this methodology may be helpful for private and for-hire
anglers to determine whether it should be expanded as an option for reducing depredation. In
practice, both acoustic pingers and gear modifications create supplementary economic burdens
for anglers, because they become responsible for purchasing pingers or new gear and may need
to hire more crew members to help with new methods for catch (Dawson et al. 2013, Hamer et
al. 2015). The method most widely accepted as best for depredation mitigation is avoidance,
because it is the only method that relies on human behavior to implement (Fader et al. 2021,
Werner et al. 2015). There are two tactics for avoidance: moving on from sites where dolphins
are and targeting fishing times and locations where dolphins can be avoided altogether (Tixier et
al. 2020). While avoidance is seen as a safer and more effective way to prevent depredation, it
may increase costs for anglers because they must travel more to move on from a fishing site
where dolphins are depredating or to get to sites where they know that dolphins may not be
21(Tixier et al. 2020). It also may require more planning and consideration for dolphin behavior
than other methods, which may be burdensome for some anglers (Werner et al. 2015). However,
implementing avoidance in the Gulf for a set period of time may cause dolphins to develop other
foraging techniques to account for losses from depredation, giving anglers more flexibility in the
long term for fishing where they would like.
As bottlenose dolphin depredation continues to be problematic for anglers, this study
provides more understanding of the trends that may be contributing to the issue, and can be used
to inform protected species management concerned with preserving local bottlenose dolphin
populations and anglers hoping to reduce interactions with dolphins. The variables deemed
significant by the model definitively point to the problems with food provisioning in Panama
City and Destin as management issues that need to be addressed. A reduction in illegal dolphin
feeding would likely prevent increases in dolphin scavenging, begging, and depredation, or even
reduce depredation over time. While reducing the number of anglers and targeting non-preferred
prey species will likely also reduce depredation, these would be unrealistic recommendations.
However, these trends do warrant consideration for fisheries and protected species management
as fundamental dynamics of the interactions between dolphins and anglers in the Gulf and
knowledge of their roles will be important for future studies of dolphin depredation. These trends
point to learned dolphin foraging tactics and prey preference as reasons for increasing
depredation in the Gulf, and mitigation techniques need to be implemented in order to reverse the
effects of these behaviors. Ultimately, other studies are needed to address the gaps in knowledge
for rod and reel depredation. Several reviews of depredation studies and mitigation techniques
echo this opinion, as data for closely examining the effects of depredation and mitigation in
different fisheries systems are often lacking or incomplete (Tixier et al. 2020, Werner et al.
222015). A more direct approach to studying these interactions would be useful to complement the
results found using the fishery observer dataset and could incorporate an examination of
potential mitigation techniques like barotrauma mitigation tools, deterrent devices, and
avoidance tactics. This study examined data from for-hire fishing vessels, which includes several
relevant stakeholder groups, but excludes private anglers. Future studies able to incorporate data
from private anglers would be better for diagnosing issues with depredation in the Gulf by
incorporating data from all affected rod and reel groups, providing more information about who
is depredated, although according to our research, it is likely that for-hire vessels experience
more depredation because there are more anglers with lines in the water. Examining the use of
bait as a trend for depredation would also be applicable. I attempted to look at bait, but there was
a large amount of missing data which rendered the variable not useful for the model. Finally,
expanding study of bottlenose dolphin depredation events on rod and reel angling ventures
across the entire Gulf of Mexico would provide managers with more information and allow them
to apply mitigation techniques more holistically across the region. It would also provide
justification for other anglers who are concerned about depredation in their areas. Ultimately,
human interactions with bottlenose dolphins are hard to avoid in the Gulf. However, increasing
food provisioning and swim-with programs has led to an increase in depredation events that is
proving detrimental to local anglers. This cycle must be broken in order to reverse these trends
and give dolphins the space they need to learn to forage without the assistance of humans.
23Acknowledgements
I would like to thank the Florida Fish and Wildlife Conservation Commission, and particularly
Oscar Ayala, for sharing the data used to generate this report, Will Cioffi for consulting on
coding for edits to the dataset and statistical analyses and Matt Duggan for his help sorting fish
species.
24References
Adimey, N.M., Hudak, C.A., Powell, J.R., Bassos-Hull, K., Foley, A., Farmer, N.A., White, L. and K.
Minch. (2014) Fishery gear interactions from stranded bottlenose dolphins, Florida manatees and
sea turtles in Florida, USA. Marine Pollution Bulletin 81: 103-115.
http://dx.doi.org/10.1016/j.marpolbul.2014.02.008
Ayala, O. (2020) Testing the efficacy of recompression tools to reduce discard mortality of reef fishes in
the Gulf of Mexico. University of South Florida Graduate Theses and Dissertations. 38 pp.
Balmer, B., Morey, J., Quigley, B., Rowles, T., Speakman, T., Ylitalo, G., Zolman, E. and L. Schwacke.
(2016). Bottlenose dolphin occurrence in St. Andrews Bay, Florida, and coastal waters near the
naval surface warfare center, Panama City division testing range. MOA-2015-029/9087.
https://www.navymarinespeciesmonitoring.us/index.php/download_file/view/1803/
Balmer, B., Watwood, S., Quigley, B., Speakman, T., Barry, K., Mullin, K., Rosel, P., Sinclair, C.,
Zolman, E. and L. Schwacke. (2019) Common bottlenose dolphin (Tursiops truncatus)
abundance and distribution patterns in St Andrews Bay, Florida, USA. Aquatic Conservation
29(3): 486-498. https://doi.org/10.1002/aqc.3001
Cantor, M., Simoes-Lopes, P.C. and F.G. Daura-Jorge. (2018) Spatial consequences for dolphins
specialized in foraging with fishermen. Animal Behavior 139: 19-27.
https://doi.org/10.1016/j.anbehav.2018.03.002
Christiansen, F., McHugh, K.A., Bejder, L., Siegal, E.M., Lusseau, D., McCabe, E.B., Lovewell. G. and
R.S. Wells. (2016) Food provisioning increases the risk of injury in a long-lived marine top
predator. Royal Society of Open Science 3: 160560. http://dx.doi.org/10.1098/rsos.160560
Coleman, F.C., Figueira, W.F., Ueland, J.S. and L.B. Crowder. (2004) The impact of United States
recreational fisheries on marine fish populations. Science 305: 1958-1960.
10.1126/science.1100397
Coleman, F.C. and C.C. Koenig. (2010) The effects of fishing, climate change, and other anthropogenic
disturbances on red grouper and other reef fishes in the Gulf of Mexico. Integrative and
Comparative Biology 50(2): 201-212. doi:10.1093/icb/icq072
Cox, T.M., Read, A.J., Swanner, D., Urian, K. and D. Waples. (2004). Behavioral responses of bottlenose
dolphins, Tursiops truncatus, to gillnets and acoustic alarms. Biological Conservation 115(2):
203-212. https://doi.org/10.1016/S0006-3207(03)00108-3
Cunningham-Smith, P., D.E. Colbert, R.S. Wells, and T. Speakman. (2006) Evaluation of human
interactions with a provisioned wild bottlenose dolphin (Tursiops truncatus) near Sarasota Bay,
Florida, and efforts to curtail the interactions. Aquatic Mammals 32:346-356.
Curtis, J.M., Tompkins, A.K., Loftus, A.J., and G.W. (2019) Stunz. Recreational angler attitudes and
perceptions regarding the use of descending devices in southeast reef fish fisheries. Marine and
Coastal Fisheries: Dynamics, Management, and Ecosystem Science 11: 506-518. DOI:
10.1002/mcf2.10102
Dawson, S.M., Northridge, S., Waples, D. and A.J. Read. (2013) To ping or not to ping: The use of active
acoustic devices in mitigating interactions between small cetaceans and gillnet fisheries.
Endangered Species Research 19: 201-221. Doi: 10.3354/esr00464
DeMaster, D.P., Fowler, C.W., Perry, S.L., and M.F. Richlen. (2001) Predation and competition: The
impact of fisheries on marine-mammal populations over the next one hundred years. Journal of
Mammalogy 82(3): 641-651.
25Department of Justice. (2006, August 8) Florida Charter Boat Captain Pleads Guilty to Shooting at
Dolphins. [Press Release].
Department of Justice. (2007, January 19) Shooting of Dolphin Leads to Federal Charges. [Press
Release].
Drumhiller, K.L., Johnson, M.W., Diamond, S.L., Robillard, M.M.R. and G.W. Stunz. (2014) Venting or
rapid recompression increase survival and improve recovery of red snapper with barotrauma.
Marine and Coastal Fisheries 6(1): 190-199. DOI: 10.1080/19425120.2014.920746
Dunshea, G., Barros, N.B., Berens McCabe, E.J., Gales, N.J., Hindell, M.A., Jarman, S.N. and R.S.
Wells. (2013) Stranded dolphin stomach contents represent the free-ranging population’s diet.
Biology Letters 9: 20121036. http://dx.doi.org/10.1098/rsbl.2012.1036
Esri Inc. (2021) ArcGIS Pro (Version 2.7). https://www.esri.com/en-us/arcgis/products/arcgis-
pro/overview
Fader, J.E., Elliott, B.W. and A.J. Read. (2021) The challenges of managing depredation and bycatch of
toothed whales in pelagic longline fisheries: Two U.S. case studies. Frontiers in Marine Science
8: 618031. doi: 10.3389/fmars.2021.618031
Gannon, D.P., Barros, N.B., Nowacek, D.P., Read, A.J., Waples, D.M. and R.S. Wells. (2005) Prey
detection by bottlenose dolphins, Tursiops truncatus: an experimental test of the passive listening
hypothesis. Animal Behavior 69: 709-720. doi:10.1016/j.anbehav.2004.06.020
Green, S.J., Akins, J.L., Maljkovic, A. and I.M. Cote. (2012) Invasive lionfish drive Atlantic coral reef
fish declines. PLoS ONE 7(3): e32596. doi:10.1371/journal.pone.0032596
Hamer, D.J., Childerhouse, S.J., McKinlay, J.P., Double, M.C. and N.J. Gates. (2015) Two devices for
mitigating odontocete bycatch and depredation at the hook in tropical pelagic longline fisheries.
ICES Journal of Marine Science 72(5): 1691-1705. doi:10.1093/icesjms/fsv013
Hamilton, S. and G.B. Baker. (2019) Technical mitigation to reduce marine mammal bycatch and
entanglement in commercial fishing gear: Lessons learnt and future directions. Reviews in Fish
Biology and Fisheries https://doi.org/10.1007/s11160-019-09550-6
Herzing, D. (2005) Transmission mechanisms of social learning in dolphins: Underwater observations of
free-ranging dolphins in the Bahamas. Autour de l'ethologie et de la cognition animale 185-193.
Lewis, J.P., Tarnecki, J.H., Garner, S.B., Chagris, D.D. and W.F. Patterson. (2020) Changes in reef fish
community structure following the Deep Water Horizon oil spill. Nature Scientific Reports
10:5621. https://doi.org/10.1038/s41598-020-62574-y
Litz, J.A., Baran, M.A., Bowen-Stevens, S.R., Carmichael, R.H., Colegrove, K.M., Garrison, L.P., Fire,
S.E., Fougeres, E.M., Hardy, R., Holmes, S., Jones, W., Mase-Guthrie, B.E., Odell, D.K., Rosel,
P.E., Saliki, J.T., Shannon, D.K., Shippee, S.F., Smith, S.M., Stratton, E.M., Tumlin, M.C.,
Whitehead, H.R., Worthy, G.A.J. and T.K. Rowles. (2014) Review of historical unusual mortality
events (UMEs) in the Gulf of Mexico (1990-2009): providing context for the multi-year northern
Gulf of Mexico cetacean UME declared in 2010. Diseases of Aquatic Organisms 112: 161-175.
doi: 10.3354/dao02807
Marine Mammal Protection Act. (1972) 16 U.S.C. §§ 1361 et seq.
McCabe, E.J.B., Gannon, D.P., Barros, N.B. and R.S. Wells. (2010) Prey selection by resident common
bottlenose dolphins (tursiops truncatus) in Sarasota Bay, Florida. Marine Biology 157: 931-942.
DOI 10.1007/s00227-009-1371-2
26McPherson, G. (2011) Acoustic methods to mitigate bycatch and depredation by marine mammals on
commercial fishing operations in Australian waters: Fishermens options. Proceedings of
Acoustics Paper Number 101. 8 pp.
National Research Council Ocean Studies Board. (2006) Review of recreational fisheries survey methods.
The National Academies Press, Washington D.C. 187 pp.
NMFS. (1994) Report to congress on results of feeding wild dolphins: 1989-1994. 84 pp.
https://repository.library.noaa.gov/view/noaa/16196
NMFS. (2016) Common bottlenose dolphin (Tursiops truncatus truncatus): Gulf of Mexico northern
coastal stock. 11 pp. https://media.fisheries.noaa.gov/dam-
migration/f2015_bodoncoastgmex508.pdf
NOAA Fisheries. (2020) 2018-2020 Bottlenose dolphin unusual mortality event southwest Florida.
Accessed on February 22, 2021 at https://www.fisheries.noaa.gov/southeast/marine-life-
distress/2018-2020-bottlenose-dolphin-unusual-mortality-event-southwest
NOAA. (2020) US Atlantic and Gulf of Mexico Marine Mammal Stock Assessments - 2019. NOAA
Technical Memorandum NMFS-NE-264 497 pp. https://media.fisheries.noaa.gov/dam-
migration/2019_sars_atlantic_508.pdf
Noke, W.D. and D.K. Odell. (2002) Interactions between the Indian River Lagoon blue crab fishery and
the bottlenose dolphin, Tursiops truncatus. Marine Mammal Science 18(4): 819-832.
Powell, J.R. and R.S. Wells. (2011) Recreational fishing depredation and associated behaviors involving
common bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Publications,
Agencies and Staff of the U.S. Department of Commerce. 308.
https://digitalcommons.unl.edu/usdeptcommercepub/308
Powell, J.R., Machiernis, A.F., Engleby, L.K., Farmer, N.A. and T.R. Spradlin. (2018) Sixteen years
later: an updated evaluation of the impacts of chronic human interactions with bottlenose
dolphins (Tursiops truncatus truncatus) at Panama City, Florida, USA. Journal of Cetacean
Research and Management 19: 79-93.
Powers, S.P. and K. Anson. (2016) Estimating recreational effort in the Gulf of Mexico red snapper
fishery using boat ramp cameras: Reduction in federal season length does not proportionally
reduce catch. North American Journal of Fisheries Management 36(5): 1156-1166. DOI:
10.1080/02755947.2016.1198284
Pulver, J.R. (2017) Sink or swim? Factors affecting immediate discard mortality for the Gulf of Mexico
commercial reef fish fishery. Fisheries Research 188(2017): 166-172.
http://dx.doi.org/10.1016/j.fishres.2016.12.018
R Core Team. (2019) R: A language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
Read, A.J. (2005) Bycatch and depredation. In Marine Mammal Research: Conservation Beyond Crisis.
The Johns Hopkins University Press, Baltimore, Maryland. 5-17.
Read, A.J. (2008) The looming crisis: Interactions between marine mammals and fisheries. Journal of
Mammalogy 89(3): 541-548.
Rechimont, M.E., Lara-Dominguez, A.L., Morteo, E., Martinez-Serrano, I., and M. Equihua. (2018)
Depredation by coastal bottlenose dolphins (Tursiops truncatus) in the southwestern Gulf of
Mexico in relation to fishing techniques. Aquatic Mammals 44(5): 469-481. DOI
10.1578/AM.44.5.2018.469
27You can also read

















































