Neolepetopsid true limpets (Gastropoda: Patellogastropoda) from Indian Ocean hot vents shed light on relationships among genera
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Zoological Journal of the Linnean Society, 2022, 194, 276–296. With 10 figures.
Neolepetopsid true limpets (Gastropoda: Patellogastropoda)
from Indian Ocean hot vents shed light on relationships
among genera
CHONG CHEN1,*, , YADONG ZHOU2,3*, HIROMI KAYAMA WATANABE1, ,
RUIYAN ZHANG2,4 and CHUNSHENG WANG2,3,4
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
1
X-STAR, Japan Agency for Marine-Earth Science and Technology (JAMSTEC), 2–15 Natsushima-cho,
Yokosuka 237-0061, Japan
2
Key Laboratory of Marine Ecosystem Dynamics, Second Institute of Oceanography, Ministry of Natural
Resources, Hangzhou, China
3
Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519082, China
4
School of Oceanography, Shanghai Jiao Tong University, Shanghai, China
Received 22 February 2021; accepted for publication 27 August 2021
Neolepetopsidae is a family of true limpets restricted to deep-sea chemosynthesis-based ecosystems. It is a small and
little-studied family with about a dozen species in three genera, namely Eulepetopsis, Neolepetopsis and Paralepetopsis,
and all named species were from the Pacific or Atlantic Oceans. Here, we describe three new species from Indian
Ocean vents, namely Eulepetopsis crystallina sp. nov. found across three ridges, Neolepetopsis ardua sp. nov.
from the Southwest Indian Ridge and Neolepetopsis prismatica sp. nov. from the Carlsberg Ridge. Given that
Neolepetopsis appears to specialize on inactive sulfide deposits, the apparent wider distribution of E. crystallina is
probably attributable to bias in sampling effort at inactive chimneys. The molecular phylogeny of Patellogastropoda,
reconstructed using the COI gene, supported the monophyly of Neolepetopsidae. These are the first molecular data
available for Neolepetopsis, confirming that the three genera are genetically distinct. Eulepetopsis appears to be
adapted to active vents, and its derived position compared with Paralepetopsis indicates a possible ‘stepping-stone’
evolutionary pathway from seeps and organic falls to vents. Our results provide new insights into this enigmatic
family and highlight the importance of surveying the vent periphery, especially given that inactive vents are being
eyed as a replacement for active ones in deep-sea mining.
ADDITIONAL KEYWORDS: deep sea – deep-sea vents – Mollusca – molecular phylogeny.
INTRODUCTION Nevertheless, there is a global bias in studying those
systems close to the developed counties in the Northern
Since the discovery of hydrothermal vents on
Hemisphere, leaving sites in the Southern Hemisphere
the Galápagos Rift in 1977 (Corliss et al., 1979),
and Indian Ocean poorly understood, especially in
chemosynthesis-based ecosystems have been a centre of
terms of biodiversity (Thaler & Amon, 2019). Vents in
focus in deep-sea explorations around the world. This is
the western Indian Ocean, eyed by multiple nations
driven, in part, by the high endemicity of animals living
for the extraction of massive sulfide deposits on the
in these habitats (Wolff, 2005; Chapman et al., 2019)
seafloor, still contain a wealth of undescribed species
attributable to their ‘extreme’ environmental conditions
(Copley et al., 2016; Zhou et al., 2018; Sun et al., 2020).
requiring special adaptations (Van Dover, 2000).
Many named species have been categorized formally
as threatened or endangered owing to extinction risks
from deep-sea mining, but those species without names
*Corresponding authors. E-mail: cchen@jamstec.go.jp; yadong_
cannot be protected effectively (Sigwart et al., 2019).
zhou@sio.org.cn
[Version of record, published online 22 October 2021; Gastropod molluscs are a major component of
http://zoobank.org/urn:lsid:zoobank.org:pub:6334CD15- biodiversity in chemosynthetic ecosystems, such as
D490-496F-802A-F162B4FF8A21] hydrothermal vents, in terms of both species richness
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296 276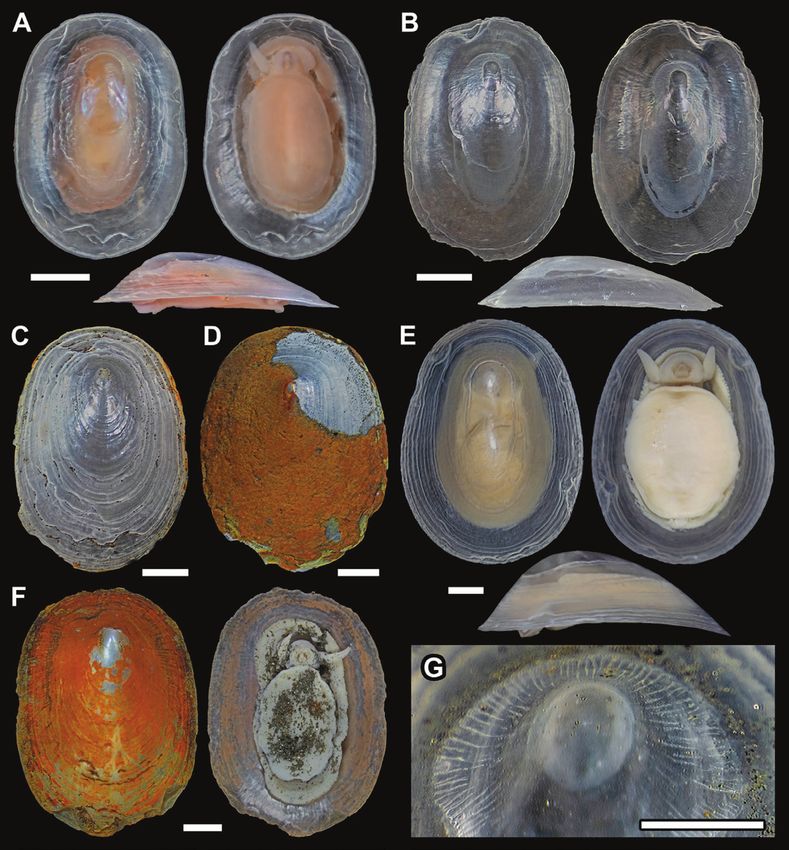
INDIAN OCEAN VENT LIMPETS 277
and abundance (Woff, 2005; Chapman et al., 2019). hydrocarbon seeps and a whale fall across the Pacific
Many families are endemic to these habitats, such and Atlantic Oceans (McLean, 1990, 2008; Beck, 1996;
as Peltospiridae, Lepetodrilidae and Neolepetopsidae Warén & Bouchet, 2001, 2009). In addition, at least two
(Sasaki et al., 2010). Of these, the limpet family undescribed species are known, one from cold seeps in
Neolepetopsidae is the only patellogastropod family the Gulf of Mexico (Aktipis & Giribet, 2010) and one
restricted to chemosynthesis-based ecosystems and is from both vents and seeps in the Gulf of California
unique among true limpets in having a radula with (Goffredi et al., 2017). It is distinguished from the other
articulate shafts while lacking basal plates (McLean, two genera in having an opaque shell with weaker
1990; Warén & Bouchet, 2001; McLean, 2008). The sculpture than Neolepetopsis, and a radula with broad
radula is variably mineralized depending on the cusps (McLean, 2008).
species and often less mineralized than in shallow- The phylogenetic position of Neolepetopsidae in
water patellogastropods, with amorphous phases or Patellogastropoda remains uncertain. An early study
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
being barely mineralized (Cruz & Farina, 2005; Warén using 18S rRNA gene fragments recovered it as sister
& Bouchet, 2009). The shell ranges from opaque to fully to Acmaeidae (Harasewych & McArthur, 2000), and
transparent (McLean, 1990). Shell microsculpture has later a different, more densely sampled study using
been described for all three genera, with Eulepetopsis the mitochondrial cytochrome oxidase c subunit I (COI)
McLean, 1990 with prismatic complex crossed gene and the nuclear histone H3 gene supported a sister
lamellar, regular foliated and simple prismatic layers, relationship with Lepetidae (Nakano & Sasaki, 2011).
Neolepetopsis McLean, 1990 carrying complex crossed Furthermore, it has been suggested on the grounds
lamellar and prismatic complex crossed lamellar of morphology that Neolepetopsis and Paralepetopsis
layers, and Paralapetopsis McLean, 1990 with a might be too similar to be separated as two genera
prismatic complex crossed lamellar structure (Kiel, (Warén & Bouchet, 2001; Warén et al., 2006). However,
2004). In addition to lacking a ctenidium, the family this question has not been addressed by molecular
is anatomically striking in having a strongly torted phylogeny so far, owing to a complete lack of genetic
oesophagus to the posterior and lacking the osphradium information from Neolepetopsis species. All phylogenetic
(Fretter, 1990). Both protoconch morphology and large analyses so far have included only Eulepetopsis and
eggs rich in yolk suggest that neolepetopsid limpets Paralepetopsis, therefore leaving the internal structure
are lecithotrophic with a planktic dispersal stage and even monophyly of the family unsettled.
(Gustafson & Lutz, 1994; Sasaki et al., 2010). Although all neolepetopsid limpets described so far
A small family with 13 described species, the have originated from either the Pacific or Atlantic
internal structure of Neolepetopsidae has been Oceans, their presence in the Indian Ocean has been
stable, with three genera (Eulepetopsis, Neolepetopsis known for two decades. Since the discovery of the
and Paralepetopsis) since the original description. first Indian Ocean hydrothermal vent in 2000, the
Neolepetopsis is characterized by clathrate sculpture Kairei field on the Central Indian Ridge, a species of
and transparent zones on the shell interior. Currently, Eulepetopsis has been listed on the fauna list (Van
it contains five species, all from the eastern Pacific Dover et al., 2001). Later, Eulepetopsis has been found
Ocean, with four described from hydrothermal vents on a number of vent fields on the Central Indian Ridge
and one from a whale fall (McLean, 1990, 2008). (Nakamura et al., 2012; Watanabe & Beedessee, 2015;
Notably, the vent species are found mostly on inactive Kim et al., 2020) and the Southwest Indian Ridge
sulfides where no other vent-endemic life are apparent (Zhou et al., 2018; Sun et al., 2020). Nevertheless, it
(McLean, 1990; Van Dover, 2019). One vent species, has not been characterized morphologically, and it has
Neolepetopsis gordensis McLean, 1990, is now also been unclear whether the records on different ridges
known from seeps off Peru, although this potentially refer to the same species.
represents an undescribed species (Warén & Bouchet, The recent discovery of hydrothermal vent fields
2001; McLean, 2008). on the Southwest Indian Ridge (Copley et al., 2016;
Eulepetopsis has remained monotypic since Zhou et al., 2018; Sun et al., 2020) and the Carlsberg
description, with Eulepetopsis vitrea McLean, 1990 Ridge (Tao et al., 2013; Wang et al., 2017; Popoola
being the only species known from hot vents of the et al., 2019; Wang et al., 2021) have resulted in
East Pacific Rise. This genus is highly characteristic, many species new to science (e.g. Zhou et al., 2019).
with an almost completely transparent shell, the outer The latest explorations of these vents yielded two
layer of which is composed of crystal-clear lathic calcite previously unreported neolepetopsids, both clearly
(McLean, 1990). assignable to Neolepetopsis, in addition to more
Paralepetopsis is the most diverse genus in terms material of Eulepetopsis from the Carlsberg Ridge.
of both species richness and habitat types, with Here, we describe and characterize these Indian Ocean
seven described species from hydrothermal vents, neolepetopsid limpets, also using materials collected
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296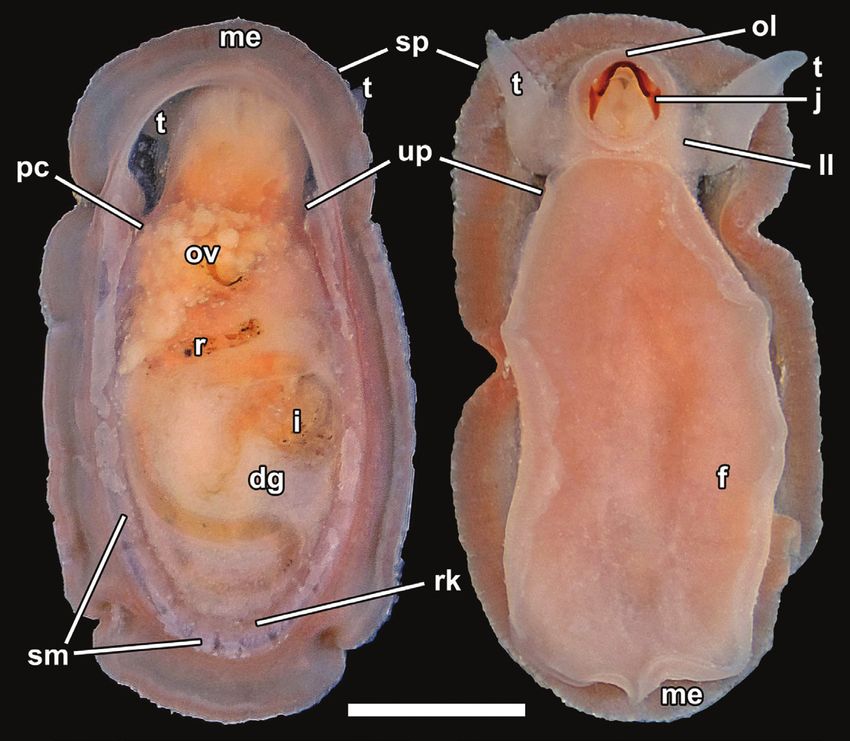
278 C. CHEN ET AL.
from the Central Indian Ridge vents. Furthermore, On the Southwest Indian Ridge, specimens were
we test the monophyly of Neolepetopsidae and seek sampled on an apparently inactive sulfide deposit
to resolve its internal genus-level structure using peripheral to the Longqi field (Copley et al., 2016)
molecular phylogenetic reconstruction that includes using a video-guided grab sampler on board R/V
the first genetic data available from Neolepetopsis. Xiangyanghong 10 cruise DY49; limpets were also
sampled from the Tiancheng field (Zhou et al., 2018;
Sun et al., 2020) on the same ridge using a seven-
function manipulator on the human-occupied vehicle
MATERIAL AND METHODS
(HOV) Jiaolong during R/V Xiangyanghong 9 cruise
Material DY35. On the Carlsberg Ridge, limpets were taken
Neolepetopsid limpets were collected from deep-sea using the same manipulator on the HOV Jiaolong
from active chimneys in the Wocan field (Wang et al.,
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
hydrothermal vents across three mid-ocean ridges
in the Indian Ocean (for a map, see Fig. 1). On the 2017) and inactive sulfide chimneys in the Daxi field
Central Indian Ridge, limpets were collected from (Wang et al., 2021) on board R/V Xiangyanghong 9
Kairei and Edmond vent fields (Van Dover et al., cruise DY38. Specimens were preserved in either
2001) using a suction sampler mounted on the deep- 10% buffered formalin or 95–99% ethanol as soon as
submergence vehicle (DSV) Shinkai 6500 on board the samples were recovered from the deep seafloor,
research vessel (R/V) Yokosuka cruise YK16-E02. after sorting of the biological material.
Figure 1. Map showing the locations of the deep-sea hydrothermal vents sampled for the present study. A, overview. B,
close-up of the Central Indian Ridge area near the Rodriguez Triple Junction. Key: 1, Wocan field, Carlsberg Ridge; 2,
Edmond field, Central Indian Ridge; 3, Kairei field, Central Indian Ridge; 4, Tiancheng field, Southwest Indian Ridge;
5, Longqi field, Southwest Indian Ridge. The maps were generated using the General Bathymetric Chart of the Oceans
(GEBCO; https://www.gebco.net/) grid display software and GEBCO 1 arc-min interval grid using the Mercator projection.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296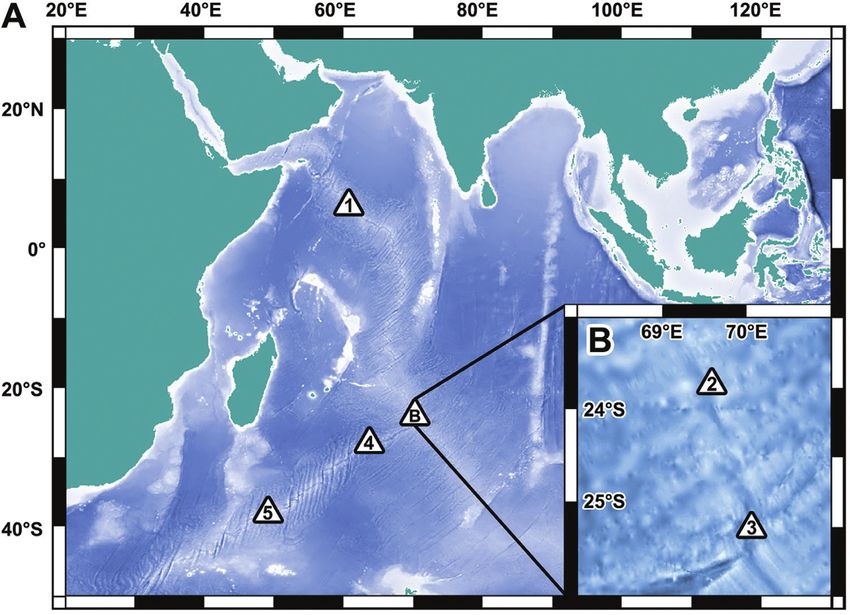
INDIAN OCEAN VENT LIMPETS 279
Morphology Phylogenetic reconstruction was undertaken by
A stereo dissecting microscope (Carl Zeiss SteREO Bayesian inference using the software package
Discovery V.12) was used to assist the observation MrBayes v.3.2.6 (Ronquist et al., 2012), using a 472 bp
and dissection of the limpet specimens collected alignment of the COI gene aligned using Geneious
from Indian Ocean vents, and photographs were R11. In addition to the newly generated sequences, we
taken using a Nikon D5000 digital single lens downloaded a selection of available COI sequences of
reflex camera mounted on the same microscope. For true limpets from GenBank with lengths > 472 bp (the
animals dehydrated in high-grade ethanol, the soft length of the shortest sequence newly generated herein).
parts were rehydrated in a gradated ethanol–MilliQ We made sure to select representatives of all currently
water series and left to sit in MilliQ water overnight accepted families, including Acmaeidae, Eoacmaeidae,
before observation was undertaken. Fine forceps and Lepetidae, Lottiidae, Nacellidae, Neolepetopsidae,
Patellidae and Pectinodontidae (Nakano & Ozawa,
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
tungsten needles were used to extract the radula
sac from the animals. Shell lengths (SL) and shell 2007; Nakano & Sasaki, 2011; Aktipis & Giribet, 2010;
widths (SW) were measured using digital callipers Goffredi et al., 2017). Two fissurellid vetigastropod
to the nearest one decimal point. sequences were selected as the outgroup (following
Nakano & Ozawa, 2007). P artition F inder v.2.1.1
(Lanfear et al., 2017) was used to select the most
Scanning electron microscopy suitable substitution model for each codon position,
Dissected radula sacs were placed in diluted which chose the GTR+I+G model for the first and
commercial bleach (1:10 dilution) under a microscope second codons and HKY+G for the third codon position.
to dissolve the tissue, while constantly being monitored Following suggestions from a previous comprehensive
to prevent disintegration, which neolepetopsid radulae phylogenetic analysis of Patellogastropoda (Nakano &
are known to be prone to (McLean, 1990, 2008). When Ozawa, 2007), we removed the problematic third codon
the radula was clean, it was immediately lifted from position from downstream analyses. For phylogenetic
the diluted bleach using tungsten needles, placed inferences, Metropolis-coupled Monte Carlo Markov
into MilliQ water and given two washes. The clean chains were run for one million generations, with
radula was lifted carefully from the water, mounted topologies being sampled every 100 generations. The
on scanning electron microscopy (SEM) stubs with software Tracer v.1.6 (Rambaut et al., 2013) was used
carbon tapes and left to dry. Shells for the observation to select a suitable burn-in value, which was 3000 in
of the protoconch and surface sculpture or structure this case. Estimation of pairwise genetic distances was
were cleaned briefly in diluted bleach and then given done using the K2P distance (Kimura, 1980) in the
two washes in MilliQ water and two further washes software package MEGA7 (Kumar et al., 2016).
in 99% ethanol before mounting directly onto SEM
stubs. All stubs were observed uncoated at 15 kV
Specimen repositories
using a Hitachi TM-3000 SEM.
Type specimens and material examined are deposited
at the National Museum of Nature and Science,
Tsukuba (NSMT), Japan and the Repository of the
Genetics
Second Institute of Oceanography (RSIO), Ministry of
E x t r a c t i o n o f g e n o m i c D N A wa s c a r r i e d o u t Natural Resources, Hangzhou, China.
using the DNeasy blood and tissue kit (Qiagen,
Valencia, CA, USA) following standard protocols.
The mitochondrial COI gene was amplified using
RESULTS
the universal primer pair HCO2198–LCO1490
(Folmer et al., 1994) in the metazoan barcoding SYSTEMATICS
region, following procedures detailed by Zhou et al.
(2018). The cycler kit (OMEGABio-tec) was used to Class Gastropoda Cuvier, 1795
purify the amplicons following the manufacturer’s Subclass Patellogastropoda Lindberg, 1986
standard protocols, and the product was sent to the Family Neolepetopsidae McLean, 1990
sequencing service at BioSune (Shanghai, China).
G eneious R11 (https://www.geneious.com) was Genus Neolepetopsis McLean, 1990
used to assemble the consensus sequence for each Type species: Neolepetopsis gordensis McLean, 1990
individual, which was then used in downstream (by original designation).
analyses. Newly generated sequences in the present
study are deposited in NCBI GenBank under the Diagnosis: Small-sized (< 10 mm) neolepetopsid
accession numbers MW543297–MW543301. limpets with coarse, typically clathrate shell sculpture.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296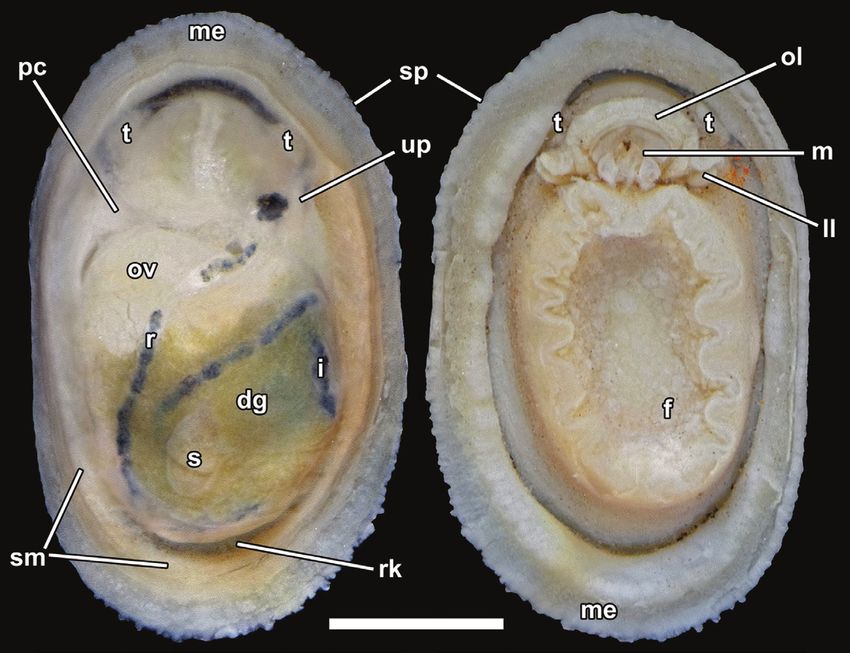
280 C. CHEN ET AL.
Shell interior with transparent areas. Radula with internal surface. Shell length ≤ 4.8 mm, shell width
sturdy central rachidian; five or seven cusped teeth, ≤ 3.3 mm (4.1 and 3.1 mm for holotype, respectively).
usually well mineralized for Neolepetopsidae (Warén Small area corresponding to apex thickest, slightly
& Bouchet, 2001; McLean, 2008). Shell profile typically opaque, thinning out towards periphery. Elliptic to
low, flat limpet-form, but may be constricted by the oval in shape, with posterior end slightly narrower
substrate type and range from flat to tall in some than anterior end; shell length-to-width ratio between
species (McLean, 1990). 1.3:1 and 1.5:1. Always low in profile, with nearly flat
shell edge. Apex at midline, nearly central or slightly
anterior. Sculpture predominantly concentric (Figs 2,
Neolepetopsis prismatica sp. nov. 3C), with strong concentric ribs at somewhat variable
(Figs 2A–D, 3, 4) interspaces between ~50 and 150 μm, even within each
individual. Radial sculpture almost lacking, with only
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
ZooBank registration: urn:lsid:zoobank.
org:act:B0DF7474-BE16-461E-B07E-632D276D0E66 weak radial striations. No nodes formed at intersection
between two directions of sculptures. Protoconch (Fig.
Diagnosis: Medium-sized Neolepetopsis (SL ≤ 4.8 mm) 3A, B) symmetrical, typical of patellogastropods,
with much stronger concentric sculpture compared 220 μm in length. Finely pitted sculpture present on
with only diminutive, nearly absent radial sculptures protoconch, especially obvious at distal end. Thin,
and lacking nodes at the intersecting points. Spacing greenish layer of periostracum present over teleoconch.
of concentric sculptures variable between ~50 and Muscle scars not clearly discernible.
150 μm within the same individual. Shell oval; posterior Radula (Fig. 3D, E) with seven mineralized teeth
end narrower than anterior; always low profile (rachidian, two pairs of laterals, pluricuspid) on
and flat limpet-form, with apex on the midline and each row and four non-mineralized marginal teeth,
nearly central or very slightly anterior. Symmetrical width ~25 μm. Cusps of mineralized teeth gradually
protoconch 220 μm in length. Greenish periostracum descending in horizontal position from rachidian
present. Interior of the semi-transparent shell carrying outwards. Rachidian sturdy; base laterally expanded
strong prismatic structural colour. Radula with short with narrow, short shaft ending in a single tapering,
shafts and heavy, well-mineralized cusps. overhanging cusp. Inner lateral also with overhanging,
but narrower, cusp stemming from narrow shaft
Type locality: Inactive sulfide deposits, Daxi possessing strong indentation to accommodate
hydrothermal vent field, Carlsberg Ridge. expanding base of rachidian. Outer pair of laterals
similar to inner pair but sturdier, with much thicker
shaft and with heavier, triangular cusp. Pluricuspid
Type material: Holotype (RSIO 38210; Fig. 2A) in tooth with broad cusp bearing three tapering tips, the
95% ethanol, SL 4.1 mm, SW 3.1 mm, Daxi vent field, two outer ones being much larger. Shaft of pluricuspid
Carlsberg Ridge, 60°10.8′E, 6°48.0′N, 3450 m deep with broad basal–lateral indentation, extension of
(station ‘DV128’), R/V Xiangyanghong 9 cruise DY38, by which connects with inner marginal tooth. Inner
a seven-function manipulator of HOV Jiaolong, dive 128, marginal with broad, overhanging edge, on strongly
11 March 2017. Paratype 1 (NSMT-Mo 79214; Figs 2B, indented, broad shaft. Outer marginal separated from
3), 95% ethanol, SL 4.2 mm, SW 2.9 mm, rehydrated inner marginal, much reduced, with narrow, tapered,
once for photography of soft parts and then stored in overhanging edge.
70% ethanol. Paratype 2 (RSIO 38211; Fig. 2C), 95% The soft parts are shown in Figure 4. Cephalic
ethanol, SL 4.0 mm, SW 2.8 mm. Paratype 3 (NSMT-Mo tentacles simple, tapering; left and right tentacles of
79215; Fig. 2D), 95% ethanol, SL 3.5 mm, SW 2.4 mm. equal size. External evidence for eyes absent. Mouth
Paratype 4 (NSMT-Mo 79216), 95% ethanol, SL 4.1 mm, with thick outer lip; labial lobe well developed. Jaw
SW 2.9 mm. Paratype 5 (NSMT-Mo 79217), 95% ethanol, present. Foot oval, with unciliated rim. Large sole
SL 3.9 mm, SW 2.8 mm. Paratype 6 (RSIO 38212), 95% demarcated from rim by deep groove. Epipodium
ethanol, SL 4.8 mm, SW 3.3 mm. Paratype 7 (NSMT-Mo lacking. Shell muscle U-shaped, situated along
79218), 95% ethanol, five small specimens. Paratype 8 posterior two-thirds of body, separated into several
(RSIO 38213), 95% ethanol, five small specimens. oval-shaped bundles decreasing in size from anterior
Paratype 9 (RSIO 38214), 95% ethanol, shell only, SL towards posterior end. Mantle edge with numerous
4.8 mm, SW 3.1 mm. All paratypes are from the same short, presumably sensory papillae; fully contracted
lot as the holotype. in specimens examined. Mantle cavity approximately
one-third of body length. Pericardium situated
Description: Shell (Fig. 2A–D) thin, mostly transparent, on left side of mantle roof, heart monotocardian,
carrying strong prismatic structural coloration on with anterior auricle and posterior ventricle (by
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296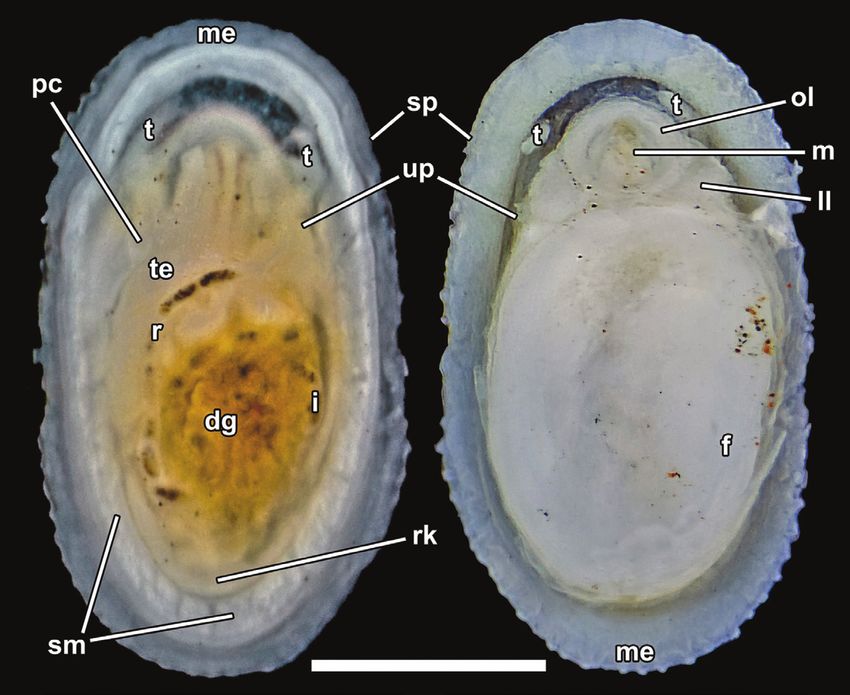
INDIAN OCEAN VENT LIMPETS 281
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
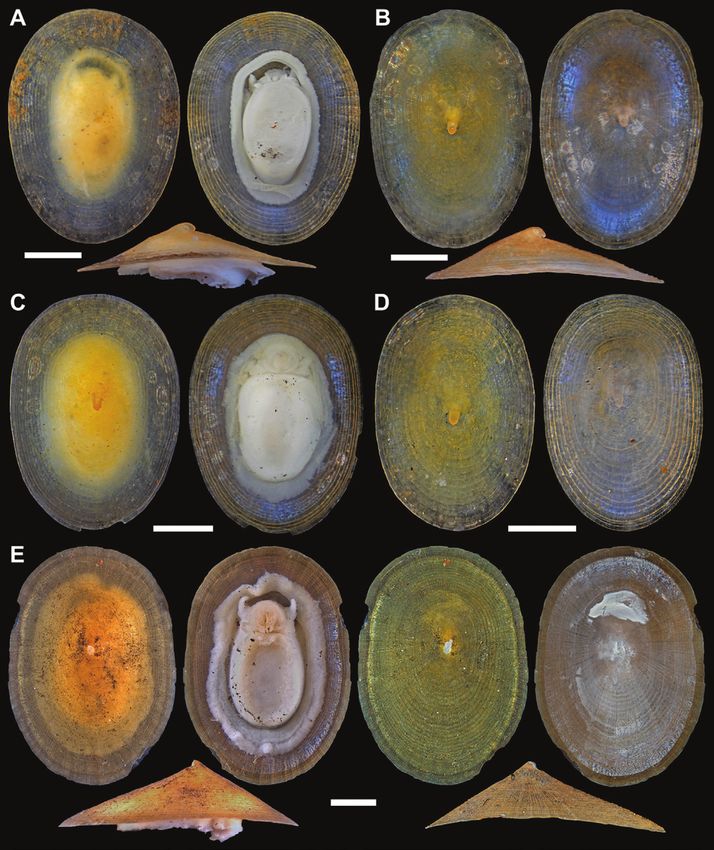
Figure 2. Neolepetopsis species. A–D, Neolepetopsis prismatica sp. nov., type specimens from the Wocan field, Carlsberg
Ridge. A, holotype (RSIO 38210), shell length (SL) 4.1 mm. B, paratype 1 (NSMT-Mo 79214), SL 4.2 mm. C, paratype 2 (RSIO
38211), SL 4.0 mm. D, paratype 3 (NSMT-Mo 79215), SL 3.5 mm. E, Neolepetopsis ardua sp. nov. from the Longqi field,
Southwest Indian Ridge, holotype (RSIO 49001), with and without the soft parts, SL 5.1 mm. Scale bars: 1 mm.
transparency). Right side of mantle roof with anus gonad located above foot at mid-ventral position,
just left of the urogenital opening situated on visible from dorsal view by transparency posterior to
sizeable papillae. Ctenidium lacking. Sexes separate; pericardium. Two kidneys present; left kidney minute
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296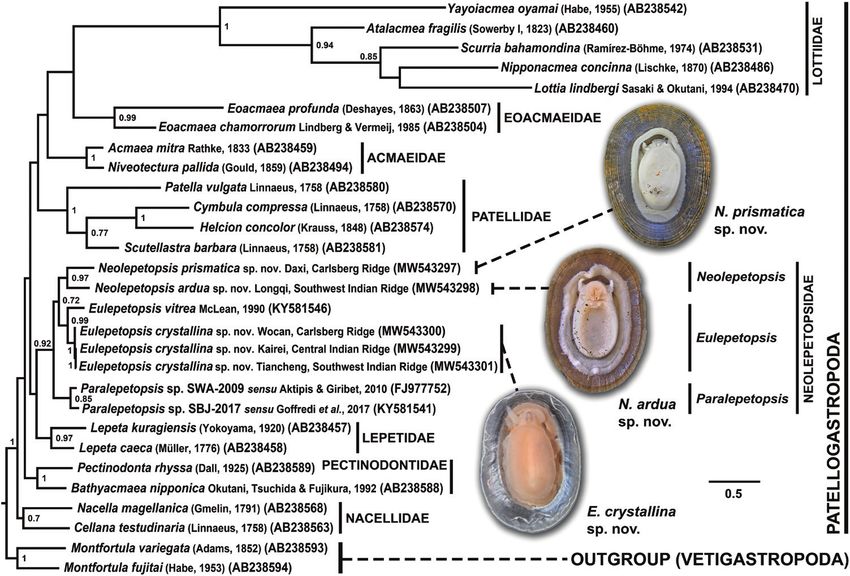
282 C. CHEN ET AL.
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 3. Neolepetopsis prismatica sp. nov., scanning electron micrographs. A, B, protoconch. Arrowheads indicate the
limits of the protoconch. A, side view. B, dorsal view. C, details of the shell sculpture. D, E, radula. Scale bars: 100 μm in A,
B; 250 μm in C; 5 μm in D, E.
compared with sizeable right kidney at far posterior. particularly the inner surface, owing to structural
Intestine and stomach embedded in large digestive colour.
gland comprising numerous branching tube-like
structures. After two loops, intestine transitions to Distribution: Currently known only from inactive
rectum, running posterior–anteriorly before curving sulfide chimneys of the Daxi hydrothermal vent field
to right, with anus exiting at right side of mantle on the Carlsberg Ridge, Indian Ocean. The Daxi field
roof. Rectum packed with dark oval faecal pellets. has a central focused venting area surrounded by
Operculum lacking. a region characterized by numerous extinct sulfide
chimneys (Wang et al., 2021), on which the present
Etymology: From the Latin adjective prismaticum new species was collected.
meaning ‘prismatic’, originally derived from Ancient
Greek πρίσμα, prísma, something sawn. It is named Remarks: Five other described species are currently
in reference to the brilliant iridescence of its shell, known in Neolepetopsis, all from the eastern Pacific
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 283
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 4. Neolepetopsis prismatica sp. nov., external anatomy (paratype 1, NSMT-Mo 79214). Left, dorsal view after
removal of the shell; right, ventral view. Abbreviations: a, anus; dg, digestive gland; f, foot; i, intestine; ll, labial lappet; m,
mouth; me, mantle edge; ol, outer lip of the mouth; pc, pericardium; r, rectum; rk, right kidney; sm, shell muscles; sp, sensory
papillae; t, cephalic tentacle; te, testis; up, urinogenital papilla. Scale bar: 1 mm.
Ocean. The shell sculpture of N. prismatica differs but unlike the longer shafts of N. gordensis. The
from these species in lacking clear nodes where radula of Neolepetopsis nicolasensis McLean, 2008
the concentric ribs meet the radial ones, owing from a whale fall off California is unique in having
to radial sculpture being extremely weak. The only five cusped teeth instead of seven (McLean,
protoconch of N. prismatica is much smaller than 2008) and therefore cannot be confused with the
that of Neolepetopsis densata McLean, 1990 from present new species. The flat shell with green
East Pacific Rise vents (McLean, 1990), the only periostracum and stronger concentric sculpture is
other Neolepetopsis species with a known protoconch also seen in Neolepetopsis verruca McLean, 1990 and
(220 vs. 400 μm). The radula of N. prismatica is also Neolepetopsis occulta McLean, 1990 from vents on
most similar to N. densata, with shorter shafts and the East Pacific Rise (McLean, 1990), but the radial
more prominent cusps than N. gordensis from vents striation on N. prismatica is far weaker than in either
on the East Pacific Rise (McLean, 1990) and also of these two species, both also exhibiting nodes where
recorded at hydrocarbon seeps off Peru (although the two sculpture types meet. For differences from
whether this record truly represents N. gordensis Neolepetopsis ardua, see below under that species.
remains uncertain; Warén & Bouchet, 2001). The
radula of other Neolepetopsis species (where known)
Neolepetopsis ardua sp. nov.
has either two (N. gordensis) or four (N. densata)
cusps on the pluricuspid (McLean, 1990), whereas (Figs 2E, 5, 6)
that of N. prismatica has three. The short shafts of ZooBank registration: urn:lsid:zoobank.
the N. prismatica radula are similar to N. densata org:act:358436F9-1BAF-452D-A97A-935F1C36BEAC
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296284 C. CHEN ET AL.
Figure 5. Neolepetopsis ardua sp. nov., scanning electron micrographs. A, apex. B, details of the shell sculpture. C, D, Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
radula. Scale bars: 100 μm in A; 200 μm in B; 10 μm in C, D.
Diagnosis: Medium-sized Neolepetopsis (SL ≤ 5.1 mm) Type locality: Apparently inactive sulfide deposits
with stronger concentric sculpture compared with off the centre of venting activity at the Longqi
radial sculptures, intersecting points of which are hydrothermal vent field, Southwest Indian Ridge.
weakly drawn out to form nodes. Concentric sculptures
between 50 and 150 μm apart. Shell profile low, Type material: Holotype (RSIO 49001; Fig. 2E), the
flat limpet-form, with apex on the midline and only specimen collected with both intact shell and
slightly anterior to the centre. Greenish periostracum animal, in 95% ethanol, SL 5.1 mm, SW 3.7 mm,
present. Longqi vent field, Southwest Indian Ridge, 49°38.88′E,
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 285
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 6. Neolepetopsis ardua sp. nov., external anatomy (paratype 1, NSMT-Mo 79219). Left, dorsal view after removal of
the shell; right, ventral view. Abbreviations: a, anus; dg, digestive gland; f, foot; i, intestine; ll, labial lappet; m, mouth; me,
mantle edge; ol, outer lip of the mouth; ov, ovary; pc, pericardium; r, rectum; rk, right kidney; s, stomach; sm, shell muscles;
sp, sensory papillae; t, cephalic tentacle; up, urinogenital papilla. Scale bar: 1 mm.
37°46.80′S, 2778 m deep (station ‘49I-S011-TVG04’), by strong, closely spaced (interspacing ranging from
R/V Xiangyanghong 10, by video-guided grab sampler, 50 to 150 μm on holotype) concentric ribs crossing
1 June 2018. Paratype 1 (NSMT-Mo 79219; Fig. 6), soft weaker, frequent radial ribs. Weak, almost indistinct
parts only, in 95% ethanol. Paratype 2 (RSIO 49010), nodes drawn out at points where concentric and radial
soft parts only, in 95% ethanol. Paratype 3 (NSMT-Mo ribs cross. Muscle scar clearly discernible. Protoconch
79220), soft parts only, in 95% ethanol. Paratype 4 unknown; holotype with seal along opening of
(RSIO 49011–49015), a lot of five individuals with protoconch (Fig. 5A). Thin, greenish periostracal layer
soft parts only, in 95% ethanol. Paratype 5 (NSMT-Mo present over shell.
79221), a lot of five individuals with soft parts only, in Radula (Fig. 5C, D) with seven mineralized
95% ethanol. All paratypes are from the same lot as (rachidian, two pairs of laterals, pluricuspid) and
the holotype. four non-mineralized (two pairs of marginals) teeth
per row, ~35 μm wide. Rachidian sturdy; base of long
Description: Shell (Fig. 2E) thin, translucent, shaft slightly expanded laterally, tapering distally
thickest near apex; only known adult shell with toward one overhanging, triangular cusp with smooth
length 5.1 mm, width 3.7 mm (length-to-width ratio cutting edge. Laterals gradually descending in
1.38:1). Shell thickness decreases towards shell edge, horizontal position outward. Inner laterals with minor
with increasing transparency, without significant indentation at base to accommodate expanded base,
prismatic structural coloration. Oval in outline; also with similar-sized overhanging, triangular, smooth
anterior end slightly narrower than posterior end cusp. Outer laterals with slightly studier shafts than
(based on holotype). Shell profile low. Apex (Fig. inner laterals, carrying broader overhanging cusps.
5A) situated on midline, slightly anterior of centre. Pluricuspid teeth positioned much lower than laterals,
Shell sculpture (Fig. 5B) finely reticulate, formed with broad overhanging cusps with two sharp tips.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296286 C. CHEN ET AL.
Shafts of pluricuspids strongly basolaterally indented; and possessing two cusps on the pluricuspid tooth
sinuous shafts connecting to inner marginal. Inner (McLean, 1990), but the mineralized cusps are
marginals with broad, indented shaft; cutting edge generally heavier than those in that species, and the
broad. Outer marginals greatly reduced, with short, marginals are more reduced in N. ardua.
narrow shafts; cusps narrow. External anatomy (Fig. 6)
similar to N. prismatica described above, but intestine
packed with darker, black coloured faecal pellets.
Genus Eulepetopsis McLean, 1990
Etymology: From Latin adjective arduus, meaning
high, steep, difficult or troublesome. It is named in Type species: Eulepetopsis vitrea McLean, 1990 (by
witness of the arduous efforts, through numerous original designation).
attempts that the authors had to make in preparing
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
and mounting the minute radula of this Neolepetopsis Diagnosis: Large (≤ 17 mm SL) neolepetopsid limpets
species. with totally transparent shell, whose outer layer
is constructed from lathic calcite. Shell profile low;
Distribution: Known only from inactive chimney shell surface almost completely smooth. Radula well
surfaces off the Longqi hydrothermal vent field on the developed and sturdy, including large inner marginals;
Southwest Indian Ridge, Indian Ocean. The sulfide outer marginal reduced to vestigial.
deposits from which the present new species was
obtained were collected by a grab sampler and, as far
as we could see from the video, they appeared inactive. Eulepetopsis crystallina sp. nov.
(Figs 7–9)
Remarks: Only one specimen (the holotype) of ZooBank registration: urn:lsid:zoobank.org:act:
N. ardua was collected intact with the shell and soft 438E4525-BBA7-437B-BA48-A8E04481FE93
parts. Shells of all other specimens available for
study have apparently been lost during the sampling Neolepetopsidae gen. sp. – Hashimoto et al., 2001:
process; some had a few shell fragments still attached, 720, table 1.
indicating that they were not shell-less when alive. All Eulepetopsis – Van Dover et al., 2001: 821, table 2.
descriptions of shell characters are therefore based Eulepetopsis sp. – Watanabe & Beedessee, 2015: 207,
solely on the holotype. table 16.1; Sun et al., 2020: 8, table 1; Kim et al., 2020:
Neolepetopsis ardua is easily distinguished from supplementary table 1.
the geographically closest species, N. prismatica, by Eulepetopsis sp. ‘SWIR’ – Zhou et al., 2018: 7, table 1.
its much stronger radial sculpture (compare Figs ‘An unnamed species known from the Kairei Vent
3C, 5B) and the presence of drawn out nodes at the Field’ – Warén et al., 2006: 83.
intersection between radial and concentric ribs.
The shell shape of N. ardua is slightly wider at the Diagnosis: Typical-sized Eulepetopsis ≤ 14.0 mm SL.
posterior (based on the holotype), whereas that of Radula with sturdy pluricuspid teeth carrying a clear
N. prismatica is either wider at the anterior end or lateral projection mid-shaft, in addition to a finely
similar in width at both ends (Fig. 2). Furthermore, serrated cutting edge.
N. ardua does not possess the same level of
iridescent structural colour on the shell interior seen Type locality: On active vent chimney, Kairei vent
on N. prismatica. The shell sculpture of N. ardua is field, Central Indian Ridge, 25°19.2315′S, 70°2.4187′E,
generally similar to that of N. densata, N. verruca 2424 m deep.
and N. occulta (McLean, 1990), but differs in that
the axial ribs are much more frequent but weak Type material: Holotype (NSMT-Mo 79222; Fig. 7A)
in strength, leading to only insignificantly raised fixed in 10% buffered formalin and stored in 70%
(as opposed to strongly raised) nodes being formed ethanol, SL 8.4 mm, SW 6.0 mm, Kairei vent field,
at the intersections. Neolepetopsis densata and Central Indian Ridge, 25°19.2315′S, 70°2.4187′E,
N. occulta have fully transparent shells (McLean, 2424 m deep, collected by suction sampler, R/V
1990), whereas that of N. ardua is only translucent. Yokosuka cruise YK16-E02, DSV Shinkai 6500
The present new species is easily distinguished dive #1449, 13 February 2016. Paratype 1 (NSMT-Mo
from both N. gordensis and N. nicolasensis by the 79223; Fig. 7B), fixed in 10% buffered formalin and
concentric sculpture being stronger than the radial stored in 70% ethanol, SL 8.7 mm, SW 8.0 mm.
ones, and by the low shell profile and the much Paratype 2 (NSMT-Mo 79224), 99% ethanol, used for
thinner, delicate shell. The radula of N. ardua is DNA extraction and sequencing, SL 10.8 mm, SW
most similar to N. gordensis in having long shafts 8.0 mm. Paratype 3 (NSMT-Mo 79225; Fig. 7C), fixed
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 287
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 7. Eulepetopsis crystallina sp. nov.. A, holotype (NSMT-Mo 79222), Kairei field, Central Indian Ridge, shell length
(SL) 8.4 mm. B, paratype 1 (NSMT-Mo 79223), Kairei field, SL 8.7 mm. C, paratype 2 (NSMT-Mo 79224), Kairei field, SL
10.8 mm. D, a specimen from Edmond field, Central Indian Ridge (NSMT-Mo 79226), with thick sulfide covering partly
removed, SL 11.8 mm. E, a specimen from Tiancheng field, Southwest Indian Ridge (RSIO 35734), SL 14.0 mm. F, a specimen
from Wocan field, Carlsberg Ridge (RSIO 38215) covered by a thin layer of sulfide deposits, SL 12.9 mm. G, close-up of the
internal surface of the shell of paratype 2 (NSMT-Mo 79224) showing the attachment area of the radial pallial muscles.
Scale bars: 2 mm in A–F; 1 mm in G.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296288 C. CHEN ET AL.
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 8. Eulepetopsis crystallina sp. nov., scanning electron micrographs. A, exterior shell surface showing a close-up of
the corroded apex. B, C, close-up of the shell interior showing zigzagged crystal edges and shell pores (arrowheads in C). D,
radula. Scale bars: 1 mm in A; 200 μm in B; 50 μm in C; 20 μm in D.
in 10% buffered formalin and stored in 70% ethanol, Xiangyanghong 9 cruise DY35, HOV Jiaolong dive 87,
SL 8.3 mm, SW 5.8 mm. All paratypes are from the 23 December 2014. Three specimens (RSIO 38215; Fig.
same lot as the holotype. 7F), Wocan vent field, Carlsberg Ridge, SL 10.5–14.0
mm, SW 7.5–10.2 mm, covered in thin layer of sulfide
Materials examined: One specimen (NSMT-Mo 79226; deposits, 60°31.8′E, 6°21.6′N, 2920 m deep, collected
Fig. 7D), Edmond vent field, Central Indian Ridge, by a seven-function manipulator, R/V Xiangyanghong
99% ethanol, SL 11.8 mm, SW 8.6 mm, 23°52.6621′S, 9 cruise DY38, HOV Jiaolong dive 129, 14 March 2017.
69°35.7959′E, 3279 m deep, collected by suction sampler,
R/V Yokosuka cruise YK16-E02, DSV Shinkai 6500 Description: Shell (Fig. 7) thin, fully transparent, with
dive #1457, 26 February 2016, covered in thick sulfide thin layer of periostracum where not corroded. Shell
layer, now removed, in part, to reveal shell surface. length oblong oval; slightly narrower at anterior end
Six specimens (RSIO 35734; Fig. 7E), Tiancheng vent than posterior end, more so in larger specimens (Fig.
field, Southwest Indian Ridge, SL 11.2–14.1 mm, 7E). Maximum known shell sizes at SL 14.0 mm, SW
SW 8.2–11.0 mm, 63°55.398′E, 27°51.030′S, 2682 m 10.2 mm. Shell profile low, flat, with margin almost
deep, collected by a seven-function manipulator, R/V aligned along one plane in smaller specimens, becoming
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 289
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 9. Eulepetopsis crystallina sp. nov., external anatomy (paratype 1, NSMT-Mo 79223). Left, dorsal view after removing
the shell; right, ventral view. Abbreviations: a, anus; dg, digestive gland; f, foot; i, intestine; j, jaw projecting from the mouth;
ll, labial lappet; me, mantle edge; ol, outer lip of the mouth; pc, pericardium; r, rectum; rk, right kidney; s, stomach; sm, shell
muscles; sp, sensory papillae; t, cephalic tentacle; te, testis; up, urinogenital papilla. Scale bar: 1 mm.
more uneven in larger ones (Fig. 7). Protoconch unknown; narrow, triangular, overhanging cusp. Inner laterals
inner surface of protoconch sealed in specimens with lost with elongate, tapering triangular cusps on a solid
protoconchs. Apex situated on midline anteriorly, about shaft carrying strong indentation to accommodate
one-quarter of shell length from anterior edge. Shell lateral supports of rachidian tooth. Outer laterals
surface almost completely smooth except for concentric about twice as broad as inner laterals; shafts with
growth lines (Fig. 7C); area near apex corroded, with weak indentation to accommodate inner laterals, each
uneven lines (Fig. 8A). Inner surface of shell showing carrying one prominent lateral projection near base.
muscle scars (Fig. 7B); area near apex surrounded by a Pluricuspid robust, more than twice as wide as outer
series of pores going into interior of shell towards apex laterals, with prominent lateral, mid-shaft projection.
(Fig. 8B, C), seen as long radial streaks by transparency Overhanging cusp of pluricuspid broad, with numerous
under optical microscopy (Fig. 7G). Zigzagged crystal fine serrations decreasing in strength outward. Laterals
edges visible on inner surface of shell with electron and pluricuspid decrease in cusp positions outward
scanning microscopy (Fig. 8C). from the rachidian. Inner marginal well formed, as
Radula (Fig. 8D) with sturdy rachidian and two broad as pluricuspid, with narrow, smooth, semi-
laterals, pluricuspid tooth, two marginals on either circular, overhanging cutting edge. Second marginal
side. Cusps not well mineralized. Rachidian well vestigial, found slightly outside of inner marginal.
supported, with laterally expanded base; shaft of Soft parts are shown in Figure 9. Cephalic tentacles
moderate length, slowly tapering apically, ending in simple conical, without appendages, elongate, tapered.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296290 C. CHEN ET AL.
No external evidence for eyes. Oral disc with muscular in E. crystallina are also noticeably longer than in
outer lip, surrounded by moderately developed labial E. vitrea (Warén & Bouchet, 2001).
lobe. Well-developed, dorsally arched jaw present, often
seen projecting from mouth in preserved specimens.
Sole of foot oval, large, with unciliated rim demarcated Genetic support
by deep groove from sole. Epipodium lacking. Shell The consensus tree for Patellogastropoda from
muscle U-shaped, separated into numerous oblong phylogenetic reconstruction by Bayesian inference
muscle bundles along the posterior three-quarters of using first and second codon positions of a 472 bp
body, with length of bundles decreasing posteriorly. alignment in the barcoding region is shown in Figure
Mantle edge with numerous fine, presumably 10. Neolepetopsis prismatica was recovered as sister
sensory papillae. Mantle cavity shallow, extending to N. ardua with strong support [Bayesian posterior
to slightly shy of one-third of body length. Heart probability (BPP) = 0.97], with the clade interpreted
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
monotocardian, with auricle anterior of ventricle (seen as genus Neolepetopsis. The three sequences of
by transparency), located within pericardium on left E. crystallina included, one from each mid-ocean ridge,
mantle roof. Ctenidium lacking. Sexes separate; gonad were recovered as a fully supported clade corresponding
located ventrally along mid-body, partly visible from to the new species. This was sister to E. vitrea, with
dorsal view slightly posterior to pericardium. Left the two species forming a strongly supported clade
kidney minute; sizeable right kidney positioned at (BPP = 0.99) representing genus Eulepetopsis.
posterior of body. Urogenital papillae on right mantle Eulepetospsis was recovered as sister to Neolepetopsis
roof on right side of anus. Intestine much wider with moderate support (BPP = 0.72), with this pair in
anteriorly, looping twice before emerging at posterior turn being sister to a moderately supported (BPP = 0.85)
end of body as rectum. Rectum runs towards anterior Paralepetopsis containing two (undescribed) species.
left before turning to anterior right and finally This means that Paralepetopsis was recovered in a basal
emerges on right mantle roof. Intestine and stomach position within Neolepetopsidae, which was recovered
entirely embedded within voluminous digestive gland, as a well-supported (BPP = 0.92) clade containing the
comprising numerous tubular structures. Operculum three abovementioned genera.
lacking. At the level of families within Patellogastropoda,
all currently established patellogastropod families
Etymology: From Latin crystallinum, crystal-like, (Nakano & Ozawa, 2007; Aktipis & Giribet, 2010;
named for its highly transparent shell. Nakano & Sasaki, 2011; Goffredi et al., 2017) were
recovered as moderately to well-supported clades
Distribution: Known from a number of hydrothermal (BPP = 0.7–1.0), but all sister relationships between
vent fields across Carlsberg Ridge (Wocan field), families were not well supported. Neolepetopsidae was
Central Indian Ridge (Kairei and Edmond fields) recovered as sister to Lepetidae, but this relationship
and Southwest Indian Ridge (Tiancheng field). Given was not well supported (BPP = 0.52).
its distribution range from the examined materials, The genetic distances (K2P distances), estimated
Eulepetopsis recorded at both Solitaire (Nakamura using 472 bp of the COI gene, among neolepetopsid taxa
et al., 2012; Watanabe & Beedessee, 2015) and Onnuri with suitable data available are shown in Table 1. The
(Kim et al., 2020) fields are most likely additional genetic distance for the three specimens of E. crystallina
records of this species. included, one from each ridge system, was 0.21–0.85%.
This is much lower than the distance between these and
E. vitrea, which ranged between 9.79 and 10.30%. The
Remarks: Eulepetopsis crystallina is similar genetic distance between N. prismatica and N. ardua
morphologically to E. vitrea, the only other known was estimated at 5.54%, and that between the two
species in the genus, described from vents on the East undescribed Paralepetopsis species (Aktipis & Giribet,
Pacific Rise. Both have highly transparent shells and 2010; Goffredi et al., 2017) was 12.26%. The average
similar anatomical features (Fretter, 1990), but are genetic distance between species assigned to the same
separable based on radula morphology, most notably genera was 9.59% (range 5.54–12.26%), and between
the pluricuspid teeth. In E. crystallina, the pluricuspid species of different genera it was 13.95% (12.27–15.09%).
has a prominent mid-shaft projection, which is lacking
in that of E. vitrea (McLean, 1990; Warén & Bouchet,
2001). The finely serrated cutting edge is another DISCUSSION
feature not mentioned in E. vitrea; the pluricuspid
teeth as a whole are much stronger and broader at Distribution and biogeography
the base in E. crystallina than in E. vitrea (McLean, The wide distribution of E. crystallina across three mid-
1990; Warén & Bouchet, 2001). Shafts of all teeth ocean ridges in the Indian Ocean might appear striking
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 291
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Figure 10. Consensus tree of Patellogastropoda from phylogenetic reconstruction using Bayesian inference, based on
472 bp of the mitochondrial COI gene at the barcoding region. Node values indicate Bayesian posterior probabilities, shown
only for those > 0.7. GenBank accession numbers of the sequences used are indicated in parentheses after the authority and
date for each species.
Table 1. Estimates of genetic distances (K2P distances; as percentages) among neolepetopsid limpets, using 472 bp of the
mitochondrial COI gene
1 2 3 4 5 6 7 8
1 Neolepetopsis prismatica Daxi (MW543297)
2 Neolepetopsis ardua Longqi (MW543298) 5.54
3 Eulepetopsis vitrea (KY581546) 14.08 15.09
4 Eulepetopsis crystallina Wocan (MW543300) 13.80 14.57 10.30
5 Eulepetopsis crystallina Kairei (MW543299) 13.53 14.84 10.05 0.64
6 Eulepetopsis crystallina Tiancheng (MW543301) 13.80 14.57 9.79 0.85 0.21
7 Paralepetopsis sp. SWA-2009 (FJ977752) 15.06 15.06 13.94 14.00 13.74 13.48
8 Paralepetopsis sp. SBJ-2017 (KY581541) 13.74 14.27 13.74 12.79 12.53 12.27 12.26
GenBank accession numbers are indicated in parentheses. New species in bold.
compared with the apparently limited distribution of known to exhibit similar lecithotrophic development
N. prismatica and N. ardua, each known from only with planktonic stages and most probably similar
one vent field. However, even though both genera are dispersal capabilities (McLean, 1990; inferred from
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296292 C. CHEN ET AL.
protoconch morphology), this is likely to be explained the world. This adds to the existing evidence that the
by the combined effect of habitat preference and the periphery of vents and the inactive sulfides probably
associated sampling bias. host different vent-endemic taxa from the well-
Eulepetopsis crystallina, much like E. vitrea on the surveyed central parts (Van Dover, 2019), and the
East Pacific Rise (Warén & Bouchet, 2001), occurs close recent discovery of two Amphisamytha Hessle, 1917
to active vent orifices and associated with other typical polychaete species only found on inactive sulfides off
vent fauna, such as Bathymodiolus Kenk & Wilson, Longqi (Zhou et al., 2019) indicates that there are
1985 mussels (Van Dover et al., 2001; Watanabe & more species waiting to be discovered. As the deep-
Beedessee, 2015). Conversely, N. prismatica and sea mining industry mulls targeting areas of inactive
N. ardua were collected on inactive sulfide deposits sulfide areas owing to the potential environmental
some distance away from the active vent orifices. damage from mining active vents (Van Dover, 2019),
This is in agreement with the other eastern Pacific it is imperative that we shift some of the sampling
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Neolepetopsis species for which the habitat is known, effort away from the dramatic vent orifices and aim
with N. densata and N. occulta being collected from to understand biology at inactive vents before mining
inactive sulfides lacking other fauna (Van Dover, 2019) commences, especially in the Indian Ocean (Sigwart
and N. verruca at one nautical mile away from the vent et al., 2019).
orifice where no other vent fauna could be collected; Another factor contributing to the realized
the habitat of N. gordensis at vents is unknown distribution range of the three Indian Ocean
(McLean, 1990). Taken together, these findings neolepetopsids, especially clear for E. crystallina,
suggest that Eulepetopsis is a genus highly adapted to is the projected dispersal barrier on the Southwest
hydrothermal vent habitat and capable of inhabiting Indian Ridge between Longqi and Tiancheng (Sun
areas near black smokers, but Neolepetopsis is et al., 2020). As discussed above, Eulepetopsis occur
restricted to peripheral areas and unable to invade the at or near the active centre of vents and, given the
most active parts of vent fields. Neolepetopsis, but not sampling efforts at Longqi, it is probably truly absent
Eulepetopsis, being known also from seeps and whale there (Zhou et al., 2018). Being widely distributed
falls (Warén & Bouchet, 2001; McLean, 2008) supports along the Carlsberg Ridge and Central Indian Ridge,
this hypothesis. extending to Tiancheng, but absent at Longqi is a
There is a stark sampling bias at hydrothermal vent pattern shared with other key vent endemic animals
sites, where the centre of venting activity is the ‘centre such as the hairy snail Alviniconcha marisindica
of the show’ and receives numerous and repeated Okutani in Johnson et al., 2014 and the vent crab
sampling efforts, whereas the vent periphery remains Austinograea rodriguezensis Tsuchida & Hashimoto,
little studied (Chen et al., 2017), and inactive sulfides 2002 (Sun et al., 2020). On the contrary, Longqi shares
are virtually unsampled in terms of biology (Van many taxa at the genus level, such as the yeti crabs
Dover, 2019). Previous dense sampling for biology on (Kiwa Macpherson, Jones & Segonzac, 2006) and
the Southwest Indian Ridge during multiple research the giant peltospirid snail Gigantopelta Chen, Linse,
cruises using submersibles at the Longqi vent field did Roterman, Copley & Rogers, 2015, with Antarctic
not yield specimens of N. ardua (Copley et al., 2016; Zhou vents on the East Scotia Ridge that are missing in
et al., 2018), because they concentrated on areas close Tiancheng and eastwards (Copley et al., 2016).
to the venting activity. The specimens in the present A number of major transform faults are present
study were yielded from a video-guided grab sampler between Tiancheng and Longqi on the Southwest
that took sulfide samples well into the peripheral areas, Indian Ridge, which is likely to act as the dispersal
a sampling method and target area mostly used by barrier for these species (Sun et al., 2020). The
geologists to collect sulfides for mineralogy studies (Van dispersal of E. crystallina to Longqi is likely also
Dover, 2019). This effect is enhanced by biologists often to be restricted by these transformation faults.
being absent on geology cruises, leading to any potential Likewise, N. ardua, with similar dispersal ability
specimens being lost to science. This sampling bias can (Sasaki et al., 2010), is unlikely to be found in
explain why Eulepetopsis appears to be much more Tiancheng. Nevertheless, E. crystallina is still one of
widely distributed. Neolepetopsis is likely to be present few species known to occur across three mid-ocean
in other Indian Ocean vents, such as those on the Central ridges in the Indian Ocean (Watanabe et al., 2018;
Indian Ridge, but has simply not been sampled yet. Sun et al., 2020) and would be a prime candidate for
The two new Neolepetopsis species described herein population genetic studies in the future. Inferring
represent the first discovery of the genus anywhere from the range of E. crystallina, the true distribution
outside eastern Pacific (McLean, 2008), with is the most range of N. prismatica is likely to be wider than the
well-studied area in terms of vents (Thaler & Amon, Daxi field and possibly extends as far as Tiancheng,
2019). It seems likely that many more Neolepetopsis but this needs more off-vent sampling in the future
species await discovery on inactive sulfides around to confirm that this is the case.
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296INDIAN OCEAN VENT LIMPETS 293
Although Kim et al. (2020) recorded Paralepetopsis bathymodioline mussels (Thubaut et al., 2013) and
ferrugivora Warén & Bouchet, 2001 from the Onnuri cocculinid limpets (Chen et al., 2020).
vent field on the Central Indian Ridge without providing In our phylogeny Neolepetopsidae was recovered
any morphological or molecular data, P. ferrugivora was as sister to Lepetidae, a relationship previously
described from the Lucky Strike vent field on the Mid- recovered by a study combining COI and H3
Atlantic Ridge and, owing to the large geographical genes (Nakano & Sasaki, 2011). Conversely, an
distance, its presence in the Indian Ocean is extremely older study using a partial 18S gene indicated a
unlikely. We consider that this is most likely to be a closer relationship between Neolepetopsidae and
misidentification of sulfide-covered E. crystallina (see Acmaeidae, rather than Lepetidae (Harasewych
Fig. 7D) or another neolepetopsid species. & McArthur, 2000). Although this lends support to
the Neolepetopsidae–Lepetidae sister-relationship
hypothesis, this crucial node was not well supported
Downloaded from https://academic.oup.com/zoolinnean/article/194/1/276/6408716 by guest on 29 June 2022
Molecular phylogeny in our phylogeny (BP = 0.52); hence, we consider
The molecular phylogeny presented herein is the first to that the sister group to Neolepetopsidae remains
include all three genera of Neolepetopsidae, and the fact uncertain. Indeed, the lack of any well-supported
that all three clustered together to form a monophyletic relationships between families in our phylogeny
group with strong support (BP = 0.92) supports the shows the limit of the COI gene, and we refrain
monophyly of Neolepetopsidae. This reconstruction also from discussing family-level relationships within
substantiates the species-level status of the three new Patellogastropoda here. Family-level systematics
species proposed herein and confirms that there is a within Patellogastropoda remain in a state of flux,
Eulepetopsis species distributed along three mid-ocean attributable, in part, to long-branch attraction. To
ridges in the Indian Ocean.This is corroborated further by resolve this reliably would require a well-sampled
the 5.54% genetic distance (K2P distance) between mitogenome-level or potentially genome-level
the two Indian Ocean Neolepetopsis species described phylogeny in the future (Uribe et al., 2019).
herein and 9.79–10.30% distance between E. crystallina
and E. vitrea. A small barcoding gap between species
assigned to the same genus and those in different Conclusions
genera was found, with the mean values being 9.59% We studied neolepetopsid limpets discovered at deep-sea
(range 5.54–12.26%) vs. 13.95% (range 12.27–15.09%), hot vents across three mid-ocean ridges in the Indian
but their ranges almost overlap. However, we were Ocean using integrative methods, revealing three new
limited by the taxa available, and denser sampling species in two genera. The two Neolepetopsis species,
of neolepetopsid species is required to gain a better namely N. prismatica from the Daxi field, Carlsberg
understanding of the genetic distances among the Ridge and N. ardua from the Longqi field, Southwest
three genera. Indian Ridge, were recovered from off-vent settings on
Our phylogenetic reconstruction also provides a first inactive sulfides. In contrast, E. crystallina is a widely
glimpse into the internal structure of Neolepetopsidae distributed species found near the actively venting
by confirming the presence of three distinct genus-level areas of vents from the Wocan field on the Carlsberg
clades corresponding to Eulepetopsis, Neolepetopsis Ridge to the Tiancheng field on the Southwest Indian
and Paralepetopsis. Although the distinctiveness Ridge. We deduce that the apparently much wider
between Neoleletopsis and Paralepetopsis has been distribution range of E. crystallina, although the two
questioned (Warén & Bouchet, 2001), our results show genera should have similar dispersal capabilities, is
that the differences seen in the shell and radular attributable to a lack of sampling effort in the inactive
characters between Neolepetopsis and Paralepetopsis areas and to the likely dispersal barriers between
(McLean, 1990, 2008) reflect internal structure of the Longqi and Tiancheng. Our phylogenetic reconstruction
family, plausibly at the genus level. includes the first molecular data from Neolepetopsis,
The basal position of Paralepetopsis within confirming the monophyly of Neolepetopsidae and that
Neolepetopsidae and the sister relationship between Neolepetopsis is a genus distinct from Paralepetopsis.
Neolepetopsis and Eulepetopsis at a more derived The basal position of Paralepetopsis, mostly known from
position suggest that Eulepetopsis evolved from a seeps and whale falls, within Neolepetopsidae and the
less vent-specialized ancestor perhaps similar to relatively derived position of the active-vent-adapted
Paralepetopsis, a genus known mostly from cold Eulepetopsis indicates a ‘stepping-stone’ pathway for the
seeps and whale falls (McLean, 2008). This suggests evolution of its vent endemicity. The two Neolepetopsis
that Eulepetopsis adapted to the most ‘extreme’ hot species described herein represent the first records
vent habitat through a ‘stepping-stone’ evolutionary of the genus outside the eastern Pacific, suggesting a
trajectory from organic falls and seeps to vents (Smith much wider distribution for the genus than previously
et al., 1989, 2015), as has been demonstrated for thought. Future explorations of hydrothermal vents
© 2021 The Linnean Society of London, Zoological Journal of the Linnean Society, 2022, 194, 276–296You can also read

















































