Flight-Fecundity Trade-offs: A Possible Mechanistic Link in Plant-Herbivore-Pollinator Systems
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
PERSPECTIVE
published: 25 April 2022
doi: 10.3389/fpls.2022.843506
Flight-Fecundity Trade-offs: A
Possible Mechanistic Link in
Plant–Herbivore–Pollinator Systems
Goggy Davidowitz 1*, Judith L. Bronstein 2 and Natasha Tigreros 1
1
Department of Entomology, University of Arizona, Tucson, AZ, United States, 2 Department of Ecology and Evolutionary
Biology, University of Arizona, Tucson, AZ, United States
Plant–herbivore and plant–pollinator interactions are both well-studied, but largely
independent of each other. It has become increasingly recognized, however, that pollination
and herbivory interact extensively in nature, with consequences for plant fitness. Here,
we explore the idea that trade-offs in investment in insect flight and reproduction may
be a mechanistic link between pollination and herbivory. We first provide a general
background on trade-offs between flight and fecundity in insects. We then focus on
Lepidoptera; larvae are generally herbivores while most adults are pollinators, making
them ideal to study these links. Increased allocation of resources to flight, we argue,
Edited by:
Anne Muola, potentially increases a Lepidopteran insect pollinator’s efficiency, resulting in higher plant
University of Turku, fitness. In contrast, allocation of resources to reproduction in the same insect species
Finland
reduces plant fitness, because it leads to an increase in herbivore population size.
Reviewed by:
Zong-Xin Ren, We examine the sequence of resource pools available to herbivorous Lepidopteran larvae
Kunming Institute of Botany (CAS), (maternally provided nutrients to the eggs, as well as leaf tissue), and to adults (nectar
China
and nuptial gifts provided by the males to the females), which potentially are pollinators.
Quint Rusman,
University of Zurich, Last, we discuss how subsequent acquisition and allocation of resources from these
Switzerland pools may alter flight–fecundity trade-offs, with concomitant effects both on pollinator
*Correspondence: performance and the performance of larval herbivores in the next generation. Allocation
Goggy Davidowitz
goggy@arizona.edu decisions at different times during ontogeny translate into costs of herbivory and/or benefits
of pollination for plants, mechanistically linking herbivory and pollination.
Specialty section:
This article was submitted to Keywords: pollination, herbivory, nutrient tradeoffs, Lepidoptera, nuptial gift, nectar
Functional Plant Ecology,
a section of the journal
Frontiers in Plant Science
INTRODUCTION
Received: 26 December 2021
Accepted: 16 March 2022 Plant–herbivore and plant–pollinator interactions are both well-established, but largely independent
Published: 25 April 2022 fields of study. Pollination is a mutually beneficial interaction and historically has been the
Citation: most thoroughly studied of all mutualisms (Bronstein, 1994). The key issue in the study of
Davidowitz G, Bronstein JL and pollination is how plants obtain and donate high-quality pollen to maximize reproductive
Tigreros N (2022) Flight-Fecundity
output. In the case of the over 85% of plant species that are animal-pollinated (Ollerton et al.,
Trade-offs: A Possible Mechanistic
Link in Plant–Herbivore–Pollinator
2011), this involves attracting and rewarding partners that will transfer pollen among flowers
Systems. of the same species. Herbivory, in contrast, is an antagonistic interaction between plants and
Front. Plant Sci. 13:843506. animals. In some cases, consumption of leaves can dramatically reduce plant growth and
doi: 10.3389/fpls.2022.843506 survival (Lehndal and Ågren, 2015). Key issues in the study of herbivory have been how
Frontiers in Plant Science | www.frontiersin.org 1 April 2022 | Volume 13 | Article 843506Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory
plants defend themselves against being eaten, and when and may mitigate damage (Davidowitz et al., 2003; Wilson et al.,
how herbivores are able to circumvent these defenses (Núñez- 2019), among other factors.
Farfán et al., 2007). Larval Lepidoptera are predominantly herbivores and most
In recent years, it has become increasingly well-recognized adults are pollinators (Hahn and Brühl, 2016), often of the
that pollination and herbivory are not, as might be suggested same plant species (Altermatt and Pearse, 2011), making them
by these contrasting concerns, independent of each other ideal to address this link between herbivory and pollination.
(Rusman et al., 2019). Rather, they interact in ways that We note that this linkage exists whether the pollinator lays
synergistically contribute to a plant’s reproductive success eggs on the same plant or on different individual plants of
(Marquis, 1992; Bronstein et al., 2007; Jacobsen and Raguso, the same species and whether the plant being eaten and the
2018; Haas and Lortie, 2020; Johnson et al., 2021). The presence plant being pollinated are different species, which may result
of herbivore damage, for instance, can reduce the likelihood in differential costs and benefits of herbivory and pollination,
that pollinators will be attracted to flowers; it can also reduce respectively.
resources necessary to produce flowers, seeds, and fruits. Here, we associate resource allocation to flight with increased
Herbivores may also simply consume the flowers. In all of pollination efficiency and allocation to fecundity with herbivory
these cases, herbivory reduces plant fitness through reduced damage. In addition to nectar foraging and pollen transfer,
effectiveness of pollination. In other situations, however, the flight is of course also used for other functions, such as to
presence of herbivores actually enhances pollination. This occurs, find mates and host plants (Chai and Srygley, 1990; Willis
for example, when a single species is both the pollinator and and Arbas, 1991; Mitra et al., 2016). However, because nectar
herbivore of the same plant species. In these cases, the probability foraging is the most relevant function of flight to a plant’s
of pollination and herbivory increase together. The best-known fitness due to its resultant pollination, we focus on the nectar
examples are highly specialized insects, such as fig wasps and foraging function of flight.
yucca moths, that pollinate plants, then lay eggs in the flowers, The efficiency of an animal as a pollinator entails more
with the pollinator’s offspring subsequently destroying a portion than just flight. It encompasses numerous pollination-related
of the developing seeds (Kato and Kawakita, 2017). More traits including multimodal signaling, used by the pollinator
common, but not as well-studied, are cases in which insects to find the flower (Raguso and Willis, 2002), the reliability of
feed on floral nectar, then lay eggs on the leaves of the same the signal used by the plant to attract the pollinator (Von
individual plant or on neighboring plants of the same species; Arx et al., 2012), proboscis length matching with nectar tube
the pollinator’s offspring in this case are folivores of their length (Haverkamp et al., 2016; Soteras et al., 2020), flower
host plant. The best-known of these herbivorous pollinators handling time (Kunte, 2007; Riffell and Alarcón, 2013), pollen
are Lepidoptera, including but not restricted to those with transport distances (Herrera, 1987), and floral constancy (Goulson
narrow diet breadths (Bronstein et al., 2009; Altermatt and et al., 1997). We focus on allocation to flight (flight muscles
Pearse, 2011). and wings), as this is the largest resource sink related to
Recent conceptual advances linking herbivory and pollination pollination (G. Davidowitz, unpublished data).
have largely adopted a plant perspective (e.g., Lucas-Barbosa, Below, we first provide a general background on trade-offs
2016; Jacobsen and Raguso, 2018; Kessler and Chautá, 2020). between flight and fecundity in insects. We then examine the
In this perspective, we develop a framework that links herbivory sequence of resource pools available to Lepidopteran herbivores
and pollination from the animal perspective instead. Specifically, and pollinators. Finally, we discuss how subsequent acquisition
we explore the idea that trade-offs between investment into and allocation of resources from these pools may alter the
flight vs. fecundity functionally link insect pollination and flight–fecundity trade-off, with concomitant effects both on
herbivory. Flight–fecundity trade-offs in insects are a well- pollinator performance and the performance of larval herbivores
studied phenomenon (Johnson, 1963; Roff, 1986, 1990, 1994; in the next generation.
Rankin and Burchsted, 1992; Dingle, 1996; Zera et al., 1999;
Zera and Brink, 2000; Zera and Larsen, 2001; Gu et al., 2006;
Hanski et al., 2006; Karlsson and Johansson, 2008; Guerra and FLIGHT–FECUNDITY TRADE-OFFS
Pollack, 2009; Tigreros and Davidowitz, 2019). At a basic level,
allocation of resources to flight will modify an insect pollinator’s In insects, allocation to flight begins with an allocation to
efficiency, with a resultant increase in plant fitness. In contrast, flight muscle and wings: larger flight muscles increase power
allocation of resources to fecundity leads to an increase in output and larger wings reduce wing loading, both of which
the herbivore population size produced in the next generation. increase flight performance (Dudley, 2002). In general, resource
Increased allocation of resources to fecundity may or may allocation to flight is essential as it allows the adult to find
not translate linearly into herbivore damage as damage may mates, disperse, and forage for additional resources. In insect
differ among populations (Marquis, 1992), the strength of pollinators in particular, the dimensions of flight muscle and
selection induced by the herbivore can differ (Agrawal et al., wings can have significant effects on pollinator flight (Dudley,
2012), tolerance vs. resistance to herbivores may mitigate damage 2002), affecting, for example, the ability to forage for nectar
(McCall et al., 2020), when during ontogeny herbivory occurs from flowers buffeted by the wind while hovering (Hedrick
effects overall damage (Boege and Marquis, 2005) and the and Daniel, 2006; Sprayberry and Daniel, 2007). Subsequent
quality of the host plant and its effect on herbivore growth investments are needed to fuel flight itself, which is the most
Frontiers in Plant Science | www.frontiersin.org 2 April 2022 | Volume 13 | Article 843506Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory
energetically expensive mode of locomotion known (McCallum of resources (King et al., 2011; Descamps et al., 2016). Increased
et al., 2013). In insects, flight can be 30-fold more costly than allocation to locomotion, for example, can improve an organism’s
terrestrial locomotion (Harrison and Roberts, 2000). Insects ability to forage and acquire additional resources. The quantity
that act as pollinators often hover while feeding on nectar, a and quality of resources that a juvenile herbivore acquires can
behavior that is energetically demanding (Biewener and Patek, modify its nectar preferences as an adult (Mevi-Schütz and Erhardt,
2018). For example, hovering hawkmoths require 170 times 2003); this in turn may influence its effectiveness as a pollinator.
more energy than basal metabolism (Bartholomew and Casey, We distinguish between plant-derived resources (foliage and
1978). The energy from nectar available to the insect nectar) and insect-derived resources (maternally provided provisions
differs across plant species and may differ among plant to the egg, and nuptial gifts that males provide to females during
populations and communities as well (Nicolson et al., 2007; copulation). These resources are available at different times during
Lebeau et al., 2016). an insect’s ontogeny (Figure 1) and differ in their relative amounts
The nectar load itself can affect the stability and of proteins, carbohydrates, and lipids (see below). These resource
maneuverability of the insect in flight, with potential effects pools can have significant consequences for the growth of the
on feeding efficiency (Mountcastle et al., 2015). Feeding efficiency, herbivorous juvenile and the pollinating adult, with potential
in turn, may translate into pollinator effectiveness (Goulson, fitness consequences to the plant. Below, we examine each of
1999). Flight distance is an important component of pollinator these resource pools in the order they are available to the insect.
efficiency as it may affect the pollen dispersal ability of the
insect pollinator (Schulke and Waser, 2001; Pasquet et al., 2008).
Allocation to reproduction involves investments into the RESOURCE ACQUISITION AND
reproductive system as well as to eggs. Larval diet can affect
ALLOCATION IN HERBIVOROUS
the number of ovarioles in the ovary, and hence the maximum
number of eggs that can be laid; fecundity is reduced on poor JUVENILES
quality larval diets due to fewer ovarioles (Sisodia and Singh,
2012; Aguila et al., 2013). In all insects, reproductive output Maternally Provisioned Resources
The first resource pool to which herbivorous insects have access
is determined by the availability of nutritional resources, whether
is provided by mothers, through the nutritional resources they
acquired during the larval or the adult stages (Wheeler, 1996;
deposit into eggs (Roach and Wulff, 1987; Bernardo, 1996;
Papaj, 2000; Awmack and Leather, 2002). This is discussed in
Fox and Czesak, 2000). In contrast to the leaf tissue that will
depth, below.
be consumed once the insect emerges from the egg (see below),
Investments in flight and fecundity trade off (two words)
nutrients in eggs include substantial amounts of proteins
because both require the same macronutrient resources, proteins,
(~40%–50%) and lipids (30%–40%). As a consequence, maternal
carbohydrates, and lipids, all of which are often in limited
egg provisioning of nutritional resources can have profound
supply (Baker and Baker, 1986; van Noordwijk and de Jong,
effects on offspring development and subsequent life-history
1986; Stearns, 1989; Zera and Harshman, 2001; Boggs, 2009;
traits (Mousseau and Dingle, 1991; Bernardo, 1996; Mousseau
Saeki et al., 2014; Tigreros and Davidowitz, 2019). Although
and Fox, 1998; Fox and Czesak, 2000; Hunt and Simmons,
other limiting resources, such as time available to devote to
2000). This in turn can influence flight–fecundity trade-offs
life-history activities, can also trade-off, nutrient-based trade-
once the offspring eclose as adults. At the same time, females
offs are probably the dominant type of trade-off in nature
experiencing flight–fecundity trade-offs may adjust the number
(Zera and Harshman, 2001; Boggs, 2009; Agrawal, 2020).
of eggs they produce as well as the quantity of nutrients
Tigreros and Davidowitz (2019) showed that in wing
provisioned to each egg (Tigreros and Davidowitz, 2019).
monomorphic insect species, 76% of studies showed a flight–
Females of the Speckled Wood butterfly, Pararge aegeria, that
fecundity trade-off when resource availability was manipulated.
are forced to fly long distances, for example, produce smaller
The more resources allocated to flight, the fewer resources
eggs and smaller offspring that take longer to develop (Gibbs
that are available for fecundity (and vice versa), resulting in
et al., 2010). Similarly, females experiencing poor nutritional
a negative association between flight and fecundity. As a
consequence, we can predict a negative association between environments during either the larval or adult stage generally
the role of an insect as an herbivore and that as a pollinator decrease the nutrients they put into eggs (Bernardo, 1996;
(see above). With this introduction to nutrient-based trade-offs Mevi-Schütz and Erhardt, 2005; Geister et al., 2008). In other
between flight and fecundity, we next examine the sequence cases, however, Lepidoptera may increase nutrient investment
of nutrient pools available to Lepidoptera. in eggs to improve offspring performance on low-quality host
plants (Rotem et al., 2003). As a consequence, the provisioned
egg itself may provide a link between the maternal and offspring
resource acquisition and allocation strategies, as well as associated
THE SEQUENCE OF RESOURCE POOLS life-history trade-offs (Figure 1).
The timing of the acquisition and allocation of nutrients can
influence acquisition of additional resources (Figure 1). Some Leaf Tissue
empirical studies suggest that allocation to traits related to acquisition The larvae of most Lepidoptera feed on green plant tissues.
ability, such as flight, may directly influence the further acquisition These tissues contain large amounts of carbohydrates, but only
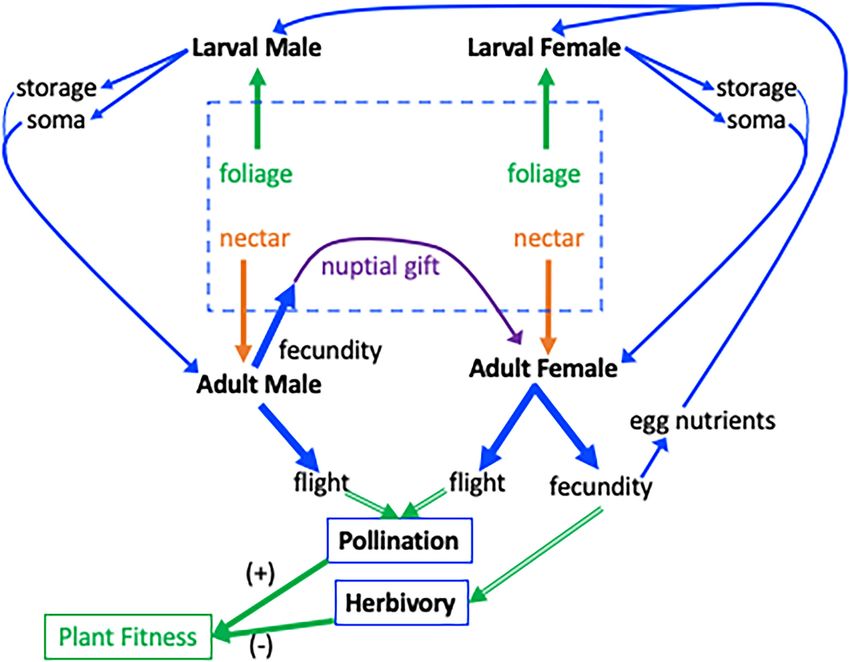
Frontiers in Plant Science | www.frontiersin.org 3 April 2022 | Volume 13 | Article 843506Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory FIGURE 1 | Interaction between a plant and a Lepidopteran that is an herbivore as a larva and a pollinator as an adult. The central dashed box indicates resource pools to the insect. Host-plant foliage is the resource for larvae (green arrows from dashed box), nectar is a resource for adults (orange arrows), and nuptial gifts are a resource given to the female by the male (purple arrow). For simplicity, only resources relevant to flight–fecundity trade-offs are shown and allocation to other functions such as maintenance, are omitted. Blue lines indicate resources and green lines indicate effects on plant fitness. Larvae consume foliage for nutrient storage and growth (soma; strait blue arrows at top) which are available as resource pools in the adult following metamorphosis (curved blue arrows). Adult Lepidoptera can allocate resources to flight or fecundity (thick blue arrows). The consequences of flight–fecundity allocation decisions to the plant (double-lined green arrows) through herbivory and pollination are indicated by the thick green arrows. Allocation of resources to fecundity by males and females reduces plant fitness, green arrow (−), via herbivory. Allocation of resources to flight increases plant fitness, (+) green arrow, through pollination. Eggs produced by male allocation to nuptial gifts, and female allocation to fecundity, produce the next generation of herbivores (rightmost blue arrow). a small fraction of the lipids and protein (nitrogen) that a After emerging, but before finding a nectar source, adults must larva needs. While some of the dietary carbohydrates are maintain their bodies and fuel flight solely with larval stores. converted into lipids (Arrese and Soulages, 2010), the limited These endogenous reserves can be used, together with adult availability of dietary protein leads to a fundamental nutritional feeding, to produce eggs and fuel flight (Figure 1). mismatch between Lepidoptera (as well as other herbivores) Two contrasting scenarios of allocation of nutrients from and their host plants (Slansky, 1978; Mattson, 1980; Wilson leaf tissue can be envisioned. First, when juvenile resources et al., 2019). For example, host plants of the cabbage butterfly are limited, due either to low abundance or to low nutritional (Pieris rapae) contain only 1.9%–5.9% N (~9.4%–36.9% protein), value of the host plant, fewer resources will be available to compared to about 13% N content in the adult bodies at eclosion “build” the adult. In one scenario, we hypothesize that fewer (Morehouse and Rutowski, 2010). To make up such differences, resources are allocated to flight but allocation to fecundity is insects engage in compensatory feeding, eating more of nutrient- maintained, resulting in reduced efficiency of the adults during poor diets to reach their nutritional requirements (Simpson the feeding stage (when pollination occurs), while maintaining and Simpson, 1990; Nestel et al., 2016). This nutritional mismatch a high level of offspring herbivory. A net reduction in plant in the larval stage often contributes to flight–fecundity trade- fitness might result. Alternatively, in a second scenario, offs in Lepidoptera, because limited nutritional resources from we hypothesize that reduced nutrients available for juvenile leaf tissue are differentially allocated to flight (wings and flight herbivores may result, in the adult stage, in reduced allocation muscle) vs. reproductive (ovaries and eggs) structures of the of resources to fecundity but not to flight. In this case, pollination adult (Tigreros and Davidowitz, 2019). Furthermore, some of efficiency may remain high and herbivore populations may the resources acquired from the larval diet are stored and be smaller in the next generation, with net fitness benefits to carried over through metamorphosis (Arrese and Soulages, 2010). the plant. Frontiers in Plant Science | www.frontiersin.org 4 April 2022 | Volume 13 | Article 843506
Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory
RESOURCE ACQUISITION AND both sperm and additional nutrients. These nutrients can be used
ALLOCATION IN POLLINATING ADULTS by the female in oogenesis and somatic maintenance (Boggs,
1990, 1997; Karlsson, 1998). In contrast to leaf tissue and
Floral Nectar nectar, nuptial gifts contain substantial amounts of protein.
Nutrient deficiencies in the larval stage, which, can lead to For example, nuptial gifts in Pierid butterflies contain as much
flight–fecundity trade-offs, might be compensated for by the as 50% protein (Bissoondath and Wiklund, 1996; Karlsson,
subsequent acquisition and allocation of nectar nutrients 1998; Tigreros, 2013) with a large percent of that being essential
(Figure 1). A growing number of studies indicate that nectar amino acids: for example, ~35% (Meslin et al., 2017). While
can be as important as larval-derived reserves in supporting providing an additional source of macronutrients for adult
both flight and fecundity in adult females. Throughout their females, nuptial gifts have the potential to both ameliorate
adult lives, moths and butterflies typically feed on floral nectars, and magnify flight–fecundity trade-offs. In Pierids, a single
which are carbohydrate-rich solutions (20%–50% sugars) enriched nuptial gift can provide the necessary nutrients to produce
by small amounts of essential and non-essential amino acids 50–80 eggs, a substantial contribution to female fecundity
(Baker and Baker, 1986; Lanza et al., 1995; Nicolson and (Karlsson, 1998; Wiklund et al., 1998; Wedell and Karlsson,
Thornburg, 2007; Willmer, 2011). In general, females that feed 2003). Amino acids supplied through nuptial gifts can change
on nectar produce more eggs than females that do not (Sasaki female reliance on amino acid-rich nectar preference (Mevi-
and Riddiford, 1984; von Arx et al., 2013). There are at least Schütz and Erhardt, 2003), which may affect the pollination
two explanations for this. First, carbohydrates from nectar efficiency of the female. At the same time, because a nuptial
provide the energy necessary to fuel flight (O’Brien, 1999), gift is more than 80% water (Boggs and Watt, 1981), an
and contribute to the synthesis of non-essential amino acids important resource in arid environments (Contreras et al.,
for egg production (O’Brien et al., 2002, 2004). Second, contrary 2013), female acquisition of nuptial gifts can increase the cost
to the paradigm that essential amino acids can only be drawn of flight by increasing wing loading. For example, a fresh
from the larval diet (O’Brien et al., 2002), some studies have spermatophore in P. rapae may add up to 10% of the female
shown that nectar-derived essential amino acids enhance eclosion mass (Tigreros, unpublished).
fecundity in Lepidoptera (Mevi-Schütz and Erhardt, 2005; Levin Males may rely on both larval- and adult-derived resources
et al., 2017b), especially when resources acquired by the larvae to produce nuptial gifts. For example, nitrogen content in larval
are limited (Mevi-Schütz and Erhardt, 2005). diets can change the composition of nuptial gifts (Bonoan
Resources acquired by male and female adult Lepidoptera et al., 2015), and nectar uptake by males can increase the size
(and other nectar-feeders) are not necessarily identical. In a of the nuptial gift by adding more nutrients than those derived
comprehensive literature review, Smith et al. (2019) showed from the larva diet (Watanabe and Hirota, 1999; Levin et al.,
that male and female pollinators differ in the species of flowers 2016). Nuptial gifts can be costly to produce, representing up
visited, as well as in their visitation frequencies. Female to 15% of the male body weight in Lepidoptera (Svärd and
pollinators tend to visit a higher diversity of flowers than Wiklund, 1989). As a consequence, males of species with
males, whereas males tend to forage over greater distances substantial nuptial gift donation may prefer to mate with
than females. These differences can potentially result in (Rutowski, 1985; Tigreros et al., 2014), and transfer more
differences between conspecific males and females in their nutrients to females that are more fecund (Bonoan et al., 2015).
quality as pollinators (Smith et al., 2019). Once nectar has In this case, a female’s ability to acquire nutrients from this
been ingested, how it is subsequently invested into life-history resource pool (Tigreros, 2013; Tigreros et al., 2014; Bonoan
functions can also differ between sexes: females metabolize et al., 2015) would depend on how she had previously allocated
nectar-derived amino acids before utilizing larval-derived amino resources to flight and fecundity (Figure 1).
acids, whereas males preferentially use amino acids from larval
stores before using those derived from nectar (Levin et al.,
THE EFFECTS OF SEQUENTIAL
2017a). Males also allocate more nectar-derived amino acids
to flight muscles than do females (Levin et al., 2017a). Finally, ACQUISITION AND ALLOCATION OF
there are sex-related differences in how essential (EAA) and RESOURCES ON PLANT FITNESS
non-essential amino acids (NEAA) are allocated: after feeding,
males metabolize EAAs more readily than females, whereas The acquisition of resources has typically been considered as
females preferentially allocate EAAs to reproduction (Levin a single event (the stem of the “Y” model, sensu van Noordwijk
et al., 2017a). and de Jong, 1986). In most systems, however, resource acquisition
and decisions governing resource allocation are not fixed, but
rather dynamic processes that change continually across an
Male Nuptial Gifts organism’s life (Zera and Harshman, 2001; Boggs, 2009; Kooijman,
Adult females can acquire nutrients from nuptial gifts, not 2009; Figure 1). Acquisition of additional resources is predicted
only from nectar. These nutritional gifts are a type of reproductive to reduce or mask potential trade-offs (Kaitala, 1987; Chippindale
investment that is widespread across animal taxa (Vahed, 1998; et al., 1993; Nijhout and Emlen, 1998; Zera and Harshman,
Lewis and South, 2012; Boggs, 2018). In insects, males transfer 2001; Harshman and Zera, 2007). This suggests that organisms
a structure called a spermatophore during mating, which includes may have a means to modulate (and even ameliorate) the
Frontiers in Plant Science | www.frontiersin.org 5 April 2022 | Volume 13 | Article 843506Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory
expression of a trade-off when acquiring resources from additional the latter become available. In many cases the adults do
pools, with implications for plant fitness. For example, females not feed at all (e.g., fig wasps and yucca moths; Kato and
of the Map butterfly, Araschnia levana, raised on low-quality Kawakita, 2017). Do the flight–fecundity trade-offs discussed
larval diets prefer nectar with amino acids, whereas females here illuminate these interactions as well?
raised on high-quality diets do not (Mevi-Schütz and Erhardt, 4. In arid environments, water is another critical resource that
2003). These nectar amino acids can enhance butterfly fecundity adult insects gain from feeding on nectar (Contreras et al.,
thereby increasing damage by the offspring herbivores (Mevi- 2013). Does this additional resource alters in any way the
Schütz and Erhardt, 2005). Thus, the sequential acquisition of resource allocation trade-offs between flight and fecundity
resources may change their allocation to flight or to fecundity we discuss here?
over time. 5. Does plant density-dependence affect how the flight–fecundity
Therefore, we may also expect the strength of the trade-off trade-off affects plant fitness? More specifically, does the
between flight and fecundity to change as the nutritional needs flight–fecundity trade-off differentially affect pollination when
and nutrient availability change across an organism’s life cycle the pollinator has numerous, vs. few, plants available at
(Figure 1). For example, an herbivore feeding on a nutritionally which it can feed, and how does the flight–fecundity trade-off
poor host plant might allocate more resources to flight at the affect herbivory when the female can lay eggs on numerous
expense of fecundity, with the potential fitness benefit to the versus few possible host plants?
plant. If, however, the emerged adult has access to an abundance
of nutrient-rich nectar, it may shift these resources to increased These, and additional, yet to be identified questions, make
fecundity (Sasaki and Riddiford, 1984; Levin et al., 2016, 2017a), flight–fecundity trade-offs an exciting area of future research
thereby obviating the flight–fecundity trade-off imposed by into the mechanistic link between pollination and herbivory,
larval resources. In another example, resources already allocated and plant–insect interactions more broadly.
to flight may be reallocated to reproduction following flight
muscle histolysis in aging butterflies (Jervis et al., 2005; Stjernholm
et al., 2005), with a resultant increase in herbivory costs to DATA AVAILABILITY STATEMENT
the plant.
The original contributions presented in the study are included
in the article/supplementary material; further inquiries can
FUTURE DIRECTIONS be directed to the corresponding author.
In this perspective, we have argued that trade-offs in resource
allocation between flight and fecundity in insects can provide AUTHOR CONTRIBUTIONS
a mechanistic link between pollination and herbivory with
subsequent effects on plant fitness. To further develop this GD, NT, and JB developed the ideas for the manuscript and
idea, we provide additional questions for future research. all authors were involved in the writing and editing. All authors
contributed to the article and approved the submitted version.
1. Here, we have focused on Lepidoptera. Do flight fecundity
trade-offs in other insect pollinator taxa, such as solitary
bees, flies, and beetles, have similar effects on plant fitness? FUNDING
2. We have argued that flight–fecundity trade-offs should have
a direct impact on plant reproduction. It will be exciting This work was supported by National Science Foundation
to explore, via models and empirical studies, how flight– (NSF-USA) grant IOS-2122282 to GD and NT.
fecundity trade-offs influence plant population dynamics and
evolution. Do different strengths of these trade-offs translate
to different effects on the plants? ACKNOWLEDGMENTS
3. We have focused on insects that feed on leaves as juveniles
and on nectar as adults. However, some specialized insect We would like to thank Sarah Britton, Noah DeFino, and
pollinators feed on seeds in the juvenile stage; still others Austin Cruz and two reviewers for comments on earlier versions
shift from feeding on leaves to feeding on flowers when of the manuscript and NSF-IOS for funding this work.
in plant populations. Science 338, 113–116. doi: 10.1126/science.
REFERENCES 1225977
Aguila, J. R., Hoshizaki, D. K., and Gibbs, A. G. (2013). Contribution of larval
Agrawal, A. A. (2020). A scale-dependent framework for trade-offs, syndromes, nutrition to adult reproduction in Drosophila melanogaster. J. Exp. Biol.
and specialization in organismal biology. Ecology 101:e02924. doi: 10.1002/ 216, 399–406. doi: 10.1242/jeb.078311
ecy.2924 Altermatt, F., and Pearse, I. S. (2011). Similarity and specialization of the larval
Agrawal, A. A., Hastings, A. P., Johnson, M. T., Maron, J. L., and Salminen, J. P. versus adult diet of European butterflies and moths. Am. Nat. 178, 372–382.
(2012). Insect herbivores drive real-time ecological and evolutionary change doi: 10.1086/661248
Frontiers in Plant Science | www.frontiersin.org 6 April 2022 | Volume 13 | Article 843506Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory Arrese, E. L., and Soulages, J. L. (2010). Insect fat body: energy, metabolism, Dingle, H. (1996). Migration: The Biology of Life on the Move. New York: and regulation. Annu. Rev. Entomol. 55, 207–225. doi: 10.1146/annurev- Oxford University Press. ento-112408-085356 Dudley, R. (2002). The Biomechanics of Insect Flight: Form, Function, Evolution. Awmack, C. S., and Leather, S. R. (2002). Host plant quality and fecundity Princeton, New Jersey: Princeton University Press. in herbivorous insects. Annu. Rev. Entomol. 47, 817–844. doi: 10.1146/ Fox, C. W., and Czesak, M. E. (2000). Evolutionary ecology of progeny size annurev.ento.47.091201.145300 in arthropods. Annu. Rev. Entomol. 45, 341–369. doi: 10.1146/annurev. Baker, H. G., and Baker, I. (1986). The occurrence and significance of amino ento.45.1.341 acids in floral nectar. Plant Syst. Evol. 151, 175–186. doi: 10.1007/BF02430273 Geister, T. L., Lorenz, M. W., Hoffmann, K. H., and Fischer, K. (2008). Adult Bartholomew, G. A., and Casey, T. M. (1978). Oxygen consumption of moths nutrition and butterfly fitness: effects of diet quality on reproductive output, during rest, preflight warm-up, and flight in relation to body size and wing egg composition, and egg hatching success. Front. Zool. 5:10. doi: 10.1186/1742- morphology. J. Exp. Biol. 76, 11–25. doi: 10.1242/jeb.76.1.11 9994-5-10 Bernardo, J. (1996). The particular maternal effect of propagule size, especially Gibbs, M., Breuker, C. J., and Van Dyck, H. (2010). Flight during oviposition egg size: patterns, models, quality of evidence and interpretations. Am. Zool. reduces maternal egg provisioning and influences offspring development in 36, 216–236. doi: 10.1093/icb/36.2.216 Pararge aegeria (L.). Physiol. Entomol. 35, 29–39. doi: 10.1111/j.1365-3032. Biewener, A., and Patek, S. (2018). Animal Locomotion. Oxford, UK: Oxford 2009.00706.x University Press. Goulson, D. (1999). Foraging strategies of insects for gathering nectar and Bissoondath, C. J., and Wiklund, C. (1996). Effect of male mating history and pollen, and implications for plant ecology and evolution. Perspectives in body size on ejaculate size and quality in two polyandrous butterflies, Pieris plant ecology, evolution and systematics 2, 185–209. napi and Pieris rapae (Lepidoptera: Pieridae). Funct. Ecol. 10, 457–464. doi: Goulson, D., Ollerton, J., and Sluman, C. (1997). Foraging strategies in the 10.2307/2389938 small skipper butterfly, Thymelicus flavus: when to switch? Anim. Behav. Boege, K., and Marquis, R. J. (2005). Facing herbivory as you grow up: the 53, 1009–1016. doi: 10.1006/anbe.1996.0390 ontogeny of resistance in plants. Trends Ecol. Evol. 20, 441–448. doi: 10.1016/j. Gu, H., Hughes, J., and Dorn, S. (2006). Trade-off between mobility and fitness tree.2005.05.001 in Cydia pomonella L. (Lepidoptera: Tortricidae). Ecol. Entomol. 31, 68–74. Boggs, C. L. (1990). A general model of the role of male-donated nutrients doi: 10.1111/j.0307-6946.2006.00761.x in female insects’ reproduction. Am. Nat. 136, 598–617. doi: 10.1086/ Guerra, P. A., and Pollack, G. S. (2009). Flight behaviour attenuates the trade- 285118 off between flight capability and in a wing polymorphic cricket. Biol. Lett. Boggs, C. L. (1997). Dynamics of reproductive allocation from juvenile and 5, 229–231. doi: 10.1098/rsbl.2008.0570 adult feeding: radiotracer studies. Ecology 78, 192–202. doi: 10.1890/0012- Haas, S. M., and Lortie, C. J. (2020). A systematic review of the direct and 9658(1997)078[0192:DORAFJ]2.0.CO;2 indirect effects of herbivory on plant reproduction mediated by pollination. Boggs, C. L. (2009). Understanding insect life histories and senescence through PeerJ 8:e9049. doi: 10.7717/peerj.9049 a resource allocation lens. Funct. Ecol. 23, 27–37. doi: 10.1111/j.1365-2435. Hahn, M., and Brühl, C. A. (2016). The secret pollinators: an overview of 2009.01527.x moth pollination with a focus on Europe and North America. Arthropod Boggs, C. L. (2018). “Male nuptial gifts: phenotypic consequences and evolutionary Plant Interact. 10, 21–28. doi: 10.1007/s11829-016-9414-3 implications,” Insect reproduction, eds. S. R. Leather and J. Hardie (CRC Hanski, I., Saastamoinen, M., and Ovaskainen, O. (2006). Dispersal-related Press)27–37. life-history trade-offs in a butterfly metapopulation. J. Anim. Ecol. 75, 91–100. Boggs, C. L., and Watt, W. B. (1981). Population structure of pierid butterflies doi: 10.1111/j.1365-2656.2005.01024.x IV. Genetic and physiological investment in offspring by male Colias. Oecologia Harrison, J. F., and Roberts, S. P. (2000). Flight respiration and energetics. 50, 320–324. doi: 10.1007/BF00344970 Annu. Rev. Physiol. 62, 179–205. doi: 10.1146/annurev.physiol.62.1.179 Bonoan, R. E., Al-Wathiqui, N., and Lewis, S. (2015). Linking larval nutrition Harshman, L. G., and Zera, A. J. (2007). The cost of reproduction: the to adult reproductive traits in the European corn borer Ostrinia nubilalis. devil in the details. Trends Ecol. Evol. 22, 80–86. doi: 10.1016/j.tree.2006. Physiol. Entomol. 40, 309–316. doi: 10.1111/phen.12116 10.008 Bronstein, J. L. (1994). Our current understanding of mutualism. Q. Rev. Biol. Haverkamp, A., Bing, J., Badeke, E., Hansson, B. S., and Knaden, M. (2016). 69, 31–51. doi: 10.1086/418432 Innate olfactory preferences for flowers matching proboscis length ensure Bronstein, J. L., Huxman, T. E.g., and Davidowitz, G. (2007). “Plant-mediated optimal energy gain in a hawkmoth. Nat. Commun. 7:11644. doi: 10.1038/ effects linking herbivory and pollination,” in Ecological Communities: Plant ncomms11644 Mediation in Indirect Interaction Webs. eds. T. Ohgushi, T. G. Craig and P. Hedrick, T. L., and Daniel, T. L. (2006). Flight control in the hawkmoth Manduca W. Price (Cambridge: Cambridge University Press), 79–103. sexta: the inverse problem of hovering. J. Exp. Biol. 209, 3114–3130. doi: Bronstein, J. L., Huxman, T., Horvath, B., Farabee, M., and Davidowitz, G. 10.1242/jeb.02363 (2009). Reproductive biology of Datura wrightii: the benefits of associating Herrera, C. M. (1987). Components of pollinator “quality”: comparative analysis with an herbivorous pollinator. Ann. Bot. 103, 1435–1443. doi: 10.1093/aob/ of a diverse insect assemblage. Oikos 50, 79–90. doi: 10.2307/ mcp053 3565403 Chai, P., and Srygley, R. B. (1990). Predation and the flight, morphology, and Hunt, J., and Simmons, L. W. (2000). Maternal and paternal effects on offspring temperature of neotropical rain-forest butterflies. Am. Nat. 135, 748–765. phenotype in the dung beetle Onthophagus taurus. Evolution 54, 936–941. doi: 10.1086/285072 doi: 10.1111/j.0014-3820.2000.tb00093.x Chippindale, A. K., Leroi, A. M., Kim, S. B., and Rose, M. R. (1993). Phenotypic Jacobsen, D. J., and Raguso, R. A. (2018). Lingering effects of herbivory and plasticity and selection in drosophila life-history evolution. I. Nutrition and plant defenses on pollinators. Curr. Biol. 28, R1164–R1169. doi: 10.1016/j. the cost of reproduction. J. Evol. Biol. 6, 171–193. doi: 10.1046/j.1420-9101. cub.2018.08.010 1993.6020171.x Jervis, M. A., Boggs, C. L., and Ferns, P. N. (2005). Egg maturation strategy Contreras, H. L., Goyret, J., von Arx, M., Pierce, C. T., Bronstein, J. L., and its associated trade-offs: a synthesis focusing on Lepidoptera. Ecol. Raguso, R. A., et al. (2013). The effect of ambient humidity on the foraging Entomol. 30, 359–375. doi: 10.1111/j.0307-6946.2005.00712.x behavior of the hawkmoth Manduca sexta. J. Comp. Physiol. A. 199, 1053–1063. Johnson, C. G. (1963). Physiological factors in insect migration by flight. Nature doi: 10.1007/s00359-013-0829-3 198, 423–427. doi: 10.1038/198423a0 Davidowitz, G., D’Amico, L. J., and Nijhout, H. F. (2003). Critical weight in Johnson, C. A., Smith, G. P., Yule, K., Davidowitz, G., Bronstein, J. L., and the development of insect body size. Evol. Dev. 5, 188–197. doi: 10.1046/j. Ferriere, R. (2021). Coevolutionary transitions from antagonism to mutualism 1525-142X.2003.03026.x explained by the co-opted antagonist hypothesis. Nat. Commun. 12:2867. Descamps, S., Gaillard, J. M., Hamel, S., and Yoccoz, N. G. (2016). When doi: 10.1038/s41467-021-23177-x relative allocation depends on total resource acquisition: implication for Kaitala, A. (1987). Dynamic life-history strategy of the Waterstrider Gerris the analysis of trade-offs. J. Evol. Biol. 29, 1860–1866. doi: 10.1111/jeb. thoracicus as an adaptation to food and habitat variation. Oikos 48, 125–131. 12901 doi: 10.2307/3565848 Frontiers in Plant Science | www.frontiersin.org 7 April 2022 | Volume 13 | Article 843506
Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory Karlsson, B. (1998). Nuptial gifts, resource budgets, and reproductive output Morehouse, N. I., and Rutowski, R. L. (2010). Developmental responses to in a polyandrous butterfly. Ecology 79, 2931–2940. doi: 10.1890/0012- variable diet composition in a butterfly: the role of nitrogen, carbohydrates 9658(1998)079[2931:NGRBAR]2.0.CO;2 and genotype. Oikos 119, 636–645. doi: 10.1111/j.1600-0706.2009.17866.x Karlsson, B., and Johansson, A. (2008). Seasonal polyphenism and developmental Mountcastle, A. M., Ravi, S., and Combes, S. A. (2015). Nectar vs. pollen trade-offs between flight ability and egg laying in a pierid butterfly. Proc. loading affects the tradeoff between flight stability and maneuverability in R. Soc. B 275, 2131–2136. doi: 10.1098/rspb.2008.0404 bumblebees. Proc. Natl. Acad. Sci. U. S. A. 112, 10527–10532. doi: 10.1073/ Kato, M., and Kawakita, A. (eds.) (2017). Obligate Pollination Mutualism. Japan: pnas.1506126112 Springer. Mousseau, T. A., and Dingle, H. (1991). Maternal effects in insect life histories. Kessler, A., and Chautá, A. (2020). The ecological consequences of herbivore- Annu. Rev. Entomol. 36, 511–534. doi: 10.1146/annurev.en.36.010191.002455 induced plant responses on plant–pollinator interactions. Emerg. Top. Life Mousseau, T. A., and Fox, C. W. (1998). The adaptive significance of maternal Sci. 4, 33–43. doi: 10.1042/ETLS20190121 effects. Trends Ecol. Evol. 13, 403–407. doi: 10.1016/S0169-5347(98)01472-4 King, E. G., Roff, D. A., and Fairbairn, D. J. (2011). Trade-off acquisition and Nestel, D., Papadopoulos, N. T., Pascacio-Villafán, C., Righini, N., allocation in Gryllus firmus: A test of the Y model. J. Evol. Biol. 24, 256–264. Altuzar-Molina, A. R., and Aluja, M. (2016). Resource allocation and doi: 10.1111/j.1420-9101.2010.02160.x compensation during development in holometabolous insects. J. Insect Physiol. Kooijman, B. (2009). Dynamic Energy Budget Theory for Metabolic Organisation. 95, 78–88. doi: 10.1016/j.jinsphys.2016.09.010 Cambridge: Cambridge University Press. Nicolson, S. W., Nepi, M., and Pacini, E. (eds.) (2007). Nectaries and Nectar. Kunte, K. (2007). Allometry and functional constraints on proboscis lengths Dordrecht: Springer. in butterflies. Funct. Ecol. 21, 982–987. doi: 10.1111/j.1365-2435.2007. Nicolson, S. W., and Thornburg, R. W. (2007). “Nectar chemistry,” in Nectaries 01299.x and Nectar. eds. S. W. Nicolson, M. Nepi and E. Pacini (United States: Lanza, J., Smith, G. C., Sack, S., and Cash, A. (1995). Variation in nectar Springer), 215–264. volume and composition of Impatiens capensis at the individual, plant, and Nijhout, H. F., and Emlen, D. J. (1998). Competition among body parts in population levels. Oecologia 102, 113–119. doi: 10.1007/BF00333318 the development and evolution. PNAS 95, 3685–3689. doi: 10.1073/ Lebeau, J., Wesselingh, R. A., and Van Dyck, H. (2016). Nectar resource pnas.95.7.3685 limitation affects butterfly flight performance and metabolism differently in Núñez-Farfán, J., Fornoni, J., and Valverde, P. L. (2007). The evolution of intensive and extensive agricultural landscapes. Proc. R. Soc. B 283:20160455. resistance and tolerance to herbivores. Annu. Rev. Ecol. Evol. Syst. 38, 541–566. doi: 10.1098/rspb.2016.0455 doi: 10.1146/annurev.ecolsys.38.091206.095822 Lehndal, L., and Ågren, J. (2015). Herbivory differentially affects plant fitness O’Brien, D. M. (1999). Fuel use in flight and its dependence on nectar feeding in three populations of the perennial herb Lythrum salicaria along a latitudinal in the hawkmoth Amphion floridensis. J. Exp. Biol. 202, 441–451. doi: 10.1242/ gradient. PLoS One 10:e0135939. doi: 10.1371/journal.pone.0135939 jeb.202.4.441 Levin, E., McCue, M. D., and Davidowitz, G. (2017a). Sex differences in the O’Brien, D. M., Boggs, C. L., and Fogel, M. L. (2004). Making eggs from nectar: utilization of essential and non-essential amino acids in Lepidoptera. J. Exp. the role of life history and dietary carbon turnover in butterfly reproductive Biol. 220, 2743–2747. doi: 10.1242/jeb.154757 resource allocation. Oikos 105, 279–291. doi: 10.1111/j.0030-1299.2004.13012.x Levin, E., McCue, M. D., and Davidowitz, G. (2017b). More than just sugar: O’Brien, D. M., Fogel, M. L., and Boggs, C. L. (2002). Renewable and nonrenewable allocation of nectar amino acids and fatty acids in a lepidopteran. Proc. R. resources: amino acid turnover and allocation to reproduction in Lepidoptera. Soc. B 284:20162126. doi: 10.1098/rspb.2016.2126 PNAS 99, 4413–4418. doi: 10.1073/pnas.072346699 Levin, E., Mitra, C., and Davidowitz, G. (2016). Fed males increase oviposition Ollerton, J., Winfree, R., and Tarrant, S. (2011). How many flowering plants are in female hawkmoths via non-nutritive direct benefits. Anim. Behav. 112, pollinated by animals? Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x 111–118. doi: 10.1016/j.anbehav.2015.11.019 Papaj, D. R. (2000). Ovarian dynamics and host use. Annu. Rev. Entomol. 45, Lewis, S., and South, A. (2012). “The evolution of animal nuptial gifts,” in 423–448. doi: 10.1146/annurev.ento.45.1.423 Advances in the Study of Behavior. eds. H. Jane Brockmann, T. J. Roper, Pasquet, R. S., Peltier, A., Hufford, M. B., Oudin, E., Saulnier, J., Paul, L., M. Naguib, J. C. Mitani and L. W. Simmons (Academic Press), 53–97. et al. (2008). Long-distance pollen flow assessment through evaluation of Lucas-Barbosa, D. (2016). Integrating studies on plant–pollinator and plant– pollinator foraging range suggests transgene escape distances. Proc. Natl. herbivore interactions. Trends Plant Sci. 21, 125–133. doi: 10.1016/j. Acad. Sci. U. S. A. 105, 13456–13461. doi: 10.1073/pnas.0806040105 tplants.2015.10.013 Raguso, R. A., and Willis, M. A. (2002). Synergy between visual and olfactory Marquis, R. (1992). “Selective impact of herbivores,” in Plant Resistance to cues in nectar feeding by naıve hawkmoths, Manduca sexta. Anim. Behav. Herbivores and Pathogens: Ecology, Evolution, and Genetics. eds. R. S. Fritz 64, 685–695. doi: 10.1006/anbe.2002.4010 and E. L. Simms (Chicago: University of Chicago Press), 301–325. Rankin, M. A., and Burchsted, J. C. A. (1992). The cost of migration in insects. Mattson, W. J. (1980). Herbivory in relation to plant nitrogen content. Annu. Annu. Rev. Entomol. 37, 533–559. doi: 10.1146/annurev.en.37.010192.002533 Rev. Ecol. Syst. 11, 119–161. doi: 10.1146/annurev.es.11.110180.001003 Riffell, J. A., and Alarcón, R. (2013). Multimodal floral signals and moth foraging McCall, A. C., Davidowitz, G., and Bronstein, J. L. (2020). How high are the decisions. PLoS One 8:e72809. doi: 10.1371/journal.pone.0072809 costs inflicted by an herbivorous pollinator? Arthropod Plant Interact. 14, Roach, D. A., and Wulff, R. D. (1987). Maternal effects in plants. Annu. Rev. 387–397. doi: 10.1007/s11829-020-09745-w Ecol. Syst. 18, 209–235. doi: 10.1146/annurev.es.18.110187.001233 McCallum, K. P., McDougall, F. O., and Seymour, R. S. (2013). A review of Roff, D. A. (1986). The evolution of wing dimorphism in insects. Evolution the energetics of pollination biology. J. Comp. Physiol. B. 183, 867–876. doi: 40, 1009–1020. doi: 10.1111/j.1558-5646.1986.tb00568.x 10.1007/s00360-013-0760-5 Roff, D. A. (1990). Selection for changes in the incidence of wing dimorphism Meslin, C., Cherwin, T. S., Plakke, M. S., Hill, J., Small, B. S., Goetz, B. J., in Gryllus-Firmus. Heredity 65, 163–168. doi: 10.1038/hdy.1990.84 et al. (2017). Structural complexity and molecular heterogeneity of a butterfly Roff, D. A. (1994). Habitat persistence and the evolution of wing dimorphism ejaculate reflect a complex history of selection. Proc. Natl. Acad. Sci. U. S. A. in insects. Am. Nat. 144, 772–798. doi: 10.1086/285706 114, E5406–E5413. doi: 10.1073/pnas.1707680114 Rotem, K., Agrawal, A. A., and Kott, L. (2003). Parental effects in Pieris rapae Mevi-Schütz, J., and Erhardt, A. (2003). Larval nutrition affects female nectar in response to variation in food quality: adaptive plasticity across generations? amino acid preference in the map butterfly (Araschnia levana). Ecology 84, Ecol. Entomol. 28, 211–218. doi: 10.1046/j.1365-2311.2003.00507.x 2788–2794. doi: 10.1890/02-3108 Rusman, Q., Lucas-Barbosa, D., Poelman, E. H., and Dicke, M. (2019). Ecology Mevi-Schütz, J., and Erhardt, A. (2005). Amino acids in nectar enhance butterfly of plastic flowers. Trends Plant Sci. 24, 725–740. doi: 10.1016/j.tplants.2019. fecundity: a long-awaited link. Am. Nat. 165, 411–419. doi: 10.1086/ 04.007 429150 Rutowski, R. L. (1985). Evidence for mate choice in a Sulphur butterfly (Colias Mitra, C., Reynoso, E., Davidowitz, G., and Papaj, D. (2016). Effects of sodium- eurytheme). Z. Tierpsychol. 70, 103–114. doi: 10.1111/j.1439-0310.1985.tb00504.x puddling on male mating success, courtship, and flight in a swallowtail Saeki, Y., Tuda, M., and Crowley, P. H. (2014). Allocation tradeoffs and life butterfly. Anim. Behav. 114, 203–210. doi: 10.1016/j.anbehav.2016. histories: a conceptual and graphical framework. Oikos 123, 786–793. doi: 01.028 10.1111/oik.00956 Frontiers in Plant Science | www.frontiersin.org 8 April 2022 | Volume 13 | Article 843506
Davidowitz et al. Flight, Fecundity Links Pollination, and Herbivory
Sasaki, M., and Riddiford, L. M. (1984). Regulation of reproductive behaviour Watanabe, M., and Hirota, M. (1999). Effects of sucrose intake on spermatophore
and egg maturation in the tobacco hawk moth, Manduca sexta. Physiol. mass produced by male swallowtail butterfly Papilio xuthus L. Zool. Sci.
Entomol. 9, 315–327. doi: 10.1111/j.1365-3032.1984.tb00713.x 16, 55–61. doi: 10.2108/zsj.16.55
Schulke, B., and Waser, N. M. (2001). Long-distance pollinator flights and Wedell, N., and Karlsson, B. (2003). Paternal investment directly affects female
pollen dispersal between populations of Delphinium nuttallianum. Oecologia reproductive effort in an insect. Proc. R. Soc. B 270, 2065–2071. doi: 10.1098/
127, 239–245. doi: 10.1007/s004420000586 rspb.2003.2479
Simpson, S. J., and Simpson, C. L. (1990). “The mechanisms of nutritional Wheeler, D. (1996). The role of nourishment in oogenesis. Annu. Rev. Entomol.
compensation by phytophagous insects,” in Insect-Plant Interactions. ed. E. A. 41, 407–431. doi: 10.1146/annurev.en.41.010196.002203
Bernays (Boca Raton, Florida: CRC Press), 111–160. Wiklund, C., Kaitala, A., and Wedell, N. (1998). Decoupling of reproductive
Sisodia, S., and Singh, B. N. (2012). Experimental evidence for nutrition regulated rates and parental expenditure in a polyandrous butterfly. Behav. Ecol. 9,
stress resistance in Drosophila ananassae. PLoS One 7:e46131. doi: 10.1371/ 20–25. doi: 10.1093/beheco/9.1.20
journal.pone.0046131 Willis, M. A., and Arbas, E. A. (1991). Odor-modulated upwind flight of the
Slansky, F. Jr. (1978). Utilization of energy and nitrogen by larvae of the sphinx moth, Manduca sexta L. J. Comp. Physiol. A. 169, 427–440. doi:
imported cabbageworm, Pieris rapae, as affected by parasitism by Apanteles 10.1007/BF00197655
glomeratus. Environ. Entomol. 7, 179–185. doi: 10.1093/ee/7.2.179 Willmer, P. (2011). Pollination and Floral Ecology. Princeton, New Jersey:
Smith, G. P., Bronstein, J. L., and Papaj, D. R. (2019). Sex differences in Princeton University Press.
pollinator behavior: patterns across species and consequences for the mutualism. Wilson, J. K., Ruiz, L., Duarte, J., and Davidowitz, G. (2019). The nutritional
J. Anim. Ecol. 88, 971–985. doi: 10.1111/1365-2656.12988 landscape of host plants for a specialist insect herbivore. Ecol. Evol. 9,
Soteras, F., Rubini Pisano, M. A., Bariles, J. B., Moré, M., and Cocucci, A. A. 13104–13113. doi: 10.1002/ece3.5730
(2020). Phenotypic selection mosaic for flower length influenced by Zera, A. J., and Brink, T. (2000). Nutrient absorption and utilization by wing
geographically varying hawkmoth pollinator proboscis length and abiotic and flight muscle morphs of the cricket Gryllus firmus: implications for
environment. New Phytol. 225, 985–998. doi: 10.1111/nph.16192 the trade-off between flight capability and early reproduction. J. Insect Physiol.
Sprayberry, J. D., and Daniel, T. L. (2007). Flower tracking in hawkmoths: 46, 1207–1218. doi: 10.1016/S0022-1910(00)00041-X
behavior and energetics. J. Exp. Biol. 210, 37–45. doi: 10.1242/jeb.02616 Zera, A. J., and Harshman, L. G. (2001). The physiology of life history trade-
Stearns, S. C. (1989). Trade-offs in life-history evolution. Funct. Ecol. 3, 259–268. offs in animals. Annu. Rev. Ecol. Syst. 32, 95–126. doi: 10.1146/annurev.
doi: 10.2307/2389364 ecolsys.32.081501.114006
Stjernholm, F., Karlsson, B., and Boggs, C. L. (2005). Age-related changes in Zera, A. J., and Larsen, A. (2001). The metabolic basis of life history variation:
thoracic mass: possible reallocation of resources to reproduction in butterflies. genetic and phenotypic differences in lipid reserves among life history morphs
Biol. J. Linn. Soc. 86, 363–380. doi: 10.1111/j.1095-8312.2005.00542.x of the wing-polymorphic cricket, Gryllus firmus. J. Insect Physiol. 47, 1147–1160.
Svärd, L., and Wiklund, C. (1989). Mass and production rate of ejaculates in doi: 10.1016/S0022-1910(01)00096-8
relation to monandry/polyandry in butterflies. Behav. Ecol. Sociobiol. 24, Zera, A. J., Sall, J., and Otto, K. (1999). Biochemical aspects of flight
395–402. doi: 10.1007/BF00293267 and flightlessness in Gryllus: flight fuels, enzyme activities and
Tigreros, N. (2013). Linking nutrition and sexual selection across life stages in a electrophoretic profiles of flight muscles from flight-capable and flightless
model butterfly system. Funct. Ecol. 27, 145–154. doi: 10.1111/1365-2435.12006 morphs. J. Insect Physiol. 45, 275–285. doi: 10.1016/S0022-1910(98)
Tigreros, N., and Davidowitz, G. (2019). Flight-fecundity tradeoffs in wing- 00123-1
monomorphic insects. Adv. Insect Physiol. 56, 1–41. doi: 10.1016/bs.
aiip.2019.02.001 Conflict of Interest: The authors declare that the research was conducted in
Tigreros, N., Mowery, M. A., and Lewis, S. M. (2014). Male mate choice favors the absence of any commercial or financial relationships that could be construed
more colorful females in the gift-giving cabbage butterfly. Behav. Ecol. as a potential conflict of interest.
Sociobiol. 68, 1539–1547. doi: 10.1007/s00265-014-1764-1
Vahed, K. (1998). The function of nuptial feeding in insects: a review of Publisher’s Note: All claims expressed in this article are solely those of the
empirical studies. Biol. Rev. 73, 43–78. doi: 10.1017/S0006323197005112 authors and do not necessarily represent those of their affiliated organizations,
van Noordwijk, A. J., and de Jong, G. (1986). Acquisition and allocation of or those of the publisher, the editors and the reviewers. Any product that may
resources: their influence on variation in life history tactics. Am. Nat. 128, be evaluated in this article, or claim that may be made by its manufacturer, is
137–142. doi: 10.1086/284547 not guaranteed or endorsed by the publisher.
Von Arx, M., Goyret, J., Davidowitz, G., and Raguso, R. A. (2012). Floral
humidity as a reliable sensory cue for profitability assessment by nectar- Copyright © 2022 Davidowitz, Bronstein and Tigreros. This is an open-access article
foraging hawkmoths. Proc. Natl. Acad. Sci. U. S. A. 109, 9471–9476. doi: distributed under the terms of the Creative Commons Attribution License (CC BY).
10.1073/pnas.1121624109 The use, distribution or reproduction in other forums is permitted, provided the original
von Arx, M., Sullivan, K. A., and Raguso, R. A. (2013). Dual fitness benefits author(s) and the copyright owner(s) are credited and that the original publication in
of post-mating sugar meals for female hawkmoths (Hyles lineata). J. Insect this journal is cited, in accordance with accepted academic practice. No use, distribution
Physiol. 59, 458–465. doi: 10.1016/j.jinsphys.2013.01.006 or reproduction is permitted which does not comply with these terms.
Frontiers in Plant Science | www.frontiersin.org 9 April 2022 | Volume 13 | Article 843506You can also read

















































