Life in the extreme environment: Structure and species richness of bird assemblages on Yuzhny Island of Novaya Zemlya, Russia - Biotaxa
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ecologica Montenegrina 39: 46-58 (2021)
This journal is available online at: www.biotaxa.org/em
http://dx.doi.org/10.37828/em.2021.39.5
Life in the extreme environment: Structure and species richness of
bird assemblages on Yuzhny Island of Novaya Zemlya, Russia
VITALY M. SPITSYN*, YAROSLAVA E. KOGUT & IVAN N. BOLOTOV
N. Laverov Federal Center for Integrated Arctic Research of the Ural Branch of the Russian Academy of Sciences,
Northern Dvina Emb. 23, 163000, Arkhangelsk, Russia
*Corresponding author: spitsyn.v.m.91993@yandex.ru
Received 2 January 2021 │ Accepted by V. Pešić: 26 January 2021 │ Published online 28 January 2021.
Abstract
Birds play a vital role in arctic environments, being multi-functional ecosystem engineers, but these animals are heavily
impacted by recent climate warming. Bird assemblages on the Arctic Ocean archipelagoes are poorly known, because
many such areas are hardly accessible to scientists. Novaya Zemlya, one of the most enigmatic places in the World, was
a closed military area from the late 1940s. This gigantic mountainous archipelago can be considered a terra incognita
by means of modern faunal, taxonomic, and ecological research. In the present study, we provide the first qualitative
data on bird assemblages of the Yuzhny Island of Novaya Zemlya, estimate the diversity of bird species through a range
of habitats, and underscore environmental factors determining the spatial distribution of avifauna in the arctic tundra
biome. In terrestrial habitats, Tundra Bean Goose (Anser fabalis rossicus), Barnacle Goose (Branta leucopsis) and
Snow Bunting (Plectrophenax nivalis) were the most abundant species. In freshwater and coastal marine habitats, both
these Arctic-breeding goose taxa, and Black-legged Kittiwake (Rissa tridactyla), Common Eider (Somateria
mollissima) and Thick-billed Murre (Uria lomvia) were the dominant species. The most species-rich bird assemblages
(11-15 species) were associated with willow tundra, freshwater lakes, and coastal sea habitats, while only a few species
were recorded in dry rocky habitats, open sea environments, and littoral areas of lakes and the sea. Mountain rocky
heathlands covering most of the area of Yuzhny Island were scarcely populated by birds, with only a few species
recorded frequently there, such as the Rough-legged Buzzard (Buteo lagopus) and Snow Bunting. Our findings
highlight that the bird assemblages on Novaya Zemlya share low species richness and that these assemblages contain a
large proportion of sea and shore bird species even in terrestrial habitats. Among the terrestrial birds, only four cold-
tolerant, common species successfully colonize these extreme environments during the short summer season.
Key words: Species diversity, extreme environment, avifaunal assemblages, migratory species, arctic-breeding birds,
Arctic Ocean archipelagoes.
Introduction
Recent climate changes significantly affect a variety of animals throughout the Arctic, including birds
(Chapin et al. 2005; Black et al. 2012; Robinson et al. 2014; Descamps et al. 2017). The structure of bird
assemblages in polar areas has changed rapidly due to range shifts of widespread generalist species (Thomas
Ecologica Montenegrina, 39, 2021, 46-58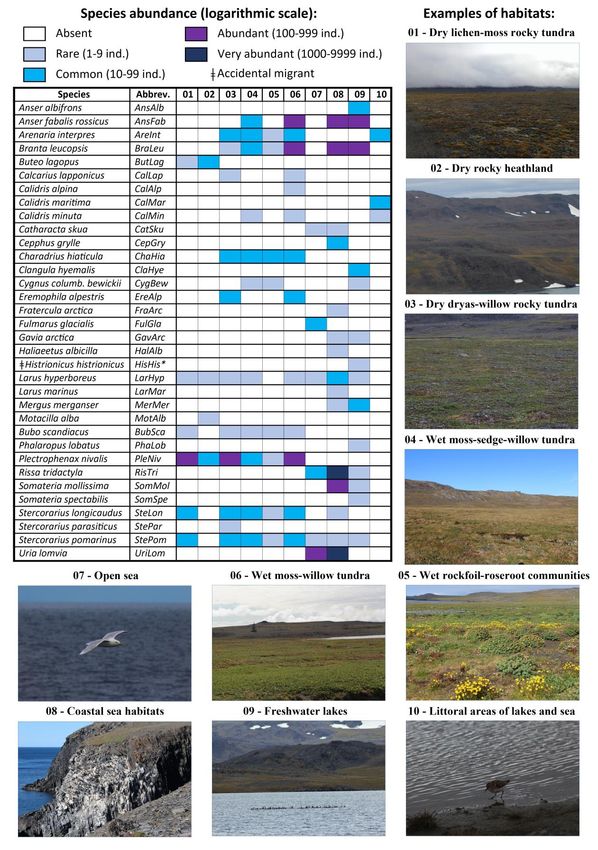
SPITSYN ET AL.
and Lennon 1999; Davey et al. 2012; Sokolov et al. 2012) and higher intensity of migrations towards the
Arctic (Black et al. 2012).
The distribution and species richness of birds on the Arctic Ocean islands are rather poorly known. A
rather large body of literature describes avifaunal assemblages and species biology throughout the North
American Arctic, e.g. the Canadian Arctic Archipelago, Greenland, and the mainland (Freedman and
Svoboda 1982; Lepage et al. 1998; Henry and Mico 2002; Latour et al. 2005; Andres 2006; Trefry et al.
2010; Black et al. 2012; Burnham et al. 2014; Kardynal et al. 2015). In contrast, islands of the European
Arctic (e.g. Svalbard) have received less attention (Jakubiec 1982; Wojczulanis-Jakubas et al. 2008;
Descamps et al. 2017). Moreover, sea bird colonies are the most popular object at the circumpolar scale
(Strøm 1994; Anker-Nilssen et al. 2000; Dickson and Gilchrist 2002; Diemer et al. 2011; Stempniewicz et al.
2017). Reports on avifaunal assemblages from islands of the Russian Arctic are virtually absent, although a
few works on those from mainland areas are available (Gilg et al. 2000; Sokolov et al. 2012).
Novaya Zemlya was a closed military area for almost half a century, but despite this fact, the
avifauna of this archipelago is relatively well-studied (e.g., Pearson 1899; Kalyakin 1993; Pokrovskaya and
Tertitsky 1993; Strøm et al. 1994; Anker-Nilssen et al. 2000; Tertitsky and Pokrovskaya 2011; Spitsyn et al.
2018, 2020). However, any data on bird assemblages of Novaya Zemlya is unavailable, except a few
evaluations of bird density and abundance (Tertitsky and Pokrovskaya 2011; Spitsyn et al. 2018).
Additionally, several papers described the range and abundance of sea bird colonies on Novaya Zemlya
(Pokrovskaya and Tertitsky 1993; Strøm et al. 1994; Anker-Nilssen et al. 2000; Rozenfeld & Spitsyn 2017).
This study aims to (1) analyze the structure of bird assemblages on the Yuzhny Island of Novaya
Zemlya, and (2) estimate environmental factors determining the species richness and spatial distribution of
birds in these habitats. To the best of our knowledge, this work represents the most comprehensive report on
the ecology of bird assemblages on the archipelago published to date.
Material and methods
Data sampling
V. M. Spitsyn collected the primary data for this study (see Tables 1-4). The fieldwork was performed on the
western coast of the Yuzhny Island of Novaya Zemlya (arctic tundra biome: Walker 2000) during the period
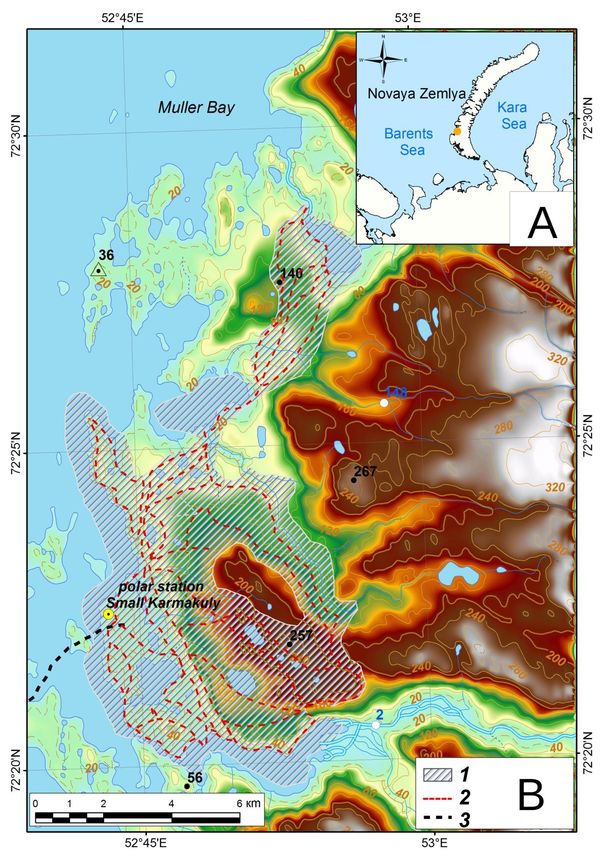
of 16 July – 12 August 2015 (Figs 1A and 1B). The study region is situated around the Malye Karmakuly
Polar Research Station (72°22´ N, 52°43´ E). Its total area is 50.5 km 2, including 40.5 km2 of terrestrial
areas, 3.7 km2 of freshwater areas and 6.3 km2 of coastal sea areas (Fig. 1B). In this region, we selected 10
habitat types as follows: (01) dry lichen-moss rocky tundra, (02) dry rocky highland heathland, (03) dry
dryas-willow rocky tundra, (04) wet moss-sedge-willow tundra, (05) wet rockfoil-roseroot communities, (06)
wet moss-willow tundra, (07) open sea area, (08) coastal sea habitats, including cliff and coastal sea areas,
(09) freshwater lakes, and (10) littoral areas of lakes and sea (Spitsyn et al. 2018). Environmental
characteristics of each habitat are presented in Table 4.
The counting of birds on the routes crossing different habitat patches (Fig. 1B) was performed daily
except for a few days with extreme weather conditions, during which wind speed exceeded 25 m/s. The
routes covered almost the entire study region, including some routes situated along the coastal areas. The
counting unit was an adult bird. Locations of each large flock of aquatic birds (e.g. various geese) and each
colony of sea birds were identified. We performed observations on the abundance and behavior of each large
waterfowl flock/sea bird colony separately. The counting unit was an adult bird present in a flock/colony as
done by Strøm et al. (1994). In the mountains, we observed birds on routes at the bottom of valleys with the
counting on all surrounding slopes (visibility of 300-500 m in each direction). For the counting of coastal
bird colonies, the approach of Strøm et al. (1994) was applied. In total, 21 daily bird counts were performed
on the 10-38 km long routes. Additional counts of sea birds were performed from the research vessels
“Professor Molchanov” and “Mikhail Somov” on 16 July 2015 and 12 August 2015, respectively. On 11
August 2015, additional counting of aquatic birds was performed using a helicopter. Following the approach
of Black et al. (2012), the number of birds observed every day was not an exact count, but an estimate based
on either the frequency with which an individual was seen, or the average size of a flock of birds. Based on
all the counts, we provide approximate estimations of every species abundance within each habitat (Tables 1-
2). The approximate abundances were transformed into categorical abundance values (Table 3) in
Ecologica Montenegrina, 39, 2021, 46-58 47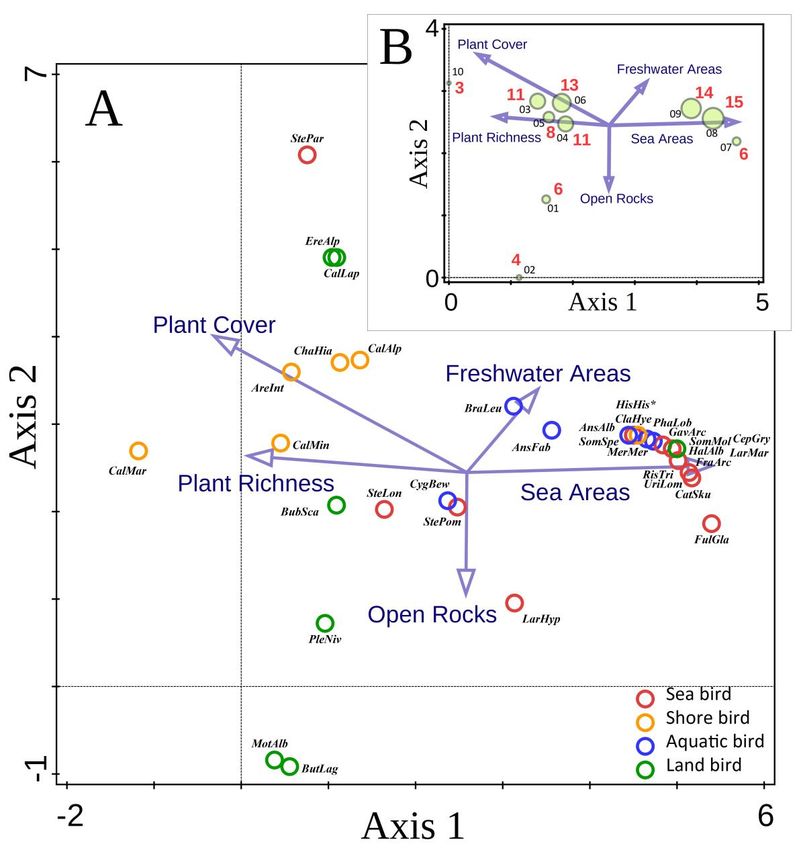
BIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
accordance with a five-point scale, which includes zero and four logarithmically (log 10) scaled categories
(Howard et al. 2015).
Figure 1. Study region on Yuzhny Island of Novaya Zemlya. (A) Map of Novaya Zemlya. The yellow circle indicates
the study region of Yuzhny Island. (B) Detailed map of the study region with the location of counting routes. The
shaded area represents the study area (1). The dashed red lines indicate the counting routes (2). The dashed black line
indicates the helicopter route (3).
48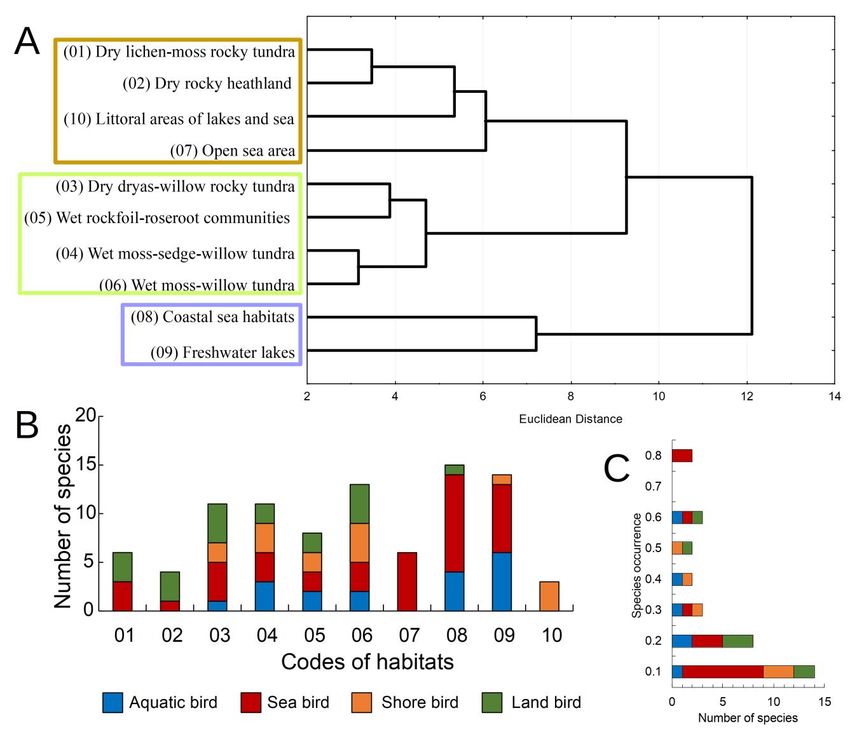
SPITSYN ET AL.
Table 1. Summary data of the counting of birds during the period of 16 July – 12 August 2015 near the Malye
Karmakuly Polar Station, Yuzhny Island, Novaya Zemlya, with notes on breeding status of these taxa.
Breeding
Breeding Approximate
in the Relative Species
on total
study total occurrence
Ecological Novaya abundance
Bird species region abundance over habitat
group Zemlya within the
(Spitsyn et (log10 range (N =
(Kalyakin study region,
al. 2018, scale)** 10)
1993) ind.*
2020)
Anser albifrons Aquatic bird Yes Yes 35 2 0.1
Anser fabalis rossicus Aquatic bird Yes Yes 635 3 0.4
Arenaria interpres Shore bird Probable YesBIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
Table 2. Approximate number of encountered individuals for each bird species over the habitat range during the period
of 16 July – 12 August 2015 based on observations near the Malye Karmakuly Polar Station, Yuzhny Island, Novaya
Zemlya.
Ecological Habitat code*
Bird species
group 01 02 03 04 05 06 07 08 09 10
Anser albifrons Aquatic bird 0 0 0 0 0 0 0 0 35 0
Anser fabalis rossicus Aquatic bird 0 0 0 20 0
Arenaria interpres Shore bird 0 0SPITSYN ET AL.
Table 3. Categorical bird species abundances (log10 scale)* over the habitat range based on observations near the Malye
Karmakuly Polar Station, Yuzhny Island, Novaya Zemlya, during the period of 16 July – 12 August 2015.
Habitat code**
Bird species Ecological group
01 02 03 04 05 06 07 08 09 10
Anser albifrons Aquatic bird 0 0 0 0 0 0 0 0 2 0
Anser fabalis rossicus Aquatic bird 0 0 0 2 0 3 0 3 3 0
Arenaria interpres Shore bird 0 0 2 2 1 2 0 0 0 2
Branta leucopsis Aquatic bird 0 0 1 2 1 3 0 3 3 0
Bubo scandiacus Land bird 1 0 1 1 1 1 0 0 0 0
Buteo lagopus Land bird 1 2 0 0 0 0 0 0 0 0
Calcarius lapponicus Land bird 0 0 1 0 0 1 0 0 0 0
Calidris alpina Shore bird 0 0 0 0 0 1 0 0 0 0
Calidris maritima Shore bird 0 0 0 0 0 0 0 0 0 2
Calidris minuta Shore bird 0 0 0 1 0 1 0 0 0 1
Catharacta skua Sea bird 0 0 0 0 0 0 1 1 0 0
Cepphus grylle Sea bird 0 0 0 0 0 0 0 2 0 0
Charadrius hiaticula Shore bird 0 0 2 2 2 2 0 0 0 0
Clangula hyemalis Sea bird 0 0 0 0 0 0 0 0 2 0
Cygnus columbianus bewickii Aquatic bird 0 0 0 1 1 0 0 0 1 0
Eremophila alpestris Land bird 0 0 2 0 0 2 0 0 0 0
Fratercula arctica Sea bird 0 0 0 0 0 0 0 1 0 0
Fulmarus glacialis Sea bird 0 0 0 0 0 0 2 0 0 0
Gavia arctica Aquatic bird 0 0 0 0 0 0 0 1 1 0
Haliaeetus albicilla Land bird 0 0 0 0 0 0 0 1 0 0
ǂHistrionicus histrionicus Sea bird 0 0 0 0 0 0 0 0 1 0
Larus hyperboreus Sea bird 1 1 1 1 0 1 1 2 1 0
Larus marinus Sea bird 0 0 0 0 0 0 0 1 0 0
Mergus merganser Aquatic bird 0 0 0 0 0 0 0 1 2 0
Motacilla alba Land bird 0 1 0 0 0 0 0 0 0 0
Phalaropus lobatus Shore bird 0 0 0 0 0 0 0 0 1 0
Plectrophenax nivalis Land bird 3 2 3 2 1 3 0 0 0 0
Rissa tridactyla Sea bird 0 0 0 0 0 0 2 4 1 0
Somateria mollissima Sea bird 0 0 0 0 0 0 0 3 2 0
Somateria spectabilis Sea bird 0 0 0 0 0 0 0 0 1 0
Stercorarius longicaudus Sea bird 2 0 2 2 1 2 0 1 0 0
Stercorarius parasiticus Sea bird 0 0 1 0 0 0 0 0 0 0
Stercorarius pomarinus Sea bird 2 0 2 2 1 2 1 1 1 0
Uria lomvia Sea bird 0 0 0 0 0 0 3 4 0 0
*Categories of logarithmic abundance scale: 0 – absent; 1 – rare species [1-9 individuals per habitat patch]; 2 – common
species [10-99 individuals per habitat patch]; 3 – abundant species [100-999 individuals per habitat patch]; and 4 – very
abundant species [1000–9999 individuals per habitat patch]. **Habitat types: (01) dry lichen-moss rocky tundra; (02)
dry rocky highland heathland; (03) dry dryas-willow rocky tundra; (04) wet moss-sedge-willow tundra; (05) wet
rockfoil-roseroot communities; (06) wet moss-willow tundra; (07) open sea area; (08) coastal sea habitats, including the
cliff and coastal sea areas; (09) freshwater lakes; and (10) littoral areas of lakes and sea. ǂAccidental migrant.
Ecologica Montenegrina, 39, 2021, 46-58 51BIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
Table 4. List of environmental variables, which were used in DCA analyses and an approximate square of habitat
patches.
Approximate Presence
Plant Presence Presence Plant
Habitat square of of
Habitat type cover of sea of rocky (floristic)
Code habitat patch, freshwater
(%) areas outcrops richness
km2 areas
01 Dry lichen-moss rocky 20 20 0 0 1 1
tundra
02 Dry rocky highland 8 5 0 0 1 1
heathland
03 Dry dryas-willow rocky 4 55 0 0 1 1
tundra
04 Wet moss-sedge-willow 4 95 0 0 0 2
tundra
05 Wet rockfoil-roserootSPITSYN ET AL.
The most species-rich bird assemblages (11-15 species) were associated with willow tundra,
freshwater lakes and coastal sea habitats, while a few species were recorded in dry rocky habitats, open sea,
and littoral areas of lakes and sea (Figs. 3A and 3B). Mountain rocky heathlands covering most of the
Yuzhny Island were scarcely populated by birds, with only four species being frequently recorded there. The
Rough-legged Buzzard (Buteo lagopus) and Snow Bunting were found to be common species in rocky
heathlands. Sea and shore bird species were presented in most surveyed assemblages (Figs. 2B and 3A).
Three sea bird species revealed the widest distribution over habitat patches, i.e. the Pomarine Skua
(Stercorarius pomarinus), Long-tailed Jaeger (Stercorarius longicaudus) and Glaucous Gull (Larus
hyperboreus) (Fig. 2C). The Barnacle Goose and Snow Bunting were most widely distributed throughout
various habitats among aquatic and land species, respectively (Fig. 2C).
Our cluster analysis uncovered three groups of relatively similar assemblages: (i) species-rich
assemblages, which were associated with vegetation-rich habitats, i.e., willow tundra and herb communities;
(ii) species-poor assemblages of dry rocky habitats, littoral areas, and open sea; and (iii) species-rich
assemblages of coastal sea habitats and freshwater lakes, which contained an amalgam of aquatic and sea
bird species (Fig. 2A).
Figure 2. Structure of bird assemblages on Yuzhny Island of Novaya Zemlya. (A) Cluster analysis of bird assemblages.
Ward’s classification approach was applied to a Euclidean distance matrix, which was calculated based on bird species
abundances (categorical estimations by using a five-point logarithmic scale). (B)-(C) Histograms of the distribution of
bird ecological groups (number of species in each group) over the range of habitats (B) and over relative occurrence of
each species through habitats (C).
Ecologica Montenegrina, 39, 2021, 46-58 53BIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
Among environmental variables, plant cover and floristic richness were important factors in
terrestrial habitats, with the most diverse bird assemblages in sites with the maximum values of these
parameters ( >90% of the total area covered by plants; plant cover contained herbs, sedges, and willows)
(Figs 3A-3B). Conversely, the lack of continuous plant cover in open rocky areas significantly decreased the
species richness of bird assemblages. The highest richness was discovered in habitats associated with
freshwater and seawater areas, such as freshwater lakes and coastal sea sites (Figs 3A-3B).
Figure 3. Species diversity of bird assemblages on Yuzhny Island of Novaya Zemlya. (A) Bi-plot of detrended
correspondence analysis (DCA) with supplementary variables, showing the ordination of species and environmental
variables. Circles indicate bird species abundance (categorical estimations by using a logarithmic scale, see Table 3),
abundances decrease with increasing distance from each point in a unimodal fashion (ter Braak and Smilauer, 2002).
Data represent independent samples from various habitats (n = 10). Total variation is 2.44, supplementary variables
account for 66.1% (adjusted explained variation is 23.8%). Eigenvalues (lambda) are 0.675, 0.162, 0.069, and 0.025 for
first (horizontal), second (vertical), third and fourth axes, respectively. The first two axes explain 34.4% of the
variation. The pseudo-canonical correlations of bird abundance and environmental variables for axes 1 and 2 are 0.77
and 0.91, respectively. For an explanation of environmental variables, see Table 4. For abbreviations of species names
see Fig. 4. (B) Bi-plot of the same analysis revealing the ordination of species richness over a range of habitats and
environmental variables. Circles indicate bird assemblages in primary types of habitats (size of each circle corresponds
to the number of bird species). The red numbers near the circles indicate species richness. The black numbers near the
circles (01–10) indicate the codes of habitat types (see Fig. 4 and Table 2 for detail).
54SPITSYN ET AL. Figure 4. Relative bird species abundance (log10 scale) over habitat patches on Yuzhny Island of Novaya Zemlya. Numbers of (01) – (10) are the codes of the habitat types. Differences between assemblages were all significant (Kruskal-Wallis test: p = 0.003). Images show habitat types; numbers indicate their codes (see Table 2 for detail). (Photos: V. M. Spitsyn). Ecologica Montenegrina, 39, 2021, 46-58 55
BIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
Discussion
The results of our short-term summer survey expand the existing knowledge on bird assemblages of Arctic
Ocean archipelagoes. For instance, only avifaunal data was available for Novaya Zemlya, including the two
most recent annotated checklists of bird species (Spitsyn et al. 2018, 2020). Here, we report through our
results, the first quantitative analysis of bird assemblages on the Yuzhny Island of Novaya Zemlya. Our data
aligns with earlier observations (Tertitsky and Pokrovskaya 2011) revealing that Snow Bunting
(Plectrophenax nivalis) to be the most abundant species in arctic tundra of the Yuzhny Island. Tertitsky and
Pokrovskaya (2011) also listed a few additional species, which were abundant in several localities, such as
the Horned Lark (Eremophila alpestris), Bean Goose (Anser fabalis), Purple Sandpiper (Calidris maritima)
and Ringed Plover (Charadrius hiaticula). Based on our data, all these species were also common
representatives in certain assemblages in 2015. The most abundant bird species observed in sea habitats of
the Yuzhny Island were the Black-legged Kittiwake (Rissa tridactyla), Common Eider (Somateria
mollissima) and Thick-billed Murre (Uria lomvia). These results correspond with those reported in a fjord on
Svalbard, in which Black-legged Kittiwake, Common Eider, Black Guillemot (Cepphus grylle) and Little
Auk (Alle alle) were the most abundant taxa (Stempniewicz et al. 2017). Diemer et al. (2011) observed nine
sea bird species in the fjords of Baffin Island, Canada, and listed Common Eider (Somateria mollissima),
Glaucous Gull (Larus hyperboreus) and Black Guillemots (Cepphus grylle) as the most abundant species.
Terrestrial bird assemblages on the Yuzhny Island share a low species richness (4-13 species) and
simplified structure. These assemblages usually contained a single top predator and a few non-predatory
species. We found that carnivorous species such as Rough-legged Buzzard (Buteo lagopus) and Snowy Owl
(Bubo scandiacus) are clearly divided by habitat preferences (see Figs. 3A and 4). The Rough-legged
Buzzard was associated with mountain rocky areas with fragmented plant cover, in which Snow Bunting and
White Wagtail (Motacilla alba) were also present. In contrast, Snowy Owl preferred habitats with more
complete plant cover and higher richness of small and medium-sized birds. Although this owl species is
known to be a lemming specialist (Therrien et al. 2014), it can also prey on a variety of bird species, as was
recorded in Iceland (Stenkewitz and Nielsen 2019). The assemblages with maximum species richness (11-13
species) were associated with willow tundra habitats. The presence of willows greatly increased species
richness of bird assemblages in arctic, alpine, and boreal regions, such as in the Yellowstone National Park,
USA (Baril et al. 2009). However, true willow-nesting species are lacking on Novaya Zemlya and all the
passerines of this area could be considered as tundra-nesting species (Sammler et al. 2008).
Basic knowledge on terrestrial bird assemblages is restricted for almost all the Arctic Ocean
archipelagoes. The Snow Bunting was the most abundant species on Ellesmere Island (Freedman and
Svoboda 1982; Trefry et al. 2010) and Devon Island (Falconer et al. 2008) in the Canadian Arctic
Archipelago, which corresponds with our data for Novaya Zemlya. This species prevailed among passerine
birds on Svalbard as well (Jakubiec 1982; Wojczulanis-Jakubas et al. 2008). In contrast, Lapland Longspur
(Calcarius lapponicus) was the dominant land bird species on Banks Island, Somerset Island, and in several
regions of mainland Canada (Henry and Mico 2002; Latour et al. 2005; Andres 2006). Lepage et al. (1998)
also showed that both the Snow Bunting and the Lapland Longspur are common breeding birds on Bylot
Island and adjacent Baffin Island, Arctic Canada. The longspur selected nest sites with a greater amount of
shrubs (Boal and Andersen 2005), which might explain the differences in abundance of the two passerine
species throughout a plethora of Arctic islands.
Freshwater and coastal sea bird assemblages on Yuzhny Island represented an amalgam of aquatic
and sea bird species (see Fig. 3A). This pattern reflects the influence of insular environments, with multiple
ecotones between patches of various ecosystems. This pattern is typical for island assemblages of birds
(Jakubiec 1982; Wojczulanis-Jakubas et al. 2008) and other living organisms (Bolotov 2014).
Acknowledgements
We are grateful to Dr. Vladimir Tarasov (Yekaterinburg, Russia) and one anonymous reviewer for their
valuable comments on earlier versions of this paper. This study was partly supported by the Ministry of
Science and Higher Education of the Russian Federation (projects АААА-А17-117033010132-2 to V.M.S.
and АААА-А18-118012390161-9 to I.N.B.), and Russian Foundation for Basic Research (projects 18-44-
292001 to I.N.B. and 19-34-90012 to I.N.B. and V.M.S.).
56SPITSYN ET AL.
References
Andres, B. A. (2006) An Arctic-breeding bird survey on the northwestern Ungava Peninsula, Quebec,
Canada. Arctic, 59, 311–318. https://doi.org/10.14430/arctic316
Anker-Nilssen, T., Bakken, V., Strøm, H., Golovkin, A., Bianki, V. & Tatrinkova, I. (Eds.) (2000) The status
of marine birds breeding in the Barents Sea Region. Rapportserien 113, Norsk Polarinstitutt,
Tromsø, Norway, 213 рp.
Baril, L., Hansen, A., Renkin, R. & Lawrence, R. (2009) Willow-bird relationships on Yellowstone’s
northern range. Yellowstone Science, 17, 19–26.
Black, A. L., Gilchrist, H. G., Allard, K. A. & Mallory, M. L. (2012) Incidental observations of birds in the
vicinity of Hell Gate Polynya, Nunavut: species, timing, and diversity. Arctic, 65(2), 145–154.
https://doi.org/10.14430/arctic4196
Boal, C. W. & Andersen, D. E. (2005) Microhabitat characteristics of Lapland longspur, Calcarius
lapponicus, nests at Cape Churchill, Manitoba. The Canadian Field-Naturalist, 119(2), 208–213.
Bolotov, I. N. (2014) Pathways of formation of the fauna of the Solovetsky Archipelago, the White Sea,
Northwest Russia. Entomological Review, 94(4), 562–578.
https://doi.org/10.1134/S0013873814040095
Burnham, K. K., Sinnett, D. R., Johnson, J. A., Burnham, J. L., Baroch, J. A. & Konkel, B. W. (2014) New
species records and changes in abundance of waterfowl in northwest Greenland. Polar Biology,
37(9), 1289–1300. https://doi.org/10.1007/s00300-014-1520-z
Chapin, F. S., Sturm, M., Serreze, M. C., McFadden, J. P., Key, J. R., Lloyd, A. H., McGuire, A. D., Rupp,
T. S., Lynch, A. H., Schimel, J. P., Beringer, J., Chapman, W. L., Epstein, H. E., Euskirchen, E. S.,
Hinzman, L. D., Jia, G., Ping, C.-L., Tape, K. D., Thompson, C. D. C., Walker, D. A. & Welker, J.
M. (2005) Role of land-surface changes in Arctic summer warming. Science, 310(5748), 657–660.
https://doi.org/10.1126/science.1117368
Davey, C. M., Chamberlain, D. E., Newson, S. E., Noble, D. G. & Johnston, A. (2012) Rise of the
generalists: evidence for climate driven homogenization in avian communities. Global Ecology and
Biogeography, 21, 568–578. https://doi.org/10.1111/j.1466-8238.2011.00693.x
Descamps, S., Aars, J., Fuglei, E., Kovacs, K. M., Lydersen, C., Pavlova, O., Pedersen, Å. Ø., Ravolainen,
V. & Strøm, H. (2017) Climate change impacts on wildlife in a High Arctic archipelago – Svalbard,
Norway. Global Change Biology, 23(2), 490–502. https://doi.org/10.1111/gcb.13381
Dickson, D. L. & Gilchrist, H. G. (2002) Status of marine birds of the southeastern Beaufort Sea. Arctic,
55(S1), 46–58. https://doi.org/10.14430/arctic734
Diemer, K. M., Conroy, M. J., Ferguson, S. H., Hauser, D. D., Grgicak-Mannion, A. & Fisk, A. T. (2011)
Marine mammal and seabird summer distribution and abundance in the fjords of northeast
Cumberland Sound of Baffin Island, Nunavut, Canada. Polar Biology, 34(1), 41–48.
https://doi.org/10.1007/s00300-010-0857-1
Falconer, C. M., Mallory, M. L. & Nol, E. (2008) Breeding biology and provisioning of nestling snow
buntings in the Canadian High Arctic. Polar Biology, 31(4), 483–489.
https://doi.org/10.1007/s00300-007-0374-z
Freedman, B. & Svoboda, J. (1982) Populations of breeding birds at Alexandra Fjord, Ellesmere Island,
Northwest Territories, compared with other arctic localities. The Canadian Field-Naturalist, 96, 56–
60.
Gilg, O., Sané, R., Solovieva, D. V., Pozdnyakov, V. I., Sabard, B., Tsanos, D., Zöckler, C., Lappo, E. G.,
Syroechkovski, Jr, E. E. & Eichhorn, G. (2000) Birds and mammals of the Lena Delta nature reserve,
Siberia. Arctic, 53(2), 118–133. https://doi.org/10.14430/arctic842
Henry, J. D. & Mico, M. (2002) Relative abundance, habitat use, and breeding status of birds in Aulavik
National Park, Banks Island, Northwest Territories. The Canadian Field Naturalist, 116(3), 393–
407.
Howard, C., Stephens, P. A., Pearce‐Higgins, J. W., Gregory, R. D. & Willis, S. G. (2015) The drivers of
avian abundance: patterns in the relative importance of climate and land use. Global Ecology and
Biogeography, 24(11), 1249–1260. https://doi.org/10.1111/geb.12377
Jakubiec, Z. (1982) A quantitative investigation on birds of the Hornsund region, SW Spitsbergen. Acta
Universitatis Wratislaviensis, 525, 77–91.
Ecologica Montenegrina, 39, 2021, 46-58 57BIRD ASSEMBLAGES ON YUZHNY ISLAND OF NOVAYA ZEMLYA
Kalyakin, V. N. (1993) The fauna of birds and mammals in the Novaya Zemlya region and assessment of its
status [In Russian]. Novaya Zemlya, 2(3), 23–90.
Kardynal K.J., Bartzen B.A. & Latour P.B. (2015) Avian communities of the Northern Mackenzie
Mountains, Northwest Territories, Canada. Arctic, 68(3), 331–340.
https://doi.org/10.14430/arctic4508
Latour, P. B., Machtans, C. S. & Beyersbergen, G. W. (2005) Shorebird and passerine abundance and habitat
use at a High Arctic breeding site: Creswell Bay, Nunavut. Arctic, 58(1), 55–65.
https://doi.org/10.14430/arctic389
Lepage, D., Nettleship, D. N. & Reed, A. (1998) Birds of Bylot Island and adjacent Baffin Island, Northwest
Territories, Canada, 1979 to 1997. Arctic, 51(2), 125–141. https://doi.org/10.14430/arctic1054
Pearson, H. J. (1899) Beyond Petsora eastward: two summer voyages to Novaya Zemlya and the islands of
Barents Sea. R.H. Porter, London.
Pokrovskaya, I. V. & Tertitsky, G. M. (1993) The current status of commercial avifauna of Novaya Zemlya
[In Russian]. Novaya Zemlya, 2(3), 91–97.
Robinson, B. G., Franke, A. & Derocher, A. E. (2014) The influence of weather and lemmings on
spatiotemporal variation in the abundance of multiple avian guilds in the Arctic. PloS ONE, 9(7),
e101495. https://doi.org/10.1371/journal.pone.0101495
Rozenfeld, S. B. & Spitsyn, V. M. (2017) The results of reconnaissance ornithological observations within
the expedition «Arctic Floating University 2015» on the research ship «Professor Molchanov» [In
Russian]. Russian Journal of Ornithology, 26(1443), 1901–1909.
Sammler, J. E., Andersen, D. E. & Skagen, S. K. (2008) Population trends of tundra-nesting birds at Cape
Churchill, Manitoba, in relation to increasing goose populations. Condor, 110(2), 325–334.
https://doi.org/10.1525/cond.2008.8438
Šmilauer, P. & Lepš, J. (2014) Multivariate Analysis of Ecological Data using Canoco 5. Cambridge:
Cambridge University Press, 361 p.
Sokolov, V., Ehrich, D., Yoccoz, N. G., Sokolov, A. & Lecomte, N. (2012) Bird communities of the Arctic
shrub tundra of Yamal: habitat specialists and generalists. PLoS ONE, 7(12), e50335.
https://doi.org/10.1371/journal.pone.0050335
Spitsyn, V. M., Glazov, P. M., Anufriev, V. V. & Rozenfeld, S. B. (2020) An updated annotated list of birds
of the Novaya Zemlya Archipelago. Biharean Biologist, 14(2), 98–104.
Spitsyn, V. M., Rozenfeld, S. B. & Bolotov, N. I. (2018) Annotated list of bird species of the Malye
Karmakuly Polar Station, Yuzhny Island of Novaya Zemlya. Biharean Biologist, 12(1), 21–26.
Stempniewicz, L., Goc, M., Kidawa, D., Urbański, J., Hadwiczak, M. & Zwolicki, A. (2017) Marine birds
and mammals foraging in the rapidly deglaciating Arctic fjord-numbers, distribution and habitat
preferences. Climatic Change, 140, 533–548. https://doi.org/10.1007/s10584-016-1853-4
Stenkewitz, U. & Nielsen, Ó. K. (2019) The Summer Diet of the Snowy Owl (Bubo scandiacus) in Iceland.
Journal of Raptor Research, 53(1), 98–101. https://doi.org/10.3356/JRR-17-95
Strøm, H., Øien, I. J., Opheim, J., Kuznetsov, E. A. & Khakhin, G.V. (1994) Seabird censuses on Novaya
Zemlya 1994. Norwegian Ornithological Society Report, 2, 1–38.
Tertitsky, G. М. & Pokrovskaya, I. V. (2011) On avifauna and bird population of Novaya Zemlya [In
Russian]. The Russian Journal of Ornithology, 20(688), 1827–1836.
Therrien, J. F., Gauthier, G., Pinaud, D. & Bêty, J. (2014) Irruptive movements and breeding dispersal of
snowy owls: a specialized predator exploiting a pulsed resource. Journal of Avian Biology, 45(6),
536–544. https://doi.org/10.1111/jav.00426
Thomas, C. D. & Lennon, J. J. (1999) Birds extend their ranges northwards. Nature, 399(6733), 213.
https://doi.org/10.1038/20335
Trefry, S. A., Freedman, B., Hudson, J. M. G. & Henry, G. H. R. (2010) Breeding bird surveys at Alexandra
Fjord, Ellesmere Island, Nunavut (1980–2008). Arctic, 63(3), 308–314.
https://doi.org/10.14430/arctic1494
Walker, D. A. (2000) Hierarchical subdivision of Arctic tundra based on vegetation response to climate,
parent material and topography. Global Change Biology, 6(S1), 19–34.
https://doi.org/10.1046/j.1365-2486.2000.06010.x
Wojczulanis-Jakubas, K., Jakubas, D. J. & Stempniewicz, L. (2008) Avifauna of Hornsund area, SW
Spitsbergen: present state and recent changes. Polish Polar Research, 29(2), 187–197.
58You can also read

















































