Neonatal and Children's Immune System and COVID-19: Biased Immune Tolerance versus Resistance Strategy - The ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Published August 21, 2020, doi:10.4049/jimmunol.2000710
Neonatal and Children’s Immune System and COVID-19:
Biased Immune Tolerance versus Resistance Strategy
Shokrollah Elahi
The recent outbreak of COVID-19 has emerged as a offensive because this evolutionary mechanism can prevent
major global health concern. Although susceptible to in- collateral damage and preserve resources in the host. The
fection, recent evidence indicates mostly asymptomatic concept of tolerance is not only restricted to infection and can
or mild presentation of the disease in infants, children, also be applied to other diseases, such as protective mecha-
and adolescents. Similar observations were made for nisms to reduce further tissue destruction in autoimmune
acute respiratory infections caused by other coronavi- diseases (8).
ruses (severe acute respiratory syndrome and Middle The natural host defense against infection aims to either
East respiratory syndrome). These observations suggest eliminate the pathogen or prevent/reduce damage to the host
that the immune system behaves differently in children (9). Thus, a host defense can be divided into two main cat-
egories: 1) the clearance or resistance mechanism and 2) the
than adults. Recent developments in the field demon-
tolerance mechanism. The resistance strategy protects the host
strated fundamental differences in the neonatal immune
by eliminating the pathogens and/or neutralizing their toxic
system as compared with adults, whereby infants re- and harmful products. This strategy is associated with the
spond to microorganisms through biased immune tol- induction of innate and adaptive immunity. Although this
erance rather than resistance strategies. Similarly, more strategy is essential to reduce the pathogen burden, it sustains
frequent/recent vaccinations in children and younger a significant cost to the host fitness (8). Collateral tissue
populations may result in trained immunity. Therefore, damage or immunopathology can be the negative conse-
the physiological abundance of certain immunosuppres- quence of host defense against infection (10). Of course, the
sive cells, a tightly regulated immune system, and/or ex- magnitude and duration of resistance mechanism can deter-
posure to attenuated vaccines may enhance trained mine the size of immunopathology. The optimal immune
immunity to limit excessive immune reaction to response requires a balance between the pathogen clearance
COVID-19 in the young. The Journal of Immunology, and the immunopathology (11). However, such balance is not
2020, 205: 000–000. always possible; thus, to reduce the fitness cost, an alternative
approach, tolerance mechanism, is crucial. Even though such
I
t has been reported that newborns and infants are highly a mechanism does not necessarily influence pathogen burden,
susceptible to infections (1). This has been attributed to it can reduce the collateral damage caused by the infection or
an underdeveloped or immature immune system (2, 3). the immune system against the pathogen (8). The best ex-
However, this notion has recently been challenged and ample is the lack of association between the pathogen burden
replaced by the presence of “active immunosuppression” in and the disease severity in bacterial infection (12, 13). Thus,
early life (4, 5). This concept contradicts the traditional no- tolerance reduces the susceptibility of the host to tissue
tion that the immune system aims to identify and neutralize damage without directly influencing the pathogen burden.
pathogens and instead suggests that the immune system en- Recent evidence indicates that infants are highly prone to
ables the host to tolerate the presence of nonpathogenic and host-mediated immunopathology in the absence of a toler-
pathogenic microorganism, which is a distinct and smart ance mechanism (14). For example, ,5–10% of neonatal
defense strategy (6). Although the offensive mechanism(s) to sepsis cases are culture proven and the rest are based on
eliminate the pathogen may work in some circumstances, it is symptoms and clinical signs, which can be related to an ex-
not always the best defense strategy as it can be enormously cessive immune response under certain circumstances (15). It
costly for the host (7). The natural selection in most cir- is thus not unpredictable that a tolerance mechanism has
cumstances favors coexistence and tolerance that are not evolved for allowing the establishment of diverse commensal
School of Dentistry, Faculty of Medicine and Dentistry, University of Alberta, Edmon- Address correspondence and reprint requests to Prof. Shokrollah Elahi, University of
ton, Alberta T6G2E1, Canada; Department of Oncology, Cross Cancer Institute, Uni- Alberta, 7020-Katz Group Centre, 11361-87th Avenue NW, Edmonton, AB T6G2E1,
versity of Alberta, Edmonton, Alberta T6G1Z2, Canada; Department of Medical Canada. E-mail address: elahi@ualberta.ca
Microbiology and Immunology, Faculty of Medicine and Dentistry, University of
Abbreviations used in this article: ACE2, angiotensin-converting enzyme 2; CEC,
Alberta, Edmonton, Alberta T6G2E1, Canada; and Li Ka Shing Institute of Virology,
CD71+ erythroid cell; HBV, hepatitis B virus; HCV, hepatitis C virus; hRSV, human
University of Alberta, Edmonton, Alberta T6G2E1, Canada
respiratory syncytial virus; MDSC, myeloid-derived suppressor cell; MERS, Middle East
ORCID: 0000-0002-7215-2009 (S.E.). respiratory syndrome; MMR, measles, mumps, and rubella; SARS, severe acute respira-
tory syndrome; Treg, regulatory T cell.
Received for publication June 15, 2020. Accepted for publication August 11, 2020.
This work was supported by the Institute of Infection and Immunity, Government of Copyright Ó 2020 by The American Association of Immunologists, Inc. 0022-1767/20/$37.50
Canada (360929 and 353953) and FASTGRANT.com.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.2000710
2 BRIEF REVIEWS: IMMUNE SYSTEM AND COVID-19 INFECTION IN THE YOUNG
microbial communities in newborns/infants (4). Also, a respiratory infections (32). Despite the extremely high fre-
tolerogenic approach is vital to prevent excess immune re- quency of human rhinovirus infection in healthy young
action to pathogens. Otherwise, a resistance mechanism children, most cases of infection remain asymptomatic or
strategy could cause increased morbidity and mortality in this present very mild symptoms (33). Human respiratory syn-
most vulnerable population (16). cytial virus (hRSV) is the main cause of respiratory infections
in infants. Although the majority of infected infants display a
Disease tolerance to viral infections in infants and children mild upper respiratory tract infection, this is now recognized
Decreased rather than increased immune response can be as- as a significant concern in the adult population (34, 35). How-
sociated with lesser immunopathology and protection against ever, there is still the unanswered question as to why some
certain pathogens in early life. The benefits of such at- unfortunate children suffer from severe bronchiolitis. Some
tenuated immune effectors are well described for viral in- risk factors such as prematurity, low birth weight, low
fections such as hepatitis B virus (HBV) and HIV in which the socioeconomic status, immunodeficiency, and pre-existing
immune regulatory mechanisms (e.g., active immunosup- conditions are reported as likely contributing factors influ-
pression) during early life attenuate excessive and poten- encing the development of a severe disease (36, 37). It has
tially harmful immune responses. For example, in infants and been reported that exacerbated immune response by infiltra-
children, HBV infection rarely results in acute hepatitis, tion of neutrophils, T cells, and proinflammatory cytokines
whereas this is common in adults (17). In addition, vertical can cause damage to respiratory tissue (38, 39). For example,
HBV infection often leads to chronic disease in infants, which hRSV upregulates NF-kB, resulting in the induction of IL-
might be associated with immunotolerance (17). This is also 18, TNF-a, CCl-5, and other inflammatory factors (40). In
the case for hepatitis C virus (HCV) as clearance of the virus line with this, the elevation of IL-6 and TNF-a has been
without treatment is more common in infants than adults associated with immunopathogenesis in hRSV-infected in-
(18). This transient HCV viremia and spontaneous viral fants with bronchiolitis (38). In particular, disease severity
clearance in children (19) might be related to the lack of a appears to be correlated with IL-6/TNF-a ratio in nasopha-
robust innate immune response, as shown by unchanged ryngeal fluids of infants with bronchiolitis (41). This implies
proinflammatory cytokines (20). These observations were that neutralization of TNF-a might have therapeutic impli-
further confirmed by a relative suppression of T cell activation cations as shown in a mouse model of respiratory syncytial
in HCV-exposed children. These observations suggest that a virus infection (42). Although the exact underlying mecha-
balanced pro- and anti-inflammatory response in HCV- nism of bronchiolitis in hRSV remains contentious, a pro-
exposed children might account for a spontaneous clearance nounced immune cell infiltration and proinflammatory
of the virus (21). The same has been found in CMV, EBV, cytokines support a role for the immune-mediated tissue
and varicella-zoster virus, which are other highly abundant damage (43). Therefore, the resulting balance of viral clear-
viral infections in children (22, 23). In most cases (75–80%) ance and immunopathology could be the key factor in de-
of CMV, infected newborns do not express clinical manifes- termining the disease outcome in respiratory syncytial virus
tations and excrete the virus for several years (24). This has infection. This differential disease outcome is also evident in
been attributed to the inability of the neonatal immune sys- some infected children with HIV as compared with adults. In
tem to control the viral replication in some organs (25). This pediatric HIV infection, disease progression in the absence of
lack of a robust immune response against the virus might be antiretroviral therapy is faster than in adults (44). However,
beneficial for the young because an inflammatory response this is not the case in all children as a subset of anti-
can result in immunopathology and collateral damage as retroviral therapy–naive children remains clinically healthy
reported in a neonatal mouse model (26). Similarly, in early with normal-for-age CD4 counts defined as pediatric non-
childhood, EBV infection is asymptomatic or associated with progressors (45). Nevertheless, the mechanisms associated
a middisease, whereas it often manifests as an acute infectious with nonprogression are markedly different in children versus
mononucleosis in adults because of an exaggerated immune adults. Nonprogressing adults, so-called elite controllers or
response (27, 28). The mechanism underlying the lack of long-term nonprogressors, are typically characterized by hav-
EBV symptoms in early life is not well defined; however, a ing polyfunctional CD8+ T cells restricted by certain HLA
lower number of new infectious virions is reported to be as- alleles (mainly HLA-B27 and HLA-B57) (46–49). Such a
sociated with mild and subclinical symptoms (29). Mecha- robust CD8+ T cell response results in the suppression of
nistically, limited viral Ags because of fewer infectious particles viremia beneath detection levels (normally ,50 HIV RNA
results in immune surveillance avoidance and is subsequently copies per milliliter of blood in elite controllers but ,10,000
associated with EBV persistence (29). As such, the occurrence in long-term nonprogressors) that high CD4+ T cell counts
of asymptomatic EBV in early childhood elicits limited virus- are maintained (46, 47). By contrast, nonprogressing HIV-
specific CD8+ T cells, whereas excessive expansion of CD8+ infected children are not enriched with protective HLA alleles
T cells is the cause of symptoms in adolescents (e.g., acute to reduce viral load (50). Therefore, in contrast to HIV-
infectious mononucleosis) (30). These observations suggest that infected adults, these children, despite having high viremia
the general concept of disease tolerance in infants can be (median 26,000 copies per milliliter), will maintain a normal
beneficial when it comes to CMV and EBV infections. The CD4+ T cell count (50) and low levels of immune activation
immunological basis for uncomplicated varicella-zoster virus in (51). This phenotype of normal CD4+ T cell counts with
children compared with life-threatening infection in healthy minimal immune activation levels despite high viral load
adolescents and adults is not well understood but might be suggests that the immune activation rather than virus repli-
related to changes in effector functions of APC and T cells cation may account for the disease progression (50). This
(31). Another example is human rhinoviruses responsible for concept is well accepted in the natural host of the SIV (e.g.,The Journal of Immunology 3
sooty mangabey and African green monkey) as low level of This examination becomes more interesting when 100%
immune activation is observed in these animals despite having mortality in adult patients has been associated with .100
high viral load (52). These observations suggest that the at- CFUs/ml blood, whereas mortality of 73% in neonates re-
tenuated immune response in this group of children enables quired .1000 CFUs/ml blood (64). Such has been validated
them to avoid the collateral damage and disease progression in in animal models that newborns can tolerate much greater
pathogenic infection and, in fact, protects them. Overall, bacterial loads than adults (65). These studies suggest that
mounting evidence supports the concept that neonatal im- newborns have a greater tolerance to bacteremia and can
munity is not underdeveloped but instead tightly regulated survive levels of bacterial load in their blood that normal
and dynamic in response to pathogenic and not pathogenic adults cannot. This delay in bacterial clearance has been re-
(e.g., microbiome) stimuli. Such an intelligent and regulated lated to inefficient phagocytosis and impaired neutrophil re-
immune system plays a crucial role in protecting growing cruitment in neonates (66, 67). For instance, it has been
infants for invaders while at the same time preventing exces- shown that neonatal monocytes/neutrophils exhibit impaired
sive immune response that can be costly for the host (53). phagocytic capabilities of Escherichia coli and Staphylococcus
Therefore, the de-escalating/tolerance versus resistance mecha- aureus in vitro, when whole blood was used (57, 68). These
nism against infections in infants is a protective and intelligent examples have contributed to the theory of neonatal imma-
mechanism for survival, continued growth, and development turity; however, this notion has been challenged by showing
when considering the infants’ limited resources. comparable phagocytosis capacity of neonatal and adult
monocytes and neutrophils in the absence of CECs in vitro
The benefits of higher infection tolerance in infants and children (57). In fact, we showed that the observed deficiency was
The newborn gets exposed to millions of bacteria in the birth related to the presence of CECs and active immunosuppres-
canal through natural delivery, which is the beginning of a sion in neonates and was not simply a state of deficiency in
long journey of host–microorganisms interactions. The fetus phagocytosis (1, 54, 57). These data highlight the difference
has minimal to no exposure to foreign Ags under normal in the used immune mechanism(s) in response to pathogens
circumstances, and therefore, in the absence of tolerance in neonates compared with adults. Given the higher mortality
mechanisms, such exposure could result in intensifying in- rate of sepsis in neonatal mice following exogenous supple-
flammation at the mucosal surfaces and skin where brisk mentation of proinflammatory cytokines (69), newborns
postnatal colonization with commensal microbes occurs (54). likely benefit from the immunosuppression that can inhibit
Therefore, a state of colonization tolerance is essential for the risk of a rapid and harmful immune response. Children
allowing the swift adaptation of commensal bacteria. This and adolescents may benefit from a similar phenomenon but
tolerance is mediated in part by the physiologically enriched by a differential mechanism. For instance, more frequent and
erythroid precursors, coexpressing CD71 (transferrin recep- more recent live attenuated viral vaccine immunizations can
tor) and CD235a in humans but CD71 and TER119 in mice promote the expansion of long-lived MDSCs in children and
(54). These cells are almost absent in healthy adults unless younger adults (70). These immature phenotype cell pop-
under certain physiological (e.g., pregnancy) or pathological ulations exhibit remarkable immunosuppressive properties by
conditions (chronic infections and cancer) (55). CD71+ ery- downregulating proinflammatory cytokine production and
throid cells (CECs) exhibit immunosuppressive properties T cell proliferation (71). Therefore, the presence of innate
and prevent a hyperimmune reaction to pathogens and immune regulatory mechanism(s) in newborns and children
commensal microbes in early life (4, 54). CECs by the se- may protect them from the collateral damage associated with
cretion of multiple soluble factors such as arginase-2, TGF-b, infections.
reactive oxygen species, or cell–cell interactions suppress in-
nate and adaptive immune cells (54, 56–61). In particular, we The choice of effector immune response in newborns and children
have shown that CECs promote the induction of regulatory The immune system not only identifies the infecting pathogen
T cells (Tregs) (61). Also, other groups have reported the (e.g., a bacterium versus a virus or a fungus) but also selects the
abundance of another subset of suppressor cells, myeloid- immune and energetic cost of its response (72). Any infection
derived suppressor cells (MDSCs), in infants (62, 63). The results in symptoms that are either directly associated with the
physiological abundance of certain immunosuppressive cells pathogen-imposed damage or with an excessive immune re-
supports the existence of a highly regulated immune system in sponse. The general concept is that the regulatory mechanisms
early life. The physiological abundance of these regulatory modulate the immune response, resulting in optimal immune
cells can reduce morbidity and mortality in neonates (16). For response in terms of magnitude and duration with minimal
example, it has been reported that neonates are more resistant immunopathology (5). However, under different circum-
to LPS challenge than adult mice. LPS/D-GalN induces liver stances, an excessive immune response can result in collateral
injury and 100% mortality in adult mice within 12 h, whereas damage and immunopathology. Not only the quantity but
it spares newborns (16). Studies also report a greater bacterial also the quality of immune response can result in different
load in newborns infected with different pathogens, which immunopathological consequences. It appears that our im-
does not seem unpredictable at first glance, because we believe mune system deploys different effector responses based on the
infants are more susceptible to infections than adults. As such, effector/cost ratio. The idea is that the low-cost response is
a higher bacterial load would appear to be normal. For in- generated first; if it fails in the clearance of infection, the ef-
stance, in the case of sepsis in adults, a bacterial load of 1–30 fector response associated with the next lowest cost is gener-
CFU/ml blood has commonly been reported, whereas in ated on a spectrum defined by the immunopathology cost
neonates this is a much higher 50–500 CFUs, and in one- (72). Sensing of microbial pathogen-associated molecular
third of infected newborns, this exceeds 1000 CFUs (12, 13). patterns by macrophages and dendritic cells in the absence of4 BRIEF REVIEWS: IMMUNE SYSTEM AND COVID-19 INFECTION IN THE YOUNG
immune regulation may induce higher cost effector responses four infants had a fever, two had very mild upper respiratory
in newborns. For example, the response of macrophages and symptoms, one had no symptoms, and for the last two, no
dendritic cells to pathogen-associated molecular patterns in information was provided (76). In another report, an infant
the presence of CECs generates an effector response of lower- (36 h postpartum) developed a case of neonatal COVID-19
cost, reduced microbicidal activity and immunopathology infection from an infected mother; however, the newborn did
(54, 57). Therefore, the presence of immunosuppression in not present any respiratory symptoms (77). Another study
the newborn/infant probably calibrates based on the weighted reported a total of 10 confirmed COVID-19 children ad-
contribution of different costs for the host. An interesting mitted to different hospitals in China with mild clinical
model has been proposed by Iwasaki and Medzhitov (72), symptoms (78). Similarly, mild symptoms were reported for
illustrating immune effector responses that have different another two pediatric patients (79). Among .44,000 con-
costs to host fitness. For example, constitutive secretion of firmed cases of COVID-19 in China, 0.9 and 1.2% were aged
secretory IgA and antimicrobial peptides (73) is a low-cost 0–10 and 10–19 y, respectively (80). Overall, children appear
protective mechanism. When this mechanism is unable to to exhibit milder symptoms following infection with
protect the host from pathogens, the next lowest-cost immune COVID-19 compared with adults, and in most cases, they are
response (e.g., tissue-resident macrophages) will be induced. asymptomatic (81, 82). Another study categorized the clinical
Recruitment of neutrophils and other immune cells to the site symptoms of 41 admitted hospital patients in Wuhan, but
of infection is the next lowest-cost immune response, followed none of them were infants (83). The largest nationwide pe-
by Th1, cytotoxic T cell response, and Th17. The immune diatric study in China reported 2143 patients with respiratory
defense against life-threatening pathogens is energetically ex- symptoms similar to COVID-19; however, only 34.1% of
pensive, and there is a trade-off between investment in a rapid them were laboratory-confirmed cases. Although this study
immune response versus growth and development in the reported that the disease was milder in infants than adults, it
infants/children (74). Therefore, the selection of the type suggested that infants are also susceptible to the virus. How-
of effector immune response in newborns/children should ever, the major limitation of this report is that more severe
be energetically less costly considering the high energy cost and critical cases were in the suspected group than in the
for growth and development (75). Therefore, these different laboratory-confirmed group, which suggests that these severe
circumstances provide reasoning for the adaptation of infec- cases might have been caused by other respiratory pathogens
tion tolerance versus infection resistance in newborns/infants but not COVID-19 (84). Similarly, in the United States,
(Fig. 1). among ∼150,000 confirmed cases of COVID-19 for which
age was noted, children ,18 y old accounted for 1.7 of cases
Infants, children, and younger adults are spared from COVID-19 with only three deaths (85). This report supports previous
Pediatric-specific risk factors for COVID-19 are not defined, data that COVID-19 cases in children present mild disease,
and there is no clear reason to date explaining why infants, but those asymptomatic children can play an important role
children, and younger adults are at reduced risk of developing in the spread of the virus in the community (85). These ob-
COVID-19 once infected with SARS-CoV-2. Newborns and servations are similar to previous outbreaks of severe acute
infants infected with SARS-CoV-2 (COVID-19) may benefit respiratory syndrome (SARS) and Middle East respiratory
from their biased immune tolerance phenotype. Despite syndrome (MERS), which resulted in high mortality in adults
thousands of reported cases of infection in China (.20,000 but none in pediatrics (86, 87). Thus far, evidence is
cases by February 6, 2020), only nine infants (0.05%) were emerging that COVID-19 infects infants and children with
hospitalized with COVID-19 (76). Among admitted cases, fairly high viral titer but they do not exhibit serious mani-
festations of the disease, and no mortality has been reported
for the pediatric age group (88). Given the number of
reported infections in adults, the number of infected infants
and children is miniscule. More importantly, infected infants
are mainly asymptomatic or exhibit very mild symptoms (76).
A similar phenomenon has been reported with the original
SARS coronavirus in mice. This study demonstrated that
young mice were able to clear the virus without significant
clinical symptoms, whereas all infected adult mice died
(89). This suggest that the differential immune response to
COVID-19 in adults versus infants may lead to a differential
clinical outcome.
Based on the collected preliminary data in COVID-19
patients, cytokine storm may play a role in enhanced disease
pathogenesis. For instance, elevated levels of a variety of cy-
tokines, such as IL-1b, IL-1RA, IFN-g, IP10, IL-7, IL-8,
IL-9, IL-10, GCSF, GMCSF, MIP1A, PDGF, FGF, VEGF,
TNF-a, and MCP1, were detected in plasma of patients as
compared with healthy adult individuals (83). In particular,
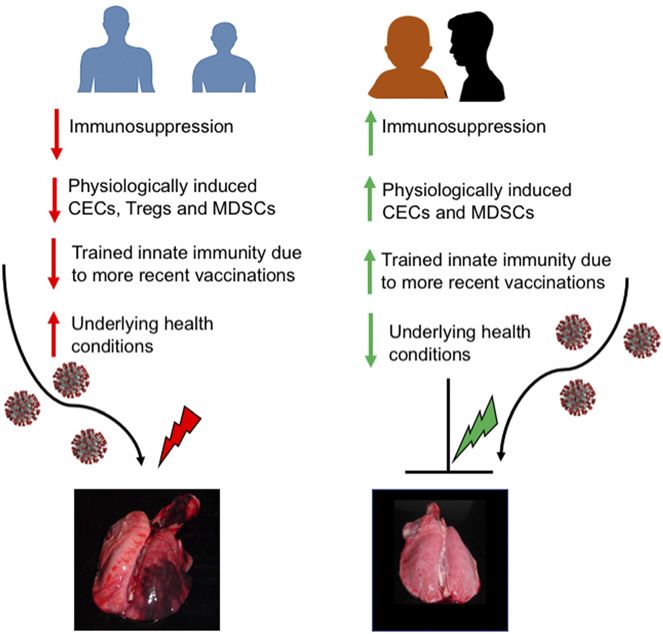
FIGURE 1. This diagram illustrates that a resistance mechanism to a admitted patients to the intensive care unit had elevated levels
pathogen in adults leads to collateral damage, but a tolerance mechanism to a of IL-2, IL-7, IL-10, IP10, MCP1, MIP1A, GCSF, and
pathogen in infants reduces such damage. TNF-a compared with those who did not require the intensiveThe Journal of Immunology 5 care unit (83). This preliminary evidence suggests that milder respiratory symptoms in infants and children. How- cytokine storm might be associated with disease severity. ever, further studies are urgently needed to test such opinions. Despite the elevated levels of cytokines that can enhance Th1-mediated immune response, IL-4 and IL-10 were also COVID-19 patients may benefit from immunosuppression higher in COVID-19 patients (83). This is in contrast to The uncontrolled inflammatory response can itself damage the the abundance of only proinflammatory cytokines IL-1b, lungs via the excessive release of proteases, reactive oxygen IFN-g, IP10, IL-12, IL-6, and MCP1 in SARS and IL-15, Il-17, species, and proinflammatory cytokines (98). In addition to IFN-g, and TNF-a in MERS corona virus infections, which localized damage, cytokine storm can have detrimental effects were associated with pulmonary inflammation and excessive on different organs and tissues. Elevated levels of cytokines lung damage (90, 91). However, there is no evidence for the such as TNF-a and IL-6 can promote septic shock syndrome cytokine levels in the plasma and respiratory tract of infected and collateral damage (98). In agreement with this concept, infants and children with COVID-19 compared with adults. several immunosuppressive strategies are recommended for Although such preliminary data indicate a role for cytokine the treatment of COVID-19 patients. Current evidence in storm in disease pathogenesis, further studies are necessary to patients with MERS and SARS suggests that corticosteroids confirm these observations. In view of the elevated amount of did not influence the mortality rate but delayed viral clearance proinflammatory cytokines induced by SARS, MERS, and (99, 100). Among a cohort of 41 COVID-19 patients, sys- COVID-19 infections, mild infection symptoms in infants temic corticosteroids were given to some patients without a may be related to the immunotolerance mechanisms during clear analysis of the outcome (83). Although the administra- early life. As such, physiological abundance of CECs and tion of systemic corticosteroids for COVID-19 patients is not MDSCs may limit excessive inflammation in response to supported by the World Health Organization guidelines (83), SARS-CoV-2 infection in infants. It is worth noting that several trials are under way for the efficacy of such treatment newborns, children, and younger adults (,18 y old) differ options (98). It is worth noting that glucocorticoids (e.g., from the immunological standpoint. Therefore, different dexamethasone, prednisone) exhibit differential effects that potential mechanisms may explain the drastic difference in should be taken into consideration (101). Nevertheless, a their mortality rates following COVID-19 infection com- most recent randomized clinical trial has shown that dexa- pared with adults (92). Although newborns/infants may methasone reduced deaths by one-third in patients on venti- benefit from the physiological abundance of immunosup- lators and by one-fifth in those receiving oxygen without pressive cells, these cells gradually disappear with age (4, 55). invasive ventilation (P. Horby, W. S. Lim, J. Emberson, Therefore, other unknown factors may account for the milder M. Mafham, J. Bell, L. Linsell, N. Staplin, C. Brightling, or asymptomatic infection in older children. For example, a A. Ustianowski, E. Elmahi, B. Prudon, C. Green, T. Felton, recent report has hypothesized that the trained nonspecific D. Chadwick, K. Rege, C. Fegan, L. C. Chappell, S. N. Faust, innate immunity following vaccination with live attenuated T. Jaki, K. Jeffery, A. Montgomery, K. Rowan, E. Juszczak, vaccines such as measles, mumps, and rubella (MMR) could J. K. Baillie, R. Haynes, and M. J. Landray, manuscript posted protect adolescents against COVID-19 (93). Thus, live at- on medRxiv). This study suggests that the immunomodulatory tenuated vaccines such as MMR, rotavirus, smallpox, and role of dexamethasone may prevent progression to respiratory bacillus Calmette-Guérin may provide nonspecific protective failure and death. Besides, there is another possibility that immunity (94). Another possibility is that such nonspecific dexamethasone (because of its immune-mediated effects) can effects translate into the induction of long-lived MDSCs (70). influence hematopoiesis (102), which merits further investi- Therefore, MDSC-mediated immunosuppression diminishes gations. Besides, multiple clinical trials are testing the efficacy any excessive inflammatory response to pathogens that can be of neutralizing IL-6, GM-CSF, TNF-a, and adjuvant therapy protective because SARS, MERS, and COVID-19 are all as- to absorb a broad range of cytokines (98). Overall these ob- sociated with severe pulmonary inflammation and sepsis (95). servations indicate a beneficial role for the immunosuppres- In support of their claim, the milder symptoms observed in sion in patients infected with SARS-CoV-2 virus. the 955 sailors in the U.S.S. Roosevelt who tested positive for COVID-19 might be related to MMR vaccination, which is COVID-19 entry in adults versus children commonly given to all Navy recruits (93). The role of Tregs The angiotensin-converting enzyme 2 (ACE2) is expressed in also needs to be taken into consideration because the age- many organs, such as the heart, lungs, intestine, and kidneys related thymic involution can lead to a decreased output of (103). SARS-CoV-2 binds to the ACE2 receptor for cell entry Tregs after the age of 50 (96). This decline in Treg frequency (103, 104) and not only gains initial entry through ACE2 but and function may contribute to age-related increased in- also downregulates ACE2 expression on cell surface such that flammation and autoimmune diseases (97). Therefore, the this enzyme cannot exert its protective role in organs (105). presence of a regulatory mechanism mediated by different Downregulation of ACE2 in the respiratory tract is linked to immunomodulatory cells may serve as a beneficial character- neutrophil infiltration that may result in angiotensin II ac- istic in the immune systems of younger populations. How- cumulation and lung injury (106, 107). Similarly, ACE2 ever, there is an urgent need to evaluate how the immune expression was found to be associated with enhanced sus- system in younger populations (as compared with older ceptibility to SARS-CoV infection (108). Although this as- populations) responds to COVID-19. We suggest that the sociation has been reported in vitro, the correlation of ACE2 tightly regulated nature of the neonate’s microenvironment expression and susceptibility to SARS-CoV-2 remains unknown can prevent the observed proinflammatory cytokine storm (109). Retrospective studies on confirmed cases of COVID-19 in adults as reported in other models (4, 57, 58). Therefore, found that children are spared from the disease (110). Although such differences may explain the mechanism underlying such the mechanism(s) underlying these observations is unknown,
6 BRIEF REVIEWS: IMMUNE SYSTEM AND COVID-19 INFECTION IN THE YOUNG
a differential expression of ACE2 in infants and children
compared with adults has been suggested (111). However,
there is no clear evidence showing the impact of age on ACE2
expression in the lungs except a recent study that has reported
that there was no difference in the activity of ACE2 in
bronchoalveolar lavage fluids of neonates, children, and adults
with acute respiratory distress syndrome (112). Another
supportive evidence for the differential ACE2 expression in
young versus old is the decline of androgens and estrogens by
age. Both these hormones upregulate ACE2 expression; thus,
their reduced production by age can result in the downreg-
ulation of ACE2 expression (113). The activation of the
renin-angiotensin system and the downregulation of ACE2
are reported to be involved in lung injury following SARS-
CoV infection (105). Animal studies indicate that the reduced
ACE2 activity or the loss of ACE2 results in neutrophil re-
cruitment, enhanced vascular permeability, and exaggerated
pulmonary edema, but in turn, supplementation of exogenous
ACE2 reverses this inflammatory response (114). Interest-
ingly, it has been reported that SARS-CoV infection down-
regulates ACE2 in lung tissues of mice, accompanied by lung FIGURE 2. This is a proposed model that illustrates the immunological
injury (105). Considering the similarities of SARS-CoV and differences between children and adults relative to COVID-19. Unlike adults,
SARS-CoV-2, there is a possibility that SARS-CoV-2 also infants/children have an immunosuppressive microenvironment, phys-
downregulates the ACE2 expression, resulting in pathology iologically enriched with immunomodulatory cells (e.g., CECs, Tregs,
and lung injury (109). The role of ACE2 appears to be and MDSCs), and because of the frequent live attenuated vaccines have
complex; on the one hand, it may increase the risk of SARS- more MDSCs but limited underlying health conditions. A green light-
ning bolt indicates a regulatory signal and the red lightning bolt shows
CoV-2 infection as the viral entry to the cell, but on the other an inflammatory signal.
hand, it can reduce lung injury following the infection. Al-
though differential tissue expression of ACE2 in children
receptors in the lungs and other organs/tissues of adults versus
versus adults needs to be determined, higher levels of plasma
infants and children. Thus, the expression pattern of ACE2
ACE2 in children have already been reported (115). There-
and SARS-CoV-2 coreceptor (the transmembrane protease
fore, further studies are required to investigate the expression
serine 2 [TMPRSS2]) (104) in adults versus children needs to
level of ACE2 in children versus adults to determine its po-
be examined. It is possible to speculate that the differential
tential role in COVID-19 infection.
expression of ACE2 and/or TMPRSS2 in children versus
adults could explain different infection outcomes. A full
Conclusions picture of the crucial host immune factors that contribute to
Emerging evidence indicates that although infants, children, the development of severe disease in some patients and milder
and adolescents are susceptible to infection with COVID-19, disease in infants and children remains poorly understood.
only a few have serious symptoms. Most infected infants are We speculate that the unique immune compartment in
asymptomatic but can act as a source of viral transmission. The children and the underlying health conditions in adults may
mechanism(s) underlying circumvention from severe form of partially explain the mild or asymptomatic disease observed in
the disease in the young (and, in particular, infants and this group of children (Fig. 2). Finally, the nonspecific trained
children) is unclear. Based on recent developments in the innate immunity may provide another explanation for the
neonatal immunology field (1, 4, 5, 54, 75), the choice of milder disease in adolescents. Therefore, further studies of the
immune response against COVID-19 in children and ado- host immune response to SARS-CoV-2 are warranted, in-
lescents versus adults may serve as an explanation for the cluding a detailed investigation of the determinants of healthy
observed differential clinical outcomes. Infants may benefit versus dysfunctional immune response to address our hy-
from the physiological abundance of immunoregulatory cells pothesis and better understand the immune correlates of
and having a tightly regulated immune system. There is protection in the younger population compared with adults.
compelling evidence for the innate immune hyperactivation
in driving the acute disease in SARS-CoV-2–infected adults.
However, the differential immune components in the young Acknowledgments
I thank Alexandria Nike Gardner for the helpful suggestions and edits.
may prevent excessive and potentially damaging immune re-
sponses to COVID-19 infection. Controlling the inflamma-
tory response to the virus may be as crucial as targeting the Disclosures
virus, as the unrestrained proinflammatory response in some The author has no financial conflicts of interest.
adults can itself result in immune cell infiltration, which
triggers a cytokine storm that mediates lung inflammation References
and injury. Of course, this is just one potential mechanism as 1. Elahi, S. 2014. New insight into an old concept: role of immature erythroid cells in
immune pathogenesis of neonatal infection. Front. Immunol. 5: 376.
other mechanisms are likely involved. Another potential 2. Adkins, B. 2000. Development of neonatal Th1/Th2 function. Int. Rev. Immunol.
mechanism might be the expression level of COVID-19 19: 157–171.The Journal of Immunology 7
3. Adkins, B., C. Leclerc, and S. Marshall-Clarke. 2004. Neonatal adaptive immunity 33. Principi, N., A. Zampiero, M. Gambino, A. Scala, L. Senatore, M. Lelii,
comes of age. Nat. Rev. Immunol. 4: 553–564. B. Ascolese, C. Pelucchi, and S. Esposito. 2015. Prospective evaluation of rhino-
4. Elahi, S. 2019. Neglected cells: immunomodulatory roles of CD71 + erythroid virus infection in healthy young children. J. Clin. Virol. 66: 83–89.
cells. Trends Immunol. 40: 181–185. 34. Falsey, A. R., and E. E. Walsh. 2000. Respiratory syncytial virus infection in
5. Kollmann, T. R., B. Kampmann, S. K. Mazmanian, A. Marchant, and O. Levy. adults. Clin. Microbiol. Rev. 13: 371–384.
2017. Protecting the newborn and young infant from infectious diseases: lessons 35. Nam, H. H., and M. G. Ison. 2019. Respiratory syncytial virus infection in adults.
from immune ontogeny. Immunity 46: 350–363. BMJ 366: l5021.
6. Read, A. F., A. L. Graham, and L. Råberg. 2008. Animal defenses against infec- 36. Lambert, L., A. M. Sagfors, P. J. Openshaw, and F. J. Culley. 2014. Immunity to
tious agents: is damage control more important than pathogen control. PLoS Biol. RSV in early-life. Front. Immunol. 5: 466.
6: e4. 37. Resch, B. 2012. Burden of respiratory syncytial virus infection in young children.
7. Behnke, J. M., C. J. Barnard, and D. Wakelin. 1992. Understanding chronic World J. Clin. Pediatr. 1: 8–12.
nematode infections: evolutionary considerations, current hypotheses and the way 38. McNamara, P. S., B. F. Flanagan, A. M. Selby, C. A. Hart, and R. L. Smyth.
forward. Int. J. Parasitol. 22: 861–907. 2004. Pro- and anti-inflammatory responses in respiratory syncytial virus bron-
8. Medzhitov, R., D. S. Schneider, and M. P. Soares. 2012. Disease tolerance as a chiolitis. Eur. Respir. J. 23: 106–112.
defense strategy. Science 335: 936–941. 39. Moreno-Solı́s, G., J. Torres-Borrego, M. J. de la Torre-Aguilar, F. Fernández-
9. Ayres, J. S., and D. S. Schneider. 2012. Tolerance of infections. Annu. Rev. Gutiérrez, F. J. Llorente-Cantarero, and J. L. Pérez-Navero. 2015. Analysis of the
Immunol. 30: 271–294. local and systemic inflammatory response in hospitalized infants with respiratory
10. Graham, A. L., J. E. Allen, and A. F. Read. 2005. Evolutionary causes and con- syncitial virus bronchiolitis. Allergol. Immunopathol. (Madr.) 43: 264–271.
sequences of immunopathology. Annu. Rev. Ecol. Evol. Syst. 36: 373–397. 40. Reimers, K., K. Buchholz, and H. Werchau. 2005. Respiratory syncytial virus M2-
11. Casadevall, A., and L. A. Pirofski. 1999. Host-pathogen interactions: redefining 1 protein induces the activation of nuclear factor kappa B. Virology 331: 260–268.
the basic concepts of virulence and pathogenicity. Infect. Immun. 67: 3703–3713. 41. Hornsleth, A., B. Klug, M. Nir, J. Johansen, K. S. Hansen, L. S. Christensen, and
12. Hall, W. H., and D. Gold. 1955. Shock associated with bacteremia; review of L. B. Larsen. 1998. Severity of respiratory syncytial virus disease related to type and
thirty-five cases. AMA Arch. Intern. Med. 96: 403–412. genotype of virus and to cytokine values in nasopharyngeal secretions. Pediatr.
13. Yagupsky, P., and F. S. Nolte. 1990. Quantitative aspects of septicemia. Clin. Infect. Dis. J. 17: 1114–1121.
Microbiol. Rev. 3: 269–279. 42. Blanco, J. C., J. Y. Richardson, M. E. Darnell, A. Rowzee, L. Pletneva,
14. Brook, B., D. Harbeson, R. Ben-Othman, D. Viemann, and T. R. Kollmann. D. D. Porter, and G. A. Prince. 2002. Cytokine and chemokine gene expression
2017. Newborn susceptibility to infection vs. disease depends on complex in vivo after primary and secondary respiratory syncytial virus infection in cotton rats. J.
interactions of host and pathogen. Semin. Immunopathol. 39: 615–625. Infect. Dis. 185: 1780–1785.
15. Hornik, C. P., D. K. Benjamin, K. C. Becker, D. K. Benjamin, Jr., J. Li, 43. Johnson, J. E., R. A. Gonzales, S. J. Olson, P. F. Wright, and B. S. Graham. 2007.
R. H. Clark, M. Cohen-Wolkowiez, and P. B. Smith. 2012. Use of the The histopathology of fatal untreated human respiratory syncytial virus infection.
complete blood cell count in early-onset neonatal sepsis. Pediatr. Infect. Dis. J. Mod. Pathol. 20: 108–119.
31: 799–802. 44. Goulder, P. J., S. R. Lewin, and E. M. Leitman. 2016. Paediatric HIV infection:
16. Zhao, J., K. D. Kim, X. Yang, S. Auh, Y. X. Fu, and H. Tang. 2008. Hyper innate the potential for cure. Nat. Rev. Immunol. 16: 259–271.
responses in neonates lead to increased morbidity and mortality after infection. 45. Paul, M. E., C. Mao, M. Charurat, L. Serchuck, M. Foca, K. Hayani,
Proc. Natl. Acad. Sci. USA 105: 7528–7533. E. L. Handelsman, C. Diaz, K. McIntosh, and W. T. Shearer; Women and Infants
17. Yim, H. J., and A. S. F. Lok. 2006. Natural history of chronic hepatitis B virus Transmission Study. 2005. Predictors of immunologic long-term nonprogression
infection: what we knew in 1981 and what we know in 2005. Hepatology 43(Suppl. in HIV-infected children: implications for initiating therapy. J. Allergy Clin.
1): S173–S181. Immunol. 115: 848–855.
18. Vogt, M., T. Lang, G. Frösner, C. Klingler, A. F. Sendl, A. Zeller, B. Wiebecke, 46. Crux, N. B., and S. Elahi. 2017. Human leukocyte antigen (HLA) and immune
B. Langer, H. Meisner, and J. Hess. 1999. Prevalence and clinical outcome of regulation: how do classical and non-classical HLA alleles modulate immune re-
hepatitis C infection in children who underwent cardiac surgery before the sponse to human immunodeficiency virus and hepatitis C virus infections? Front.
implementation of blood-donor screening. N. Engl. J. Med. 341: 866–870. Immunol. 8: 832.
19. Sasaki, N., A. Matsui, M. Momoi, F. Tsuda, and H. Okamoto. 1997. Loss of 47. Elahi, S., W. L. Dinges, N. Lejarcegui, K. J. Laing, A. C. Collier, D. M. Koelle,
circulating hepatitis C virus in children who developed a persistent carrier state M. J. McElrath, and H. Horton. 2011. Protective HIV-specific CD8+ T cells
after mother-to-baby transmission. Pediatr. Res. 42: 263–267. evade Treg cell suppression. [Published erratum appears in 2011 Nat. Med. 17:
20. Lutckii, A., B. Strunz, A. Zhirkov, O. Filipovich, E. Rukoiatkina, D. Gusev, 1153.] Nat. Med. 17: 989–995.
Y. Lobzin, B. Fischler, S. Aleman, M. Sällberg, and N. K. Björkström. 2020. 48. Elahi, S., and H. Horton. 2012. Association of HLA-alleles with the immune
Evidence for B cell maturation but not trained immunity in uninfected infants regulation of chronic viral infections. Int. J. Biochem. Cell Biol. 44: 1361–1365.
exposed to hepatitis C virus. Gut. DOI: 10.1136/gutjnl-2019-320269. 49. Migueles, S. A., A. C. Laborico, W. L. Shupert, M. S. Sabbaghian, R. Rabin,
21. Babik, J. M., D. Cohan, A. Monto, D. J. Hartigan-O’Connor, and J. M. McCune. C. W. Hallahan, D. Van Baarle, S. Kostense, F. Miedema, M. McLaughlin, et al.
2011. The human fetal immune response to hepatitis C virus exposure in utero. J. 2002. HIV-specific CD8+ T cell proliferation is coupled to perforin expression and
Infect. Dis. 203: 196–206. is maintained in nonprogressors. Nat. Immunol. 3: 1061–1068.
22. Carvalho-Queiroz, C., M. A. Johansson, J. O. Persson, E. Jörtsö, T. Kjerstadius, 50. Adland, E., P. Paioni, C. Thobakgale, L. Laker, L. Mori, M. Muenchhoff,
C. Nilsson, S. Saghafian-Hedengren, and E. Sverremark-Ekström. 2016. Associ- A. Csala, M. Clapson, J. Flynn, V. Novelli, et al. 2015. Discordant impact of HLA
ations between EBV and CMV seropositivity, early exposures, and gut microbiota on viral replicative capacity and disease progression in pediatric and adult HIV
in a prospective birth cohort: a 10-year follow-up. Front Pediatr. 4: 93. infection. PLoS Pathog. 11: e1004954.
23. Blumental, S., and P. Lepage. 2019. Management of varicella in neonates and 51. Muenchhoff, M., E. Adland, O. Karimanzira, C. Crowther, M. Pace, A. Csala,
infants. BMJ Paediatr. Open 3: e000433. E. Leitman, A. Moonsamy, C. McGregor, J. Hurst, et al. 2016. Nonprogressing
24. Pass, R. F., S. Stagno, W. J. Britt, and C. A. Alford. 1983. Specific cell-mediated HIV-infected children share fundamental immunological features of nonpatho-
immunity and the natural history of congenital infection with cytomegalovirus. J. genic SIV infection. Sci. Transl. Med. 8: 358ra125.
Infect. Dis. 148: 953–961. 52. Silvestri, G., D. L. Sodora, R. A. Koup, M. Paiardini, S. P. O’Neil,
25. Huygens, A., N. Dauby, D. Vermijlen, and A. Marchant. 2014. Immunity to H. M. McClure, S. I. Staprans, and M. B. Feinberg. 2003. Nonpathogenic SIV
cytomegalovirus in early life. Front. Immunol. 5: 552. infection of sooty mangabeys is characterized by limited bystander immunopa-
26. Slavuljica, I., D. Kveštak, P. C. Huszthy, K. Kosmac, W. J. Britt, and S. Jonjić. thology despite chronic high-level viremia. Immunity 18: 441–452.
2015. Immunobiology of congenital cytomegalovirus infection of the central 53. Yu, J. C., H. Khodadadi, A. Malik, B. Davidson, É. D. S. L. Salles, J. Bhatia,
nervous system—the murine cytomegalovirus model. Cell. Mol. Immunol. 12: V. L. Hale, and B. Baban. 2018. Innate immunity of neonates and infants. Front.
180–191. Immunol. 9: 1759.
27. Dunmire, S. K., K. A. Hogquist, and H. H. Balfour. 2015. Infectious mononu- 54. Elahi, S., J. M. Ertelt, J. M. Kinder, T. T. Jiang, X. Zhang, L. Xin, V. Chaturvedi,
cleosis. Curr. Top. Microbiol. Immunol. 390: 211–240. B. S. Strong, J. E. Qualls, K. A. Steinbrecher, et al. 2013. Immunosuppressive
28. Merlo, A., R. Turrini, R. Dolcetti, D. Martorelli, E. Muraro, P. Comoli, and CD71+ erythroid cells compromise neonatal host defence against infection. Nature
A. Rosato. 2010. The interplay between Epstein-Barr virus and the immune sys- 504: 158–162.
tem: a rationale for adoptive cell therapy of EBV-related disorders. Haematologica 55. Elahi, S., and S. Mashhouri. 2020. Immunological consequences of extramedullary
95: 1769–1777. erythropoiesis: immunoregulatory functions of CD71+ erythroid cells. Haemato-
29. Ferressini Gerpe, N. M., A. G. Vistarop, A. Moyano, E. De Matteo, logica 105: 1478–1483.
M. V. Preciado, and P. A. Chabay. 2020. Distinctive EBV infection characteristics 56. Delyea, C., N. Bozorgmehr, P. Koleva, G. Dunsmore, S. Shahbaz, V. Huang, and
in children from a developing country. Int. J. Infect. Dis. 93: 139–145. S. Elahi. 2018. CD71+ erythroid suppressor cells promote fetomaternal tolerance
30. Jayasooriya, S., T. I. de Silva, J. Njie-jobe, C. Sanyang, A. M. Leese, A. I. Bell, through arginase-2 and PDL-1. J. Immunol. 200: 4044–4058.
K. A. McAulay, P. Yanchun, H. M. Long, T. Dong, et al. 2015. Early virological 57. Dunsmore, G., N. Bozorgmehr, C. Delyea, P. Koleva, A. Namdar, and S. Elahi.
and immunological events in asymptomatic Epstein-Barr virus infection in African 2017. Erythroid suppressor cells compromise neonatal immune response against
children. PLoS Pathog. 11: e1004746. Bordetella pertussis. J. Immunol. 199: 2081–2095.
31. Nader, S., R. Bergen, M. Sharp, and A. M. Arvin. 1995. Age-related differences in 58. Dunsmore, G., P. Koleva, N. Ghobakhloo, R. T. Sutton, L. Ambrosio, X. Meng,
cell-mediated immunity to varicella-zoster virus among children and adults im- N. Hotte, V. Nguyen, K. L. Madsen, L. A. Dieleman, et al. 2018. Lower abun-
munized with live attenuated varicella vaccine. J. Infect. Dis. 171: 13–17. dance and impaired function of CD71+ erythroid cells in inflammatory bowel
32. Esposito, S., C. Daleno, G. Prunotto, A. Scala, C. Tagliabue, I. Borzani, E. Fossali, disease patients during pregnancy. J Crohns Colitis 13: 230–244.
C. Pelucchi, and N. Principi. 2013. Impact of viral infections in children with 59. Namdar, A., G. Dunsmore, S. Shahbaz, P. Koleva, L. Xu, J. Jovel, S. Houston, and
community-acquired pneumonia: results of a study of 17 respiratory viruses. In- S. Elahi. 2019. CD71 + erythroid cells exacerbate HIV-1 susceptibility, mediate
fluenza Other Respir. Viruses 7: 18–26. trans-infection, and harbor infective viral particles. mBio 10: e02767-19.8 BRIEF REVIEWS: IMMUNE SYSTEM AND COVID-19 INFECTION IN THE YOUNG
60. Namdar, A., P. Koleva, S. Shahbaz, S. Strom, V. Gerdts, and S. Elahi. 2017. 89. Day, C. W., R. Baric, S. X. Cai, M. Frieman, Y. Kumaki, J. D. Morrey, D. F. Smee,
CD71 + erythroid suppressor cells impair adaptive immunity against Bordetella and D. L. Barnard. 2009. A new mouse-adapted strain of SARS-CoV as a lethal
pertussis. Sci. Rep. 7: 7728. model for evaluating antiviral agents in vitro and in vivo. Virology 395: 210–222.
61. Shahbaz, S., N. Bozorgmehr, P. Koleva, A. Namdar, J. Jovel, R. A. Fava, and 90. Wong, C. K., C. W. Lam, A. K. Wu, W. K. Ip, N. L. Lee, I. H. Chan, L. C. Lit,
S. Elahi. 2018. CD71+VISTA+ erythroid cells promote the development and D. S. Hui, M. H. Chan, S. S. Chung, and J. J. Sung. 2004. Plasma inflammatory
function of regulatory T cells through TGF-b. PLoS Biol. 16: e2006649. cytokines and chemokines in severe acute respiratory syndrome. Clin. Exp.
62. Gantt, S., A. Gervassi, H. Jaspan, and H. Horton. 2014. The role of myeloid- Immunol. 136: 95–103.
derived suppressor cells in immune ontogeny. Front. Immunol. 5: 387. 91. Mahallawi, W. H., O. F. Khabour, Q. Zhang, H. M. Makhdoum, and
63. Gervassi, A., N. Lejarcegui, S. Dross, A. Jacobson, G. Itaya, E. Kidzeru, S. Gantt, B. A. Suliman. 2018. MERS-CoV infection in humans is associated with a pro-
H. Jaspan, and H. Horton. 2014. Myeloid derived suppressor cells are present at inflammatory Th1 and Th17 cytokine profile. Cytokine 104: 8–13.
high frequency in neonates and suppress in vitro T cell responses. PLoS One 9: 92. Liu, W., Q. Zhang, J. Chen, R. Xiang, H. Song, S. Shu, L. Chen, L. Liang,
e107816. J. Zhou, L. You, et al. 2020. Detection of covid-19 in children in early January
64. Dietzman, D. E., G. W. Fischer, and F. D. Schoenknecht. 1974. Neonatal 2020 in Wuhan, China. N. Engl. J. Med. 382: 1370–1371.
Escherichia coli septicemia--bacterial counts in blood. J. Pediatr. 85: 128–130. 93. Fidel, P. L., Jr., and M. C. Noverr. 2020. Could an unrelated live attenuated
65. Fitzpatrick, E. A., D. You, B. Shrestha, D. Siefker, V. S. Patel, N. Yadav, vaccine serve as a preventive measure to dampen septic inflammation associated
S. Jaligama, and S. A. Cormier. 2017. A neonatal murine model of MRSA with COVID-19 infection? mBio 11: e00907-20.
pneumonia. PLoS One 12: e0169273. 94. Moorlag, S. J. C. F. M., R. J. W. Arts, R. van Crevel, and M. G. Netea. 2019.
66. Filias, A., G. L. Theodorou, S. Mouzopoulou, A. A. Varvarigou, S. Mantagos, and Non-specific effects of BCG vaccine on viral infections. Clin. Microbiol. Infect. 25:
M. Karakantza. 2011. Phagocytic ability of neutrophils and monocytes in neo- 1473–1478.
nates. BMC Pediatr. 11: 29. 95. Zhou, F., T. Yu, R. Du, G. Fan, Y. Liu, Z. Liu, J. Xiang, Y. Wang, B. Song,
67. Zhang, Q., A. P. Coveney, S. Yu, J. H. Liu, Y. Li, S. Blankson, H. P. Redmond, X. Gu, et al. 2020. Clinical course and risk factors for mortality of adult inpatients
J. H. Wang, and J. Wang. 2013. Inefficient antimicrobial functions of innate with COVID-19 in Wuhan, China: a retrospective cohort study. [Published er-
phagocytes render infant mice more susceptible to bacterial infection. Eur. J. ratum appears in 2020 Lancet 395: 1038.] Lancet 395: 1054–1062.
Immunol. 43: 1322–1332. 96. Fessler, J., A. Ficjan, C. Duftner, and C. Dejaco. 2013. The impact of aging on
68. Silveira-Lessa, A. L., C. Quinello, L. Lima, A. C. C. Redondo, M. E. J. R. Ceccon, regulatory T-cells. Front. Immunol. 4: 231.
M. Carneiro-Sampaio, and P. Palmeira. 2016. TLR expression, phagocytosis and 97. Tsaknaridis, L., L. Spencer, N. Culbertson, K. Hicks, D. LaTocha, Y. K. Chou,
oxidative burst in healthy and septic newborns in response to Gram-negative and R. H. Whitham, A. Bakke, R. E. Jones, H. Offner, et al. 2003. Functional assay for
Gram-positive rods. Hum. Immunol. 77: 972–980. human CD4+CD25+ Treg cells reveals an age-dependent loss of suppressive ac-
69. Wynn, J. L., C. S. Wilson, J. Hawiger, P. O. Scumpia, A. F. Marshall, J. H. Liu, tivity. J. Neurosci. Res. 74: 296–308.
I. Zharkikh, H. R. Wong, P. Lahni, J. T. Benjamin, et al. 2016. Targeting IL-17A 98. Tay, M. Z., C. M. Poh, L. Renia, P. A. MacAry, and L. F. P. Ng. 2020. The trinity
attenuates neonatal sepsis mortality induced by IL-18. Proc. Natl. Acad. Sci. USA of COVID-19: immunity, inflammation and intervention. Nat. Rev. Immunol. 20:
113: E2627–E2635. 363–374.
70. Lilly, E. A., M. Ikeh, E. E. Nash, P. L. Fidel, Jr., and M. C. Noverr. 2018. Im- 99. Arabi, Y. M., Y. Mandourah, F. Al-Hameed, A. A. Sindi, G. A. Almekhlafi,
mune protection against lethal fungal-bacterial intra-abdominal infections. mBio 9: M. A. Hussein, J. Jose, R. Pinto, A. Al-Omari, A. Kharaba, et al; Saudi Critical
e01472-17. Care Trial Group. 2018. Corticosteroid therapy for critically ill patients with
71. Youn, J. I., and D. I. Gabrilovich. 2010. The biology of myeloid-derived sup- Middle East respiratory syndrome. Am. J. Respir. Crit. Care Med. 197: 757–767.
pressor cells: the blessing and the curse of morphological and functional hetero- 100. Stockman, L. J., R. Bellamy, and P. Garner. 2006. SARS: systematic review of
geneity. Eur. J. Immunol. 40: 2969–2975. treatment effects. PLoS Med. 3: e343.
72. Iwasaki, A., and R. Medzhitov. 2015. Control of adaptive immunity by the innate 101. Okoye, I. S., L. Xu, J. Walker, and S. Elahi. 2020. The glucocorticoids prednisone
immune system. Nat. Immunol. 16: 343–353. and dexamethasone differentially modulate T cell function in response to anti-PD-
73. Elahi, S., R. M. Buchanan, S. Attah-Poku, H. G. Townsend, L. A. Babiuk, and 1 and anti-CTLA-4 immune checkpoint blockade. Cancer Immunol. Immunother.
V. Gerdts. 2006. The host defense peptide beta-defensin 1 confers protection 69: 1423–1436.
against Bordetella pertussis in newborn piglets. Infect Immun. 74: 2338–2352. 102. Narla, A., S. Dutt, J. R. McAuley, F. Al-Shahrour, S. Hurst, M. McConkey,
74. McDade, T. W., V. Reyes-Garcı́a, S. Tanner, T. Huanca, and W. R. Leonard. D. Neuberg, and B. L. Ebert. 2011. Dexamethasone and lenalidomide have dis-
2008. Maintenance versus growth: investigating the costs of immune activation tinct functional effects on erythropoiesis. Blood 118: 2296–2304.
among children in lowland Bolivia. Am. J. Phys. Anthropol. 136: 478–484. 103. Yan, R., Y. Zhang, Y. Li, L. Xia, Y. Guo, and Q. Zhou. 2020. Structural basis for the
75. Harbeson, D., R. Ben-Othman, N. Amenyogbe, and T. R. Kollmann. 2018. recognition of SARS-CoV-2 by full-length human ACE2. Science 367: 1444–1448.
Outgrowing the immaturity myth: the cost of defending from neonatal infectious 104. Hoffmann, M., H. Kleine-Weber, S. Schroeder, N. Kr€ uger, T. Herrler,
disease. Front. Immunol. 9: 1077. S. Erichsen, T. S. Schiergens, G. Herrler, N. H. Wu, A. Nitsche, et al. 2020.
76. Wei, M., J. Yuan, Y. Liu, T. Fu, X. Yu, and Z. J. Zhang. 2020. Novel coronavirus SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a
infection in hospitalized infants under 1 year of age in China. JAMA 323: 1313–1314. clinically proven protease inhibitor. Cell 181: 271–280.e8.
77. Wang, S., L. Guo, L. Chen, W. Liu, Y. Cao, J. Zhang, and L. Feng. 2020. A case 105. Kuba, K., Y. Imai, S. Rao, H. Gao, F. Guo, B. Guan, Y. Huan, P. Yang, Y. Zhang,
report of neonatal 2019 coronavirus disease in China. Clin. Infect. Dis. 71: 853–857. W. Deng, et al. 2005. A crucial role of angiotensin converting enzyme 2 (ACE2) in
78. Cai, J., J. Xu, D. Lin, Z. Yang, L. Xu, Z. Qu, Y. Zhang, H. Zhang, R. Jia, P. Liu, SARS coronavirus-induced lung injury. Nat. Med. 11: 875–879.
et al. 2020. A case series of children with 2019 novel coronavirus infection: clinical 106. Gu, H., Z. Xie, T. Li, S. Zhang, C. Lai, P. Zhu, K. Wang, L. Han, Y. Duan,
and epidemiological features. Clin. Infect. Dis. DOI: 10.1093/cid/ciaa198. Z. Zhao, et al. 2016. Angiotensin-converting enzyme 2 inhibits lung injury in-
79. Ji, L. N., S. Chao, Y. J. Wang, X. J. Li, X. D. Mu, M. G. Lin, and R. M. Jiang. duced by respiratory syncytial virus. Sci. Rep. 6: 19840.
2020. Clinical features of pediatric patients with COVID-19: a report of two 107. Yang, P., H. Gu, Z. Zhao, W. Wang, B. Cao, C. Lai, X. Yang, L. Zhang, Y. Duan,
family cluster cases. World J. Pediatr. 16: 267–270. S. Zhang, et al. 2014. Angiotensin-converting enzyme 2 (ACE2) mediates influ-
80. Lee, P. I., Y. L. Hu, P. Y. Chen, Y. C. Huang, and P. R. Hsueh. 2020. Are children enza H7N9 virus-induced acute lung injury. Sci. Rep. 4: 7027.
less susceptible to COVID-19? J. Microbiol. Immunol. Infect. 53: 371–372. 108. Hofmann, H., M. Geier, A. Marzi, M. Krumbiegel, M. Peipp, G. H. Fey,
81. Cao, Q., Y. C. Chen, C. L. Chen, and C. H. Chiu. 2020. SARS-CoV-2 infection T. Gramberg, and S. Pöhlmann. 2004. Susceptibility to SARS coronavirus S
in children: transmission dynamics and clinical characteristics. J. Formos. Med. protein-driven infection correlates with expression of angiotensin converting en-
Assoc. 119: 670–673. zyme 2 and infection can be blocked by soluble receptor. Biochem. Biophys. Res.
82. Guan, W. J., Z. Y. Ni, Y. Hu, W. H. Liang, C. Q. Ou, J. X. He, L. Liu, H. Shan, Commun. 319: 1216–1221.
C. L. Lei, D. S. C. Hui, et al; China Medical Treatment Expert Group for Covid- 109. Guo, J., Z. Huang, L. Lin, and J. Lv. 2020. Coronavirus disease 2019 (COVID-
19. 2020. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. 19) and cardiovascular disease: a viewpoint on the potential influence of
Med. 382: 1708–1720. angiotensin-converting enzyme inhibitors/angiotensin receptor blockers on onset
83. Huang, C., Y. Wang, X. Li, L. Ren, J. Zhao, Y. Hu, L. Zhang, G. Fan, J. Xu, and severity of severe acute respiratory syndrome coronavirus 2 infection. J. Am.
X. Gu, et al. 2020. Clinical features of patients infected with 2019 novel coro- Heart Assoc. 9: e016219.
navirus in Wuhan, China. [Published erratum appears in 2020 Lancet 395: 496.] 110. Li, Q., X. Guan, P. Wu, X. Wang, L. Zhou, Y. Tong, R. Ren, K. S. M. Leung, E. H.
Lancet 395: 497–506. Y. Lau, J. Y. Wong, et al. 2020. Early transmission dynamics in Wuhan, China, of
84. Dong, Y., X. Mo, Y. Hu, X. Qi, F. Jiang, Z. Jiang, and S. Tong. 2020. Epide- novel coronavirus-infected pneumonia. N. Engl. J. Med. 382: 1199–1207.
miological characteristics of 2143 pediatric patients with 2019 coronavirus disease 111. Bhat, B. V., and S. Ravikumar. 2020. Challenges in neonatal COVID-19 infec-
in China. Pediatrics. DOI: 10.1542/peds.2020-0702. tion. Indian J. Pediatr. 87: 577–578.
85. CDC COVID-19 Response Team. 2020. Coronavirus disease 2019 in children - 112. Schouten, L. R., A. H. van Kaam, F. Kohse, F. Veltkamp, L. D. Bos, F. M. de
United States, February 12-April 2, 2020. MMWR Morb. Mortal Wkly. Rep. 69: Beer, R. T. van Hooijdonk, J. Horn, M. Straat, E. Witteveen, et al; MARS
422–426. consortium. 2019. Age-dependent differences in pulmonary host responses in
86. Lau, J. T., M. Lau, J. H. Kim, H. Y. Tsui, T. Tsang, and T. W. Wong. 2004. ARDS: a prospective observational cohort study. Ann. Intensive Care 9: 55.
Probable secondary infections in households of SARS patients in Hong Kong. 113. Cristiani, L., E. Mancino, L. Matera, R. Nenna, A. Pierangeli, C. Scagnolari, and
Emerg. Infect. Dis. 10: 235–243. F. Midulla. 2020. Will children reveal their secret? The coronavirus dilemma. Eur.
87. Memish, Z. A., J. A. Al-Tawfiq, A. Assiri, F. A. AlRabiah, S. Al Hajjar, Respir. J. 55: 2000749.
A. Albarrak, H. Flemban, R. F. Alhakeem, H. Q. Makhdoom, S. Alsubaie, and 114. Zhang, H., and A. Baker. 2017. Recombinant human ACE2: acing out angio-
A. A. Al-Rabeeah. 2014. Middle East respiratory syndrome coronavirus disease in tensin II in ARDS therapy. Crit. Care 21: 305.
children. Pediatr. Infect. Dis. J. 33: 904–906. 115. Bénéteau-Burnat, B., B. Baudin, G. Morgant, F. C. Baumann, and J. Giboudeau.
88. Lu, Q., and Y. Shi. 2020. Coronavirus disease (COVID-19) and neonate: what 1990. Serum angiotensin-converting enzyme in healthy and sarcoidotic children:
neonatologist need to know. J. Med. Virol. 92: 564–567. comparison with the reference interval for adults. Clin. Chem. 36: 344–346.You can also read

















































