Metagenomic insights into the taxonomy, function, and dysbiosis of prokaryotic communities in octocorals
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Keller-Costa et al. Microbiome (2021) 9:72
https://doi.org/10.1186/s40168-021-01031-y
RESEARCH Open Access
Metagenomic insights into the taxonomy,
function, and dysbiosis of prokaryotic
communities in octocorals
T. Keller-Costa1* , A. Lago-Lestón2*, J. P. Saraiva3, R. Toscan3, S. G. Silva1, J. Gonçalves4, C. J. Cox4, N. Kyrpides5,
U. Nunes da Rocha3 and R. Costa1,4,5
Abstract
Background: In octocorals (Cnidaria Octocorallia), the functional relationship between host health and its symbiotic
consortium has yet to be determined. Here, we employed comparative metagenomics to uncover the distinct
functional and phylogenetic features of the microbiomes of healthy Eunicella gazella, Eunicella verrucosa, and
Leptogorgia sarmentosa tissues, in contrast with the microbiomes found in seawater and sediments. We further
explored how the octocoral microbiome shifts to a pathobiome state in E. gazella.
Results: Multivariate analyses based on 16S rRNA genes, Clusters of Orthologous Groups of proteins (COGs), Protein
families (Pfams), and secondary metabolite-biosynthetic gene clusters annotated from 20 Illumina-sequenced
metagenomes each revealed separate clustering of the prokaryotic communities of healthy tissue samples of the
three octocoral species from those of necrotic E. gazella tissue and surrounding environments. While the healthy
octocoral microbiome was distinguished by so-far uncultivated Endozoicomonadaceae, Oceanospirillales, and
Alteromonadales phylotypes in all host species, a pronounced increase of Flavobacteriaceae and Alphaproteobacteria,
originating from seawater, was observed in necrotic E. gazella tissue. Increased abundances of eukaryotic-like
proteins, exonucleases, restriction endonucleases, CRISPR/Cas proteins, and genes encoding for heat-shock proteins,
inorganic ion transport, and iron storage distinguished the prokaryotic communities of healthy octocoral tissue
regardless of the host species. An increase of arginase and nitric oxide reductase genes, observed in necrotic E.
gazella tissues, suggests the existence of a mechanism for suppression of nitrite oxide production by which
octocoral pathogens may overcome the host’s immune system.
(Continued on next page)
* Correspondence: tinakellercosta@tecnico.ulisboa.pt; alago@cicese.mx
1
Instituto de Bioengenharia e Biociências (iBB), Instituto Superior Técnico
(IST), Universidade de Lisboa, Av. Rovisco Pais 1, 1049-001 Lisbon, Portugal
2
División de Biología Experimental y Aplicada (DBEA), Centro de
Investigación Científica y de Educación Superior de Ensenada (CICESE), Carr.
Ensenada-Tijuana 3918, Zona Playitas, C.P 22860 Ensenada, Baja California,
Mexico
Full list of author information is available at the end of the article
© The Author(s). 2021 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License,
which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if
changes were made. The images or other third party material in this article are included in the article's Creative Commons
licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the
data made available in this article, unless otherwise stated in a credit line to the data.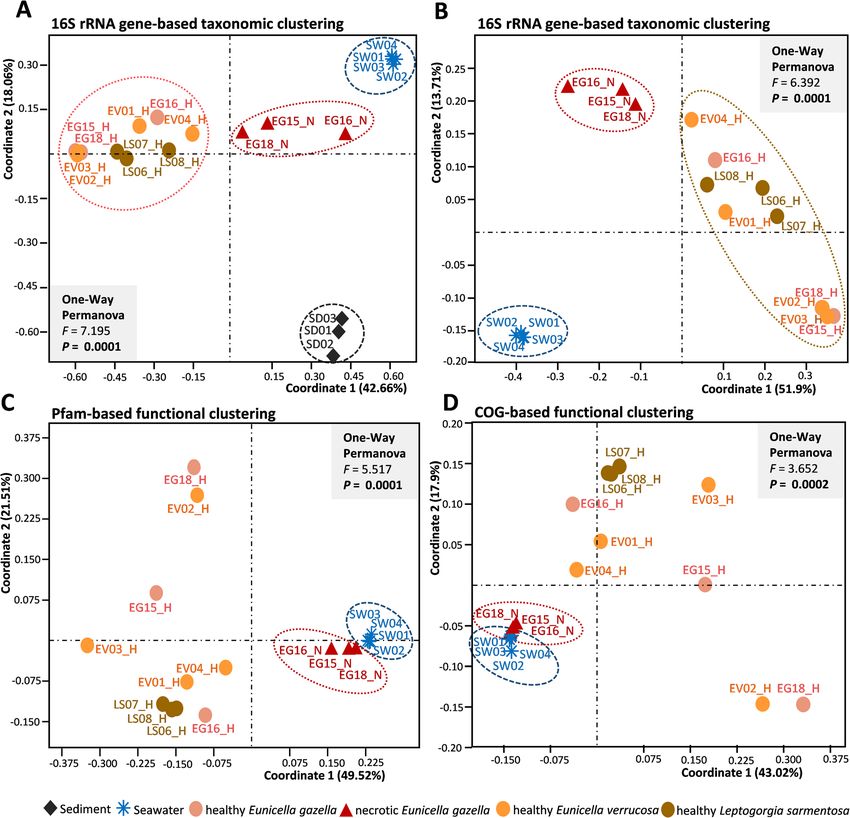
Keller-Costa et al. Microbiome (2021) 9:72 Page 2 of 21 (Continued from previous page) Conclusions: This is the first study to employ primer-less, shotgun metagenome sequencing to unveil the taxonomic, functional, and secondary metabolism features of prokaryotic communities in octocorals. Our analyses reveal that the octocoral microbiome is distinct from those of the environmental surroundings, is host genus (but not species) specific, and undergoes large, complex structural changes in the transition to the dysbiotic state. Host-symbiont recognition, abiotic-stress response, micronutrient acquisition, and an antiviral defense arsenal comprising multiple restriction endonucleases, CRISPR/Cas systems, and phage lysogenization regulators are signatures of prokaryotic communities in octocorals. We argue that these features collectively contribute to the stabilization of symbiosis in the octocoral holobiont and constitute beneficial traits that can guide future studies on coral reef conservation and microbiome therapy. Keywords: Host-microbe interactions, Symbiosis, Holobiont, Gorgonians, Eunicella, Leptogorgia, Secondary metabolism, Necrosis Background to the Oceanospirillales genus Endozoicomonas, which Communities of beneficial, commensal, and opportunis- can make up to 96% of an octocoral’s prokaryotic com- tic microbes are an integral part of most animals, and munity and are considered core symbionts [8–12, 15– together, they may form an ecological unit, called the 18]. Some Endozoicomonas phylotypes are capable of holobiont [1]. Corals are no exception [2], and coral- dimethylsulfoniopropionate (DMSP) metabolization and microbiome interactions are assumed to play a key role are thought to play a role in sulfur cycling within their in host’s resilience in a climate change scenario. How- host [17, 19]. The maintenance of a functional micro- ever, our ability to benefit from such interactions in con- biome seems essential to coral holobiont fitness [20], servation practices is severely limited due to our lack of and bacterial symbionts of corals have been implicated understanding of the microbial functions that are indis- in several important functions such as carbon acquisi- pensable for holobiont health and functioning. tion [17, 21, 22], nitrogen fixation [8, 17, 23], sulfur cyc- Octocorals (Cnidaria, Anthozoa, Octocorallia), charac- ling [8, 17, 19, 24], phosphorous supply [17], and terized by the eight-fold symmetry of their polyps, com- biosynthesis of antimicrobial compounds [25, 26]. In- prise a lineage of metazoans that diverged from deed, octocorals and their microbial associates are pro- scleractinian corals (Hexacorallia) more than 540 million lific producers of bioactive compounds with ca. 200 years ago [3]. Over 3500 octocoral species have been de- novel structures described yearly [27] and hence present scribed to date [4]. Octocorals are distributed worldwide an enormous potential for the blue economy sector [28]. [5] and are model organisms in the study of the evolution Abiotic and biotic stressors such as infectious diseases of the metazoan immune and hormonal system. In tem- and climate change have led to significant mortalities in perate marine biomes, octocorals can make up to 95% of Mediterranean and North-East Atlantic octocoral popula- the total biomass, dictating carbon flux dynamics, increas- tions during the past two decades [29–33]. Such mortality ing benthic biodiversity, and functioning as true ecosystem events can alter critical ecosystem processes and result in engineers [6, 7]. biodiversity loss in the benthos of temperate zones [30]. As with other scleractinian corals, octocorals live in Despite their devastating impacts, the community shifts symbiosis with complex microbial communities. How- underpinning microbiome dysbiosis, tissue necrosis, and ever, current knowledge of the structure and function of ultimately mortality in octocorals remain largely un- the octocoral-associated microbiome is still relatively known. A 16S rRNA gene-based pyrosequencing study scant. Many temperate octocoral species are azoox- [34] and a few, bacterial cultivation-based studies have anthellate (in the Mediterranean Sea, more than 95%), pointed towards infections with Vibrio pathogens as meaning they lack the photosynthetic Symbiodinia- disease-causing agents [31, 32]; however, holistic experi- ceae symbionts [8]. Instead, microbial associations in mental and/or omics-based investigations are still lacking. these octocorals are presumed to be dominated by che- Such studies are necessary to reveal the functional features motrophic prokaryotes [8]. Most octocoral microbiome of the microbiomes of healthy octocorals and the environ- studies have been restricted to 16S rRNA gene-based mental and biotic triggers of disease, and to develop ef- taxonomic profiling, yet they have revealed that octo- fective mitigation strategies in the long term. coral bacterial communities are diverse, distinct from In this study, we aimed to, firstly, determine whether seawater, and often host species specific [8–14]. Their the functional and taxonomic structure of prokaryotic bacterial assemblages are commonly dominated by Pro- communities inhabiting temperate octocorals are (i) host teobacteria, particularly Gammaproteobacteria belonging species specific and (ii) distinct from those of the
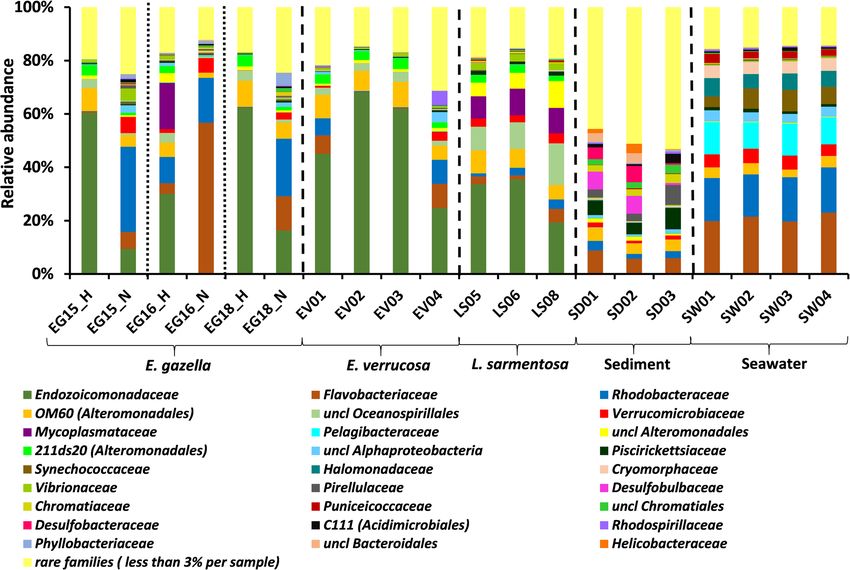
Keller-Costa et al. Microbiome (2021) 9:72 Page 3 of 21 environmental vicinities and, secondly, to reveal func- phylogenetic relationships of secondary metabolite bio- tions that could serve as indicators of octocoral holo- synthetic gene clusters (BGCs) detected in the samples, biont health. To this end, we employed shotgun thereby illuminating shifts in secondary metabolism and metagenome sequencing of the microbiomes of healthy the potential for natural product biosynthesis across tissue samples from three octocoral species, Eunicella octocoral and seawater microbiomes. gazella (N = 3), E. verrucosa (N = 4), and Leptogorgia sarmentosa (N = 3), in addition to necrotic E. gazella tis- Results sue (N=3) (Additional file 1: Figure S1), and their sur- Overview of the microbial metagenome dataset rounding seawater (N = 4) and sediment (N = 3). To About 20 million paired end reads of 101 nucleotides in reveal shifts in prokaryotic community structures be- length were generated per sample. In total, 46.12 Gb of tween the different octocoral species, their health states, sequencing information was obtained across the 20 and surrounding environments, we firstly performed metagenome samples analyzed in this study (Add- taxonomic 16S rRNA gene-based profiling of all 20 itional file 2: Table S1). A total of 291,597,021 high- metagenomes. Subsequently, we used Clusters of Ortho- quality unassembled reads (averaging 14,579,851 reads logous Groups of proteins (COGs) and Protein families per sample) were scanned by the MGnify v.2.0 metage- (Pfam)-based annotation to reveal differentially abun- nomics pipeline from the European Bioinformatics Insti- dant functions across the prokaryotic communities in tute (EMBL-EBI) [35] for the detection of both protein- octocorals (health and necrotic tissues) and seawater. coding sequences (CDS) and rRNA genes (Add- Because octocorals are well-known to be prolific sources itional file 2: Table S1). The latter dataset was used in of natural products with a range of bioactivities, these this study to delineate and compare the prokaryotic, 16S functional assessments were complemented with an rRNA gene-based taxonomic profiles of all samples examination of the abundance, diversity, and (Figs. 1 and 2a, b). Functional analyses were performed Fig. 1 Prokaryotic community composition of octocoral samples, seawater, and sediment. Family-level community profiles of healthy (EG_H) and necrotic (EG_N) Eunicella gazella tissue, healthy Eunicella verrucosa (EV01-EV04), and Leptogorgia sarmentosa (LS06-LS08) specimens as well as seawater (SW01-SW04) and sediment samples (SD01-SD03). Relative abundances are displayed for taxa representing more than 3% of the total dataset reads. Taxa with abundances below 3% across the data are collectively labeled as “rare families”
Keller-Costa et al. Microbiome (2021) 9:72 Page 4 of 21 Fig. 2 Multivariate analysis of the prokaryotic community profiles. Ordinations are shown at the taxonomic (phylotype, OTU) (a and b) and functional (Pfam and COG) (c and d) levels. In a, sediment samples were included in the ordination analysis while b shows the same ordination without sediment data. Principal coordinates analyses (PCoA) were performed using the Bray-Curtis similarity matrix calculated from Hellinger-transformed abundance data. All ordinations are shown in Eigenvalue scale. Healthy octocoral samples are represented by colored circles (salmon — healthy Eunicella gazella (EG15_H - EG18H); orange — healthy Eunicella verrucosa (EV01 - EV04); olive — healthy Leptogorgia sarmentosa (LS06 - LS08)), necrotic E. gazella (EG15_N - EG18_N) by red triangles, sediment (SD01 - SD03) by black diamonds, and seawater (SW01 - SW04) by blue asterisks. Discrete grouping of the sample categories was statistically supported by one-way PERMANOVA tests with each 999 permutations on assembled metagenomes (Additional file 2: Table S2), Distinct alpha-diversity measures revealed for octocoral- using the Integrated Microbial Genomes & Microbiomes associated and environmental prokaryotic communities (IMG/M) annotation pipeline from the Joint Genome Alpha-diversity analyses encompassed 93,589 high-quality Institute (DOE-JGI—see “Methods” for details). For reads classified into 1041 prokaryotic OTUs (12 archaeal more information on the features of the 16S rRNA gene and 1029 bacterial OTUs defined at 97% 16S rRNA gene dataset used in taxonomic profiling and of the metagen- similarity) obtained from the 20 unassembled metagen- ome assemblies subjected to functional analysis, see ome samples (Additional file 2: Tables S3 and S4). Our Additional file 1: extended results. primer-less, shotgun metagenomic approach showed
Keller-Costa et al. Microbiome (2021) 9:72 Page 5 of 21
reduced OTU richness in healthy octocoral specimens and 396 families were detected in seawater (26,039
compared with surrounding environments. OTU richness reads), necrotic E. gazella tissue (19,927 reads), and sedi-
and (Shannon’s) diversity measures were significantly ments (10,449 reads), respectively. This reinforces the
higher (t-test, P = 0.001 and P = 0.0173) in necrotic than notion that host-selection processes occur at taxonomic
in healthy E. gazella tissue (see Additional file 1: extended ranks higher than phylotype (i.e., “OTUs”) level and are
results for details). a major driver of divergent prokaryotic community
structures in octocoral-associated and free-living settings
Octocoral-associated and environmental prokaryotic (Fig. 1). Rhodobacteraceae, Flavobacteriaceae, and Ver-
communities are divergent at high taxonomic ranks rucomicrobiaceae were dominant members of the micro-
Congruent with the notion of highly divergent prokaryotic biomes of seawater and necrotic E. gazella tissue, while
communities being found in distinct habitats, differences Endozoicomonadaceae and uncultivated families OM60
in taxonomic composition of the microbiomes of necrotic and 211ds20 (both in the order Alteromonadales) char-
and healthy octocoral tissue, sediments, and seawater acterized the healthy octocoral tissue across all examined
(hereafter termed “sample categories”) were observed even species (Fig. 1). Several bacterial families showed a more
at high taxonomic ranks (e.g., phylum and class levels). consistent association and higher relative abundances in
Proteobacteria was the most dominant phylum across the L. sarmentosa than in the Eunicella species, including
entire dataset. However, the relative abundance of Proteo- Mycoplasmataceae (Mollicutes, Tenericutes) and Vibrio-
bacteria varied markedly between sample categories, ran- naceae, and so-far unclassified families in the Alteromo-
ging from highly dominant 80 to 98% proportions in nadales and Oceanospirillales orders (Fig. 1).
healthy octocoral tissues (depending on the species) while At the finest level of taxonomic resolution used in this
dropping to 62% in necrotic E. gazella tissue and to 54% study (i.e., OTUs), ordination analysis revealed that the
and 53% in sediment and seawater, respectively. Besides prokaryotic communities of healthy octocoral tissue,
Proteobacteria, Bacteroidetes (28.1%), Verrucomicrobia necrotic E. gazella tissue, sediments, and seawater are
(7.8%), and Cyanobacteria (7.1%) were the most abundant significantly different from one another (Fig. 2a, b).
phyla in the highly consistent taxonomic profiles of sea- While the prokaryotic communities of healthy octocorals
water, while Bacteroidetes (16.9%), Planctomycetes (6.7%), were distinguished from the other sample categories
and Actinobacteria (5.0%) ranked as the next most abun- along the horizontal axis of the ordination diagram, sedi-
dant phyla in sediments. The highest number of prokary- ment samples clustered far apart from all other sample
otic phyla (Additional file 2: Table S3), classes categories along the vertical axis (Fig. 2a). However,
(Additional file 1: Figure S2), families (Fig. 1), and OTUs when sediment samples were omitted from the analyses,
(see above) among all sample categories was found in sed- we could observe the clustering of seawater samples
iments, highlighting the complex nature of these prokary- away from octocoral samples (Fig. 2b). This is congruent
otic communities. Among healthy octocoral tissues, the with the observation that families such as Synechococca-
class Mollicutes in the Tenericutes phylum possessed a ceae, Pelagibacteraceae, Halomonadaceae, and Cryomor-
consistent pattern of association with L. sarmentosa, but phaceae (Fig. 1) were characteristic of seawater but were
not with the two Eunicella species examined here (Add- found to be absent or in very low abundance in octo-
itional file 1: Figure S2). Notably, Poribacteria that are coral samples. Among the healthy tissue samples of all
usually regarded as “sponge-specific” were the only octocoral species, those from L. sarmentosa formed a
phylum that was present in the healthy tissue of all three more cohesive cluster in the ordination diagram than
octocoral species (despite its low relative abundance), but the samples representing both Eunicella species where
absent in necrotic E. gazella, sediment, and seawater. The higher dispersion in community profiles was observed
class Gammaproteobacteria within the Proteobacteria (see below).
phylum dominated the prokaryotic communities in From a total of 1041 OTUs detected across the data,
healthy tissues of all host species, while a drastic shift to- 89 OTUs were common to healthy octocoral tissue, sea-
wards dominance of the classes Alphaproteobacteria (Pro- water, and sediments, of which only 25 OTUs were
teobacteria) and Flavobacteriia (Bacteroidetes) was present in every single metagenome sample from these
observed in necrotic E. gazella tissues (see Additional file 1: groups (Additional file 1: Figure S3a, b). Nine OTUs
extended results for more details on class abundance dis- were shared by healthy tissue samples of all octocoral
tributions across sample categories). species while not being present in any of the sediment
Healthy octocoral samples (all three species together) or seawater samples. Of these, three OTUs stood out,
hosted a total of 220 prokaryotic families (across 37,174 being present in all 10 healthy tissue samples: uncl.
analyzed reads), whereby E. gazella hosted 127 (20,420 Endozoicomonadaceae (OTU_866), Shewanella (OTU_
reads), E. verrucosa 174 (13,435 reads), and L. sarmen- 805), and Pseudoalteromonas (OTU_943). Higher pro-
tosa 123 (3319 reads) families. In comparison, 252, 263, portions of “specific” OTUs were identified in seawaterKeller-Costa et al. Microbiome (2021) 9:72 Page 6 of 21
(28%) and sediment (44%) samples than in the three shared exclusively with healthy E. gazella tissue (Add-
octocoral species, which harbored between 5% (L. sar- itional file 1: Figure S3c, d).
mentosa) and 13% (E. verrucosa) unique, species-specific No significant change in abundance of Vibrio spp. (five
OTUs (Additional file 1: Figure S3a). Vibrio spp. OTUs were present in the dataset) was ob-
served between healthy and necrotic E. gazella tissue
Necrotic octocoral tissue is heavily colonized by typical (paired t-test, P = 0.703). In addition, no Vibrio OTU
seawater bacteria was among those 93 OTUs identified by the “Indicspe-
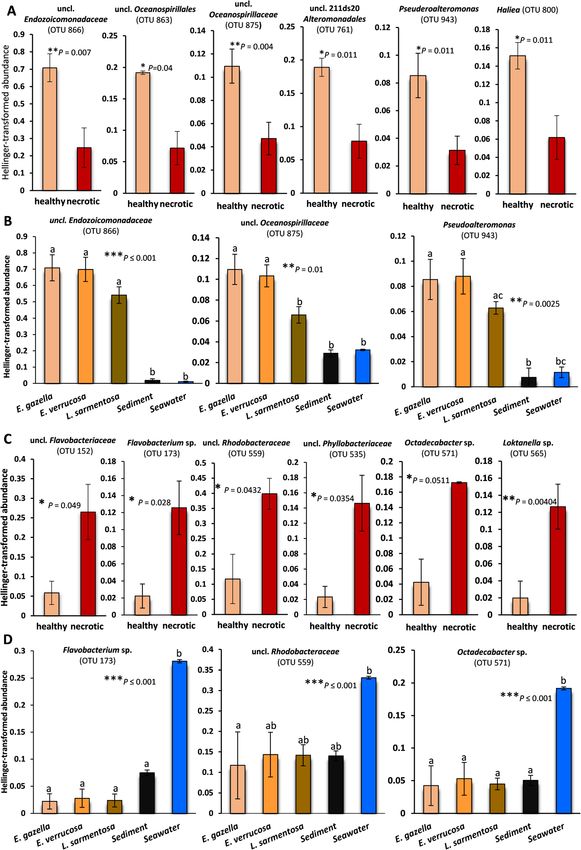
The 10 most enriched OTUs in healthy octocoral tis- cies” analysis to be associated with the necrotic state.
sue—identified by similarity percentage (SIMPER) ana- However, healthy L. sarmentosa samples had a higher
lyses—comprised currently unclassified and uncultured abundance of Vibrio spp. phylotypes (ANOVA, P =
Alteromonadales and Oceanospirillales/Endozoicomona- 0.0035) than healthy E. gazella and E. verrucosa, sedi-
daceae phylotypes, as well as Pseudoalteromonas, Haliea, ment, and seawater. The latter four sample categories
and Mycoplasma sp. (Mollicutes, Tenericutes). While shared similar Vibrio spp. abundances.
most of these phylotypes were usually abundant in all
octocoral species investigated (e.g., OTUs 866, 875, and Functional profiling of prokaryotic communities in
943, representing Endozoicomonadaceae, Oceanospirilla- octocorals and seawater
ceae, and Pseudoalteromonas spp., respectively—Fig. 3), For functional comparisons, we focused on prokaryote-
Mycoplasma sp. (OTU 1008, Additional file 2: Table enriched metagenome assemblies, after eukaryotic con-
S4a) was consistently dominant only in L. sarmentosa. tigs had been filtered out (Additional file 2: Table S5). A
Nevertheless, the relative abundances of these taxa were total of 7742 Pfam and 4570 COG entries were identi-
significantly reduced in necrotic tissues and in the sur- fied. The number of Pfams identified per sample group
rounding sediment and seawater (Fig. 3a, b; Add- ranged from 4286 in seawater to 4887 in healthy L. sar-
itional file 2: Table S4b). Thus, they could be considered mentosa, while the number of COGs ranged from 3473
signatures of the healthy, temperate gorgonian micro- in healthy E. gazella to 3914 in seawater. The full Pfam
biome. This interpretation was further supported by an and COG entry lists for all assembled metagenome sam-
“Indicspecies” [36, 37] analysis, here used to assess the ples, and corresponding ANOVA results for each entry,
association between OTUs and health states in E. gazella are provided as supplementary information (Add-
(Additional file 2: Table S4c). “Indicspecies” returned 17 itional file 2: Table S6 and S7).
OTUs associated with healthy E. gazella tissue and 93
OTUs associated with necrotic E. gazella tissue. From Ordination analysis of functional profiles
those 17 OTUs, nine were also among the top 10 most Both Pfam- and COG-based multivariate analyses (Fig.
differentiating phylotypes identified in our SIMPER test. 2c, d) showed significant differences between sample
Sixteen of the 17 phylotypes found to be associated with categories (one-way PERMANOVA, P < 0.001). Pairwise
healthy tissue by the “indicspecies” analysis belonged to comparisons revealed that seawater functional profiles
Gammaproteobacteria, most often in the orders Ocea- differed from those of each octocoral species in both
nospirillales and Alteromonadales, demonstrating the Pfam- and COG-based annotations. Moreover, in the
high level of congruency between the results of different Pfam-based analysis, we found differences between
statistical tests conducted in this study. some, but not all, octocoral sample groups, with E. ver-
In contrast, various cultivable and so-far uncultivable rucosa profiles being different from L. sarmentosa and
Flavobacteriaceae, Rhodobacteraceae, and Verrucomicro- necrotic E. gazella. In both, COG and Pfam-based func-
biales phylotypes, including transient taxa such as Tenaci- tional analyses, necrotic E. gazella samples clustered
baculum, Flavobacterium, Aquimarina, Octadecabacter, more closely to seawater, suggesting a higher functional
and Loktanella, which have been previously cultivated overlap between these two prokaryotic communities.
from marine hosts [9, 38–41], were enriched in necrotic Healthy octocoral samples (all three species together)
octocoral tissue. While their abundance was generally low showed a more dispersed pattern of distribution in the
in healthy tissue of all three octocoral species, it sharply PCoA plots, pointing towards a higher functional variation
increased in seawater (Fig. 3c, d; Additional file 2: Table and plasticity within the healthy octocoral tissue. In this
S4b). This points to a pronounced colonization of the nec- respect, functional profiles corroborated OTU-level taxo-
rotic octocoral tissue by seawater taxa. Further evidence nomic profiles in which healthy octocoral samples also
for opportunistic colonizers from surrounding environ- showed, altogether, a large within-group variation and
ments taking control of necrotic tissue communities was some degree of overlap between samples from different
provided by Venn diagram analysis. This showed that nec- host species. However, we note that L. sarmentosa sam-
rotic E. gazella tissue shared over 40% of its OTUs exclu- ples formed more concise clusters in both taxonomic- and
sively with seawater and/or sediment, while only 5% were functional-based ordinations (Fig. 2), hinting at a moreKeller-Costa et al. Microbiome (2021) 9:72 Page 7 of 21 Fig. 3 Bacterial phylotypes which are significantly enriched in healthy (a) or necrotic (c) Eunicella gazella tissue. For comparison, the respective abundances of some of these phylotypes (OTUs) in the other octocoral species, sediment, and seawater are shown in b and d. Bars represent average proportions (%) ± standard errors. Paired t-tests (a and c) or one-way ANOVAs (b and d) followed by Bonferroni tests were used to check for significant differences between sample groups. Statistical significance was established at P-values ≤ 0.05. Letters or asterisks above error bars indicate significant differences (*P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001). Respective P-values are presented in the graphs. All phylotypes shown here are among the top ten phylotypes respectively enriched in healthy or in necrotic E. gazella tissue (Additional file 2: Table S4b,c) that contributed most to the dissimilarities between these microbiomes, as revealed by SIMPER and “Indicspecies” analyses selective structuring of prokaryotic communities by this between sample groups in all taxonomic and functional species than by Eunicella spp. Tests on the multivariate analyses (16S-OTU (p = 0.018), COG (p = 0.0065), Pfam homogeneity of group dispersion (variances) corroborated (p = 0.0074)). Pairwise comparisons revealed that the dis- this observation, showing overall significant differences persion among the OTU profiles of the coral sample
Keller-Costa et al. Microbiome (2021) 9:72 Page 8 of 21
groups was similar (homogenous) while the dispersion of for magnesium and cobalt transporters and the iron
seawater profiles was significantly smaller than that of storage proteins ferritin and bacterioferritin were signifi-
healthy and necrotic E. gazella samples. Dispersion among cantly enriched in the healthy octocoral tissue.
COG profiles from seawater samples was significantly All these functions were not only greatly reduced in
smaller than those of healthy coral samples but similar to the microbiome of necrotic E. gazella tissue but also in
those of necrotic coral samples. Overall, the highest levels the surrounding seawater (Additional file 1: Figure S4a).
of dispersion were observed in the microbiomes of healthy By contrast, the microbiomes from healthy E. verrucosa
tissue samples from both Eunicella species, followed by and L. sarmentosa tissue followed a pattern similar to
healthy L. sarmentosa and then necrotic E. gazella, while that observed in the healthy E. gazella samples. How-
the dispersion in seawater was smallest. ever, while the relative abundances of the abovemen-
tioned gene functions in E. verrucosa and E. gazella were
Functional signatures of prokaryotic communities in highly similar, in L. sarmentosa, some functions (e.g.,
healthy octocorals exonuclease III and CRISPR/Cas endonucleases) tended
To identify the functional signatures of the prokaryotic to be of somewhat lower abundance (Additional file 1:
communities in healthy octocoral tissue, we analyzed the Figure S4a).
COG entries that showed overall significant differences
between sample groups based on ANOVA tests per- Functional signatures of prokaryotic communities in
formed with the metagenomics analyses pipeline MGAP necrotic octocoral tissue
of the IMG/M platform from DOE-JGI. Here, we fo- We found many significantly enriched functions in nec-
cused on the COGs that were particularly enriched in rotic E. gazella tissue (when compared to healthy tissue
healthy in comparison with necrotic E. gazella samples samples) that are related to pathogenicity, virulence, and
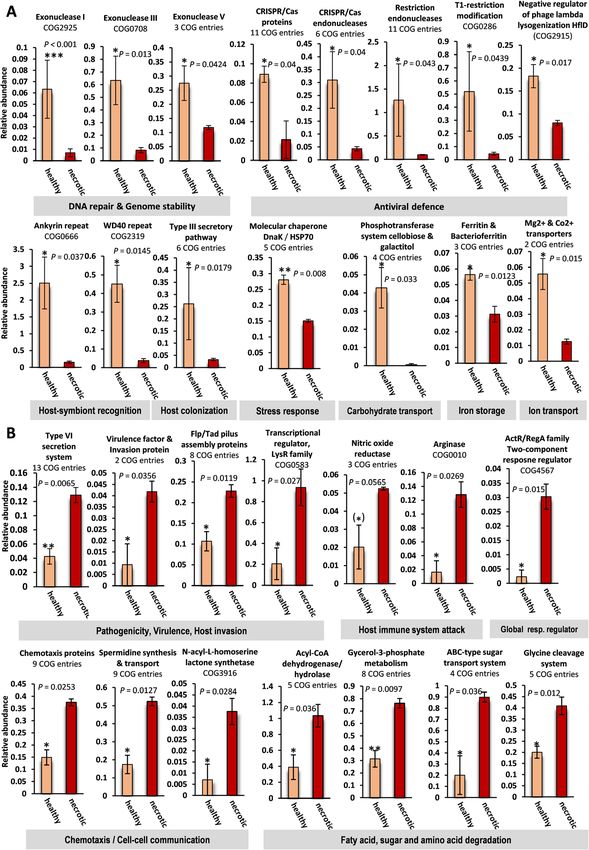
(Additional file 2: Table S7a-c). Abundances of exonu- host invasion. These included 13 COG functions linked
cleases of types I, III, and V involved in DNA repair and to the type VI secretion system and eight COGs affili-
genome stability were higher in the prokaryotic commu- ated with Flp/Tad pilus assembly proteins, two COGs
nities of healthy E. gazella tissue (Fig. 4a) while much linked to the virulence factor B protein, and the invasion
less abundant in necrotic tissue. The prokaryotic com- protein IalB and a LysR type transcriptional regulator
munities of healthy tissue were also significantly (Fig. 4b). The prokaryotic communities of necrotic E.
enriched in a large number of COGs related to antiviral gazella tissue were further defined by an increased abun-
defense. These included multiple COGs related to re- dance of COGs related to chemotaxis protein synthesis,
striction endonucleases, clustered regularly interspaced spermidine (polyamine) synthesis and transport, and N-
short palindromic repeats (CRISPR)/CAS proteins (e.g., acetyl-L-homoserine lactone production. We also found
Cmr3 to Cmr6, Csh2), and CRISPR/CAS endonucleases nitric oxide reductase and arginase-encoding genes
(e.g., Cas1, Cas2, Cas3), as well as two COGs related to a enriched in necrotic E. gazella samples, which help bac-
T1 restriction modification system and a negative regu- terial pathogens to overcome the host’s immune re-
lator of phage lysogenization, respectively. Another fea- sponse. Moreover, the two-component response
ture of the prokaryotic communities of healthy E. regulatory system of the ActR/RegA family—characteris-
gazella tissue were functions encoding for eukaryotic- tic of Rhodobacteraceae bacteria—was also enriched in
like proteins (ELPs), such as ankyrin and WD40 repeats, necrotic tissue samples. Finally, efficient fatty acid, sugar,
which are important for host-symbiont recognition. and amino acid breakdown characterized the prokaryotic
COGs related to the type III secretory pathway, involved communities of necrotic octocoral tissue, as evidenced
in host colonization, were found at higher abundance in by the increased abundance of over 20 COG entries as-
the prokaryotic communities of healthy tissue. The abil- sociated with acyl-CoA dehydrogenases, glycerol-3-
ity to respond to heat and/or heavy metal stress was an- phosphate metabolism, ABC-type sugar transport, and
other characteristic of the healthy octocoral microbiome, glycine cleavage systems. This suggests a feasting of the
evidenced by an increased abundance of genes encoding microbial community on the compromised tissue. Many
for molecular chaperones, particularly the bacterial heat- of these pathogenicity-related functions were signifi-
shock-protein DnaK (COG0443; also known as HSP70). cantly enriched in necrotic samples in comparison with
Genes involved in carbohydrate cycling and micronu- all other surveyed microbiomes (Fig. 4, Additional file 1:
trient acquisition, transport, and storage were another Figure S4 b, c).
signature of the prokaryotic community of healthy E.
gazella tissue. We found several COGs related with Prokaryotic communities in octocorals and seawater
phosphotransferase systems that facilitate the import of encode for distinct secondary metabolisms
sugar dimers and monomers resulting from polysacchar- AntiSMASH analyses predicted 462 secondary metabol-
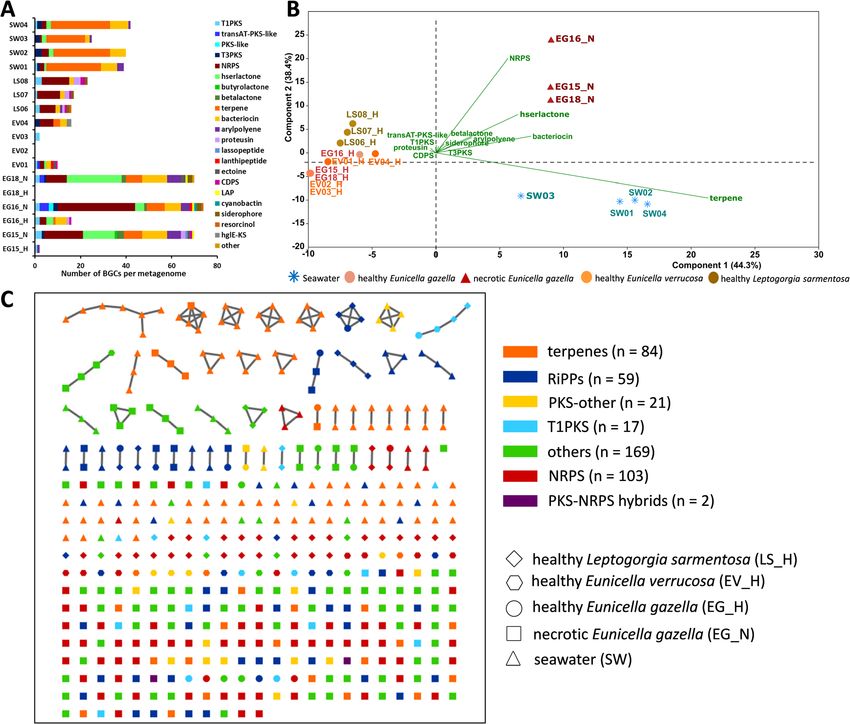
ide breakdown into the cell. Moreover, genes encoding ite BGCs within the 17 assembled metagenomes,Keller-Costa et al. Microbiome (2021) 9:72 Page 9 of 21 Fig. 4 Gene functions which are enriched in the microbiomes of healthy or necrotic Eunicella gazella tissue. Average proportions (%) ± standard errors of Clusters of Orthologous Groups of proteins (COGs) that are significantly augmented in the microbiomes of healthy (a) or necrotic (b) Eunicella gazella tissue. If a given function was represented by more than one COG entry across the dataset, the proportions of these functionally belonging COGs were summed, and the number of COGs that contributed to each bar chart is given below chart titles (see Additional file 2: Table S7(C) for the COG entries used). If only one COG entry contributed to a chart, the respective COG ID is given. Paired t-tests were used to check for significant differences between sample groups. Statistical significance was established at P-values ≤ 0.05. Asterisks above error bars indicate significant differences (*P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001). Respective P-values are presented in the graphs although most BGCs were located at contig edges and (53), and T1PKS (21). The highest number of BGCs was are incomplete. The five most dominant BGCs in the found in necrotic octocoral samples with ≥70 clusters dataset belonged to terpenes (119), followed by NRPS/ per metagenome, followed by seawater samples with up NRPS-like (108), bacteriocins (69), homoserine lactones to 42 clusters per metagenome and healthy L.
Keller-Costa et al. Microbiome (2021) 9:72 Page 10 of 21
sarmentosa with up to 23 clusters. Healthy Eunicella healthy octocoral holobiont, and intricate shifts in func-
samples had a low number of BGCs, ranging from 0 to tional and taxonomic structures which underpin a state
16. Seawater samples were largely dominated by terpene of dysbiosis of the octocoral microbiome. These findings
BGCs; several of which were related to carotenoids, were accompanied by a thorough description of environ-
followed by bacteriocin, T3PKS, and NRPS clusters. mental prokaryotic communities, allowing us to address
Necrotic E. gazella samples possessed the highest diver- the connectivity between microbial taxa and functions
sity of BGCs, which were consistently identified in each across healthy and necrotic octocoral tissue, seawater,
replicate sample, as NRPS, bacteriocin, homoserine lac- and sediments. Our multi-layered approach encom-
tone, aryl-polyene, and terpene BGCs (Fig. 5a, Add- passed comparisons of taxonomic (16S rRNA gene),
itional file 2: Table S8a). The metagenomes of healthy L. functional (COGs and Pfams), and secondary metabolic
sarmentosa were defined predominantly by NRPS and (BGCs) profiles, all of which support the notion of highly
ribosomally synthesized and post-translationally modi- divergent prokaryotic community structures across all
fied peptide (RiPPs; mainly proteusin) BGCs, while Euni- sample categories here examined.
cella samples harbored various polyketide synthase
BGCs, mainly of “type 1”. Corroborating ordination ana- Unclassified and uncultured Oceanospirillales and
lyses of taxonomic and functional profiles (Fig. 2), prin- Alteromonadales taxa characterize the temperate
cipal components analysis (PCA) confirmed a separate octocoral microbiome
grouping of samples into three distinct clusters (healthy Some species of the orders Alteromonadales and Ocea-
octocorals, necrotic E. gazella, and seawater) according nospirillales are known to engage in mutualistic interac-
to their BGC profiles, with high statistical support tions with their hosts. Pseudoalteromonas porphyrae, for
(PERMANOVA: F = 22.04, P = 0.0001; Fig. 5b). Only example, is a marine alga symbiont known to promote
13% (60 in 462) of the predicted BGCs showed some plant growth by stimulating germination and shoot
level of homology with known BGCs publicly available growth through catalase production [42]. Another Pseu-
in the MIBig database, with 31 (7%) BGCs having a simi- doalteromonas sp. strain, OT59, isolated from a healthy
larity value above or equal to 60% (Additional file 2: specimen of the octocoral Leptogorgia alba produces the
Table S8b, c). polyketide alteramide A, a compound that displays light-
The BiG-SCAPE algorithm grouped the BGCs pre- dependent antifungal activity [26]. In the darkness, the
dicted with antiSMASH into 103 NRPS, 84 terpene, 59 strain produces larger quantities of alteramide A which
RiPPs (which include bacteriocins and lantipeptides), 21 inhibits the growth of the putative coral pathogen Peni-
“PKS-other,” 17 type I PKS, two PKS-NRPS hybrids, and cillium citrinum, implying that coral-associated bacteria
169 “other” and non-classifiable “Gene Cluster Families” could protect their host from infections during hetero-
(GCFs) (Fig. 5c, Additional file 2: Table S9). Most of trophic night feeding when coral polyps are most ex-
these families included only a single BGC. GCFs con- posed. Moreover, coral-associated Pseudoalteromonas
taining two or more BGCs were found mainly among spp. were recently shown to function as coral probiotics
terpenes, followed by RiPPs and other, non-classifiable as part of a “beneficial microbes for corals” (BMCs) con-
clusters. Although quite abundant in the octocoral mi- sortium that was able to reduce coral bleaching under
crobial metagenomes, NRPS clusters were mostly unre- temperature and pathogen challenges in aquarium ex-
lated singletons, suggestive of an unprecedented high periments [43].
diversity of NRPS present in these host-associated High abundances of Gammaproteobacteria, comprising
microbiomes. Congruent with trends observed via ordin- specific Endozoicomonadaceae and other Oceanospiril-
ation analysis, most GCFs with more than one BGC ap- lales, Alteromonadales, and Cellvibrionales phylotypes,
peared to be habitat specific rather than shared between were found in this study to be characteristic of healthy
sample types. For example, the majority of terpene GCFs octocoral tissue. Previous surveys recognized the domin-
with two or more BGCs exclusively contained seawater- ance of Endozoicomonas-like phylotypes in many coral-
derived BGCs while the T1-PKS GCFs with more than bacterial assemblages, particularly in temperate octocorals
one BGC were only present in healthy samples of all and Eunicella species [8–13, 16]. Yet here we reveal that
three octocoral species (Fig. 5c). the structure and dynamics of the gammaproteobacterial
consortium associated with healthy corals may be more
Discussion complex than previously thought. In agreement with van
This study is the first cultivation-independent, shotgun de Water et al. [10], we observed that these typical Gam-
metagenome-based survey of the taxonomic and func- maproteobacteria phylotypes may shift in proportions
tional features of prokaryotic communities in octocorals across different host genera (Leptogorgia vs. Eunicella)
and its surrounding environments. Our comparative and greatly reduce in abundance during the transition
analyses revealed many distinct, adaptive traits of the from healthy to necrotic states. Several of the phylotypesKeller-Costa et al. Microbiome (2021) 9:72 Page 11 of 21 Fig. 5 Secondary metabolite coding potential in the prokaryotic communities of healthy and necrotic octocoral samples and seawater. a Distribution of biosynthetic gene clusters (BGCs, N = 462) across these 17 assembled, metagenomes. The BGC counts per compound class were obtained using antiSMASH version 5.0. b Principal component analysis (PCA) biplot based on the BGCs (arrows) found in the metagenomes of the microbiomes of healthy (EG_H, salmon circles) and necrotic (red triangles; EG_N) Eunicella gazella tissue, healthy Eunicella verrucosa (orange circles; EV01 - EV04), healthy Leptogorgia sarmentosa (olive circles; LS06 - LS08), and seawater (blue asterisks; SW01 - SW04). c Similarity network of 455 BGCs predicted by antiSMASH and grouped into biosynthetic gene cluster families (GCFs) across seven major compound classes using the BiG-SCAPE algorithm. The network was rendered in Cytoscape. Nodes represent amino acid sequences of BGC domains, and their different shapes indicate the origin of the sampled metagenome. BGC classes are color-coded with number of GCFs per class given in brackets. A majority of the GCFs (“Others,” N = 169) could not be classified using current BiG-SCAPE BGC nomenclature. Most GCFs were composed by only one BGC (singletons), with GCFs containing two or more BGCs represented through BGC networks inferred from protein sequence homology. PKS, polyketide synthase; NRPS, non-ribosomal peptide synthetase; RiPPs, ribosomally synthesized and post-translationally modified peptides; hserlactone, homoserine lactone; CDPS, tRNA-dependent cyclodipeptide synthases; hglE-KS, heterocyst glycolipid synthase-like PKS; LAP, linear azol(in)e-containing peptides that we found significantly enriched in healthy E. gazella enriched in healthy tissue, only two corresponded to cul- can be considered core members of the microbiome of tured, described species of the genera Pseudoalteromo- temperate gorgonian corals as they were found in all nas and Haliea, respectively. The other seven tested healthy samples of the three octocoral species, com- Gammaproteobacteria on that top-ten list were not clas- plying with the “core microbiome” definition suggested by sifiable at genus or even family level. These phylotypes, van Oppen and Blackall [44]. although belonging to a bacterial class well represented Our primer-less, metagenomic-centered approach re- by a high diversity of cultivatable species, are likely ob- vealed that among the top 10 most distinctive phylotypes ligatory symbionts whose growth requirements are
Keller-Costa et al. Microbiome (2021) 9:72 Page 12 of 21
difficult to meet under standard laboratory conditions. early successional stages. However, further studies, involv-
Moreover, beyond being thus far uncultivatable, these ing more octocoral species and more replicate samples
Gammaproteobacteria spp. are likely missed by ampli- per health condition, are necessary to fully understand
con sequencing strategies which are commonly used in beta-diversity trends in healthy and diseased coral
the description of coral microbiome diversity. For ex- colonies.
ample, the unclassified Oceanospirillaceae and Alteromo- As a clear sign of “dysbiosis” [44] and community
nadales phylotypes as well as the OM60 and 211ds20 destabilization, we found significant changes in micro-
families (Alteromonadales), found in this study to be sig- biome composition correlated with the necrotic tissue
nificantly enriched in healthy octocoral tissue samples, of E. gazella. Such changes were evidenced by a pro-
were not detected in our earlier amplicon-based study of nounced decrease in abundance of the multiple Gam-
the closely related octocoral E. labiata sampled at the maproteobacteria taxa mentioned above with a
same location [9]. In that previous study, the most abun- simultaneous, significant increase in Rhodobactera-
dant OTUs more often belonged to well-known, cultur- ceae, Flavobacteriaceae, and Verrucomicrobiaceae phy-
able genera such as Shewanella, Vibrio, or Ruegeria [9]. lotypes. The latter reached relative abundances similar
This emphasizes the importance of primer-less, metage- to the ones found in seawater, pointing to an intru-
nomics tools in the study of microbiome structuring and sion of typical seawater taxa into the necrotic octo-
functioning in nature as to avoid cultivation and primer coral tissue. Increased abundances of opportunistic
bias. Finally, both our taxonomic and functional analyses Rhodobacteraceae and/or Flavobacteriaceae were pre-
point towards a correlation of octocoral microbiome viously reported for hard corals at anthropogenically
structure with host phylogeny. This trend is commonly impacted sites, under climate change stressors [48,
referred to as “phylosymbiosis” which, in this study, was 49] and associated with white band [50] and grey
detectable at the genus—but not species—level owing to patch disease [47]. However, in contrast to previous,
the distinctive features of Leptogorgia and Eunicella pro- bacterial cultivation-based investigations of temperate
karyotic communities (Additional file 1: extended octocorals which suggested an infectious disease
discussion). caused by Vibrio pathogens as the necrosis-causing
agent [31, 32], our cultivation-independent, primer-
Complex structural shifts characterize dysbiosis of the less metagenomics approach did not show increased
octocoral microbiome Vibrio abundances in the microbiomes of necrotic E.
Despite a lower alpha-diversity (suggestive of host-driven gazella tissue when compared to healthy tissue. Ra-
control of microbiome assembly in the healthy state), sur- ther, dysbiosis in E. gazella involves complex struc-
prisingly, we found a higher beta-diversity (i.e., dispersal tural changes and abundance shifts of bacterial
in community structure among samples) in healthy octo- populations. In agreement with previous findings [20,
coral tissue as compared to necrotic E. gazella tissue, both 51], we therefore advocate for a re-evaluation of the
at the taxonomic and functional gene content level. This “Vibrio-paradigm” in future studies. These may com-
would contrast the lately postulated “Anna Karenina bine molecular and cultivation-based detection of mi-
principle,” which argues that dysbiotic individuals vary crobial pathogens from diseased octocoral tissue to
more in microbial community composition than healthy assess cause-effect relationships using infection exper-
individuals since the microbiological changes induced by iments in microcosms. It is now known that regular
many perturbations are stochastic, leading to unstable cultivation procedures usually favor the growth of low
community states [45, 46]. Yet lower community similar- abundance, but rather opportunistic, bacteria from
ity between healthy scleractinian corals was also reported complex microbiomes as divergent as those from soils
in a recent 16S amplicon sequencing study on a new [52] and marine sponges [38, 53]. Our data suggest
“grey-patch” disease [47]. The higher beta-diversity ob- that a similar pattern is likely to occur for fast-
served in the study of Sweet et al. [47] and in this study growing, cultivatable Vibrio spp. from coral micro-
may be accounted for because of the long lifespans of biomes. Their eventual absence or low representation
many corals which result in structural adjustments of their in amplicon-sequencing datasets [34] may not neces-
microbiome to maintain holobiont fitness under changing sarily derive from primer bias, but from the fact that
micro-environmental conditions. Alternatively, our results they are simply low abundant across several animal-
suggest that the process of necrosis of the octocoral tissue microbe associations as suggested elsewhere for fish
may follow deterministic succession events, leading to [54] and marine sponges [53]. Future surveys on dif-
convergent community structures across different individ- ferent host species, sampling locations, and seasons
uals. We speculate that this might be the case particularly will shed light on whether the pattern of dysbiosis
for late stages of necrosis, after the starting decaying tissue observed here is universal across coral-microbiome
had been firstly colonized by eventual opportunists in associations.Keller-Costa et al. Microbiome (2021) 9:72 Page 13 of 21 DNA stabilization mechanisms and antiviral defense are High abundances of eukaryotic-like repeat protein- hallmarks of octocoral-microbiome symbiosis encoding genes may facilitate host-symbiont recognition Our study shows that the functional profiles of prokary- in the octocoral holobiont otic communities in healthy tissue of the three octocoral The prevalence of eukaryotic-like proteins (ELPs), species (albeit quite variable within Eunicella specimens) namely ankyrin and WD40 repeats, likely underpins the were distinctive from necrotic octocoral tissue and sea- mechanisms through which host-symbiont recognition water. Exonucleases I, III, and V, found at higher abun- and a stable partnership with the host are enabled within dance in healthy samples of the three octocoral species, octocorals. Ankyrin repeats were by far the most abun- have been thoroughly characterized in the model bacter- dant type of ELPs in the healthy E. gazella microbiome, ium Escherichia coli [55]. Exonucleases I and III are typ- and their role in preventing phagocytosis of bacteria by ically involved in dsDNA repair and homologous eukaryotic cells has been documented experimentally recombination, including the repair of oxidative and ion- [62]. Given their presumed eukaryotic origin, the pres- izing radiation damage, which may contribute to a ence of ELPs in bacterial genomes is interpreted as the higher plasticity of the microbiome of healthy octocorals result of lateral host-microbe gene transfer. The estab- to respond to environmental stressors. In addition, the lishment of mutualistic interactions also requires the highly processive exonuclease V possesses also an endo- suppression of local immune responses to allow success- nuclease function which restricts bacteriophage infec- ful colonization of the host. Although type III secretion tions [55]. Indeed, we here reveal for the first time that systems have primarily received attention due to their the prokaryotic consortium of healthy octocorals main- role in host colonization by pathogenic bacteria, there is tains a sophisticated antiviral defense arsenal, compris- growing evidence that beneficial bacteria also use this ing not only the type V exonuclease but also multiple system to secrete immune-suppressive effectors into host restriction endonucleases together with a complex CRIS cells to facilitate host colonization [63]. The enrichment PR/Cas system and phage lysogenization regulators. of type III secretion system encoding genes in the micro- Since an enrichment of CRISPR/Cas endonucleases was biomes of healthy octocoral tissue observed in this study common to all specimens of the three octocoral species supports this notion. studied here, we argue that CRISPR-associated proteins are a true hallmark of the healthy, temperate octocoral The octocoral microbiome is enriched in abiotic stress microbiome. The CRISPR/Cas machinery functions as response mechanisms an efficient prokaryotic immune system that transmits As a likely adaptative response to heat, oxidative, and/or resistance to bacteriophages and other foreign genetic other abiotic stressors, we found an enrichment of genes elements such as plasmids [56], permitting bacterial encoding for the molecular chaperone DnaK (HSP70) in survival within a densely populated holobiont. A sym- the prokaryotic consortium of healthy tissue from all biotic community well protected from viral infections three octocoral species. HSP70-like heat shock proteins and foreign DNA that is brought in through the are ubiquitous in nature, present in most if not all mi- suspension-feeding activity of the octocoral polyps is cro- and macro-organisms and have been reported to be likely more stable, and such improved defense mecha- highly expressed, for example, by the marine bacterium nisms may be a prerequisite for lasting interactions Vibrio harveyi as well as the stony coral Stylophora pis- with the host. Although undescribed in octocorals, a tillata in response to in vivo heat exposure [64, 65]. This higher incidence of CRISPR-Cas and restriction endo- finding suggests plasticity of the octocoral microbiome nucleases has been reported in earlier metagenomics to respond to environmental stressors. We also found surveys on the microbiomes of healthy, marine evidence that the prokaryotic community in healthy E. sponge specimens [57, 58]. Recent analyses of viromes gazella tissue could contribute with specific micronutri- in hard corals revealed lower viral loads in these ents to the holobiont, such as iron, cobalt, and magne- hosts than in seawater in spite of higher bacterial sium. The latter are essential cofactors for numerous abundances in the coral host. This suggests a prevail- enzymes. Cobalt, for example, is an essential factor of ing lysogenic, instead of lytic, mode of interaction be- vitamin B12 (cobalamin) which is required by eukaryotes tween coral symbionts and their bacteriophages [59, for growth and synthesized by gastric cavity bacteria in 60]. However, a sophisticated CRISPR/Cas defense the semi-closed gastrovascular system of corals [17]. The system and potential prophage was also identified on supply of B vitamins, among other nutrients, was also a the genomes of two Cyanobacteria species isolated key feature identified on the metagenome-assembled ge- from black band diseased coral [61]. Future studies of nomes of bacterial symbionts from the scleractinian the octocoral virome hold promise in revealing the coral Porites lutea [66]. Indeed, biosynthesis pathways role of bacteriophages in mediating host-microbe in- for cobalamin (B12) and other B vitamins are also a teractions within these animals. common feature of alphaproteobacterial symbionts of
Keller-Costa et al. Microbiome (2021) 9:72 Page 14 of 21
marine sponges [38], suggesting that these pathways activity, and protection from oxidative and acid stress
could be generally important for marine invertebrate [71]. Future research shall invest in linking function with
growth. taxonomy through the analysis of metagenome-
assembled genomes (MAG) to associate the pathogenic
Pathogenicity features and potential to silence the host features and virulence factors described here with spe-
immune response revealed for prokaryotic assemblages cific, putative coral pathogens.
in necrotic octocoral tissue
Many metabolic features related to macronutrition, par- Differential natural product biosynthesis capacities
ticularly lipid, amino acid, and sugar degradation, were revealed for octocoral and seawater microbiomes
enriched in necrotic octocoral tissue. This suggests an Octocorals are prolific sources of natural products with
enhanced catabolic capacity of the prokaryotic consor- fascinating and unusual chemical structures and bioac-
tium from necrotic E. gazella samples to consume the tivities of interest to medicine and biotechnology. Des-
dying octocoral tissue and organic matter made avail- pite growing evidence from culture-based studies for an
able. The necrotic tissue was largely characterized by existing role of microbial associates in the production of
pathogenic features that facilitate host invasion, viru- some of these bioactive compounds [28], a mechanistic
lence, and host immune system attack, which are pos- understanding of the contribution of the octocoral
sibly tightly regulated by quorum-sensing mechanisms, microbiome (including uncultured symbionts) to the
as evidenced by an increased abundance of homoserine chemical diversity of octocorals is lacking. In this study,
lactone synthetase-encoding genes. we illuminate the natural product biosynthesis potential
This opportunistic community of prokaryotes further within octocoral-associated and free-living prokaryotic
shows the ability to overcome attacks by the host’s im- communities and show higher numbers and diversity of
mune system by quenching its nitric oxide (NO) produc- natural product BGCs in the metagenomes of necrotic
tion through nitric oxide reductase and arginase activity. octocoral tissue than in healthy tissue and seawater. Our
NO production is a typical cellular stress response in results suggest that bacterial colonizers of necrotic octo-
cnidarians that attempts to kill pathogens with a nitric coral tissue are well equipped to compete with resident
oxide burst [67]. Oxidative stress has been reported as a microbes and to damage the host-tissue through anti-
driver of scleractinian coral-Symbiodiniaceae mutualism microbial compounds and toxin production. However,
breakdown, and components of the integrated stress re- we also found a high diversity of NRPS and RiPPs en-
sponse pathway, including reactive species such as NO, coding BGCs in the prokaryotic metagenomes of healthy
have been extensively detected in coral holobionts as L. sarmentosa specimens, while seawater communities
stress responses to warm water and infection [67, 68]. were shown to be prolific sources of diverse and novel
Yet several pathogens possess sophisticated mechanisms terpene BGCs. Intriguingly, within each compound class,
to counteract NO production [69]. A crucial amino acid these BGCs were mostly unrelated with one another,
that modulates the cellular immune response during in- possessing dissimilar gene composition and architecture
fection is arginine, which is the common substrate for (i.e., synteny), and did not show significant similarity to
both inducible nitric oxide synthase (iNOS) and argi- known BGCs in public databases. This highlights the
nase, as well as spermidine production. Generation of amount of genetic novelty likely present in both free-
NO from arginine is responsible for efficient immune re- living and host-associated marine microbiomes and pin-
sponse and cytotoxicity of host cells to kill invading points the L. sarmentosa holobiont and bacterioplankton
pathogens. But the conversion of arginine to ornithine prokaryotes as possible targets for future bioprospection
and urea via the arginase pathway drains the substrate studies.
necessary for NO production from the host and supports Several of the BGCs found in necrotic tissue belonged
the growth of bacterial pathogens [69]. Competition be- to Flavobacteriaceae and Rhodobacteraceae which are
tween iNOS and arginase for arginine can thus contrib- phylotypes significantly enriched in these samples. For
ute to the outcome of several bacterial infections. example, an aryl polyene/resorcinol gene cluster from
Moreover, high arginase activity will result in ornithine necrotic tissue samples shared a high degree of similarity
production, an essential substrate for the polyamine with flexirubin, a pigment typical of Flavobacteria spe-
pathway, leading to putrescine and spermidine synthesis cies, displaying highest homology to the flexirubin-
[70]. In line with this, we found multiple genes related encoding BGC of Flavivirga. We further identified a
to spermidine synthesis and transport enriched in the BGC encoding for the siderophore bisucaberin B in nec-
metagenomes of necrotic tissue. Spermidine has multiple rotic tissue samples with a closest sequence match to
functions in bacterial pathogenicity, including the induc- Aquimarina, another Flavobacteriaceae genus found in
tion of host cell apoptosis, biofilm formation, escape increased abundance in necrotic octocoral tissue. Bisuca-
from phagolysosomes, bacteriocin production, toxin berin B has previously been isolated from the sponge-You can also read

















































