Declines in wintering bird richness over 14 years in Northern California - UC Natural Reserve System
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Declines in wintering bird richness
over 14 years in Northern California
Elise Gonzales1, Charin Park2, Andie Van Horn2
1
University of California, San Diego; 2University of California, Santa Barbara
ABSTRACT
Climate change is creating shifts in seasonal timing. The effects of climate change on
California can be seen through decreased precipitation and increased fire frequency in
inland communities. The state is along a major migratory passageway and is home to
populations of resident birds. Lake County’s chaparral, grassland, and woodland habitats
are hotspots of diversity. This area experienced multiple major fires in the past decade.
In this study, we conducted point count surveys to investigate bird communities, which
we compared to historic data from the early 2000s. We found that overall species richness
shifted since the initial study due to a decrease in populations of resident birds. There
was greater overall species richness in oak woodland habitat. Insectivore and herbivore
richness were greater in areas of complex vegetation. Burn history and serpentine soil
had no effect on bird abundance or richness. These observations demonstrate the
importance of long-term data collection and identify habitats which should be prioritized
for future management.
Keywords: Wintering birds, dietary niche, phenology, serpentine, climate change
INTRODUCTION term studies are especially powerful for
tracking responses as they emerge (Gibbs
Climate change has had catastrophic and Breisch 2001), Lindenmayer et al. 2012).
effects on biodiversity. Shifts in seasonal In California, climate change has been
timing have been shown to hamper studied through a variety of abiotic factors.
reproductive success in many species, as California’s droughts are a result of
fluctuating temperatures and crashes in prey precipitation deficits co-occurring with
abundance have led to the starvation or warming conditions; these conditions have
freezing of young (Crick 2004). Due to their increased in recent decades (Diffenbaugh et
ectothermy and limited vagility, amphibians al. 2015). Inland areas of California face both
are especially sensitive to changing climate, drier conditions and increased fire frequency
and are frequently-studied systems for (Hayhoe et al. 2004).
observing the effects of altered climate on The biota of California is rich and diverse.
population dynamics (BeeBee 1995). Home to two-thirds of North America’s bird
However, the impacts of climate change species, the state is located at the center of
differ across study systems. For migratory a major north-south migratory passage
organisms such as mammals and birds, long extending from Patagonia to the Arctic
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 1/9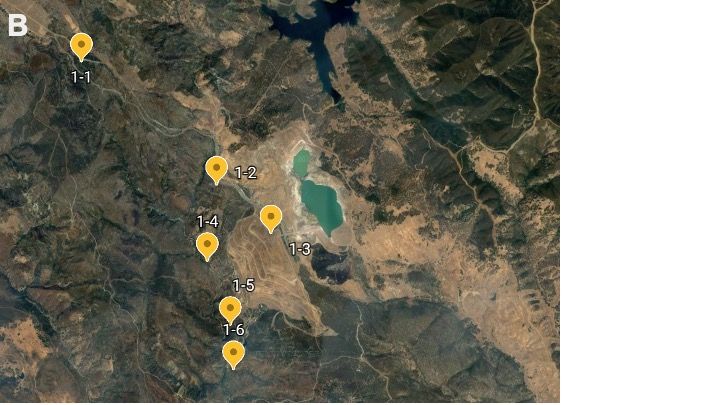
tundra (Grinnell and Miller 1944). Every 3. How do bird communities compare
year, millions of waders, raptors, waterfowl, across habitats (including serpentine
and passerines seek refuge in the state’s and non-serpentine habitats)?
heterogeneous landscape.
We expected bird communities of the
Lake County, California is a biodiversity
early 2000s and of 2021 to differ, with the
hotspot for the state’s bird populations.
most marked decreases in areas of more
Over 300 species rely on its montane
recent burning (Roberts et al. 2012). We
chaparral, grasslands, and oak woodlands
expected the highest species richness and
(eBird 2021). Due to the area’s persistent
abundance in the oak woodland habitats
drought and wildfires of recent decades, the
because of its vegetation structure, which
need for long-term avian community studies
provides a greater variety of nest and perch
has become crucial, as these studies have
sites, as well as greater foraging
the potential to create a better
opportunities (DeGraaf 1991). Because
understanding of how these communities
species have greater reproductive success
are impacted by climate change.
and higher richness and abundances where
In this study, we investigated winter bird
their requirements are met, we expected to
communities across habitats of McLaughlin
see a relationship between dietary niche and
Natural Reserve, within Lake and Napa
habitat type (DeGraaf 1991). Lastly, we
Counties of Northern California. While
hypothesized a lower abundance and
citizen science data on bird communities
species richness in serpentine habitats due
exists in the larger Lake County region, it has
to anecdotal observations made by the
limitations in its conservation applications
McLaughlin Natural Reserve director (Cathy
due to overrepresentation of accessible
Koehler pers. comm. 2021).
areas. The most recent avian surveys at the
reserve were conducted from 2004 to 2007. METHODS
These surveys were originally meant to be
long-term research; however, they were not 2.1 Study System
maintained. Since then, the reserve has
experienced two high-intensity wildfires – The Donald and Sylvia McLaughlin Natural
the 2015 Jerusalem Fire and the 2020 SCU Reserve, located in the Blue Ridge Berryessa
Lightning Complex. There was a decline in Natural Area, in Lake and Napa County,
precipitation, as seen in California’s drought California, is a 7,050-acre reserve with
period from 2012-2014 (Williams et al. varying plant communities, including those
2015). To assess changes in bird characterized by serpentine soils, and others
communities, defined as species richness more typical of coastal mountain range
and total abundance, since the early 2000s, habitats, such as oak woodlands, grasslands,
we investigated three main questions: and chaparral (Callizo 1992). We surveyed all
the aforementioned habitat types. The
1. How have bird communities changed
reserve had populations of both resident
over the past decades?
and wintering birds.
2. How do bird communities respond to
fire?
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 2/9
2.2 Data Collection
We used historical point count data from
McLaughlin Natural Reserve Stewardship
Director, Paul Aigner. We used data from
December–January of 2004, 2006, and 2007
to control for seasonal variance. We
conducted our survey from February 23-28,
2021. Despite controlling for seasonal
variance, the weather on 2021 survey days
was particularly windy.
To estimate avian abundance and assess
species richness across habitats, we
conducted point counts with fixed-distance
circular plots (radius = 100 meters). We
surveyed 19 sites at McLaughlin Natural
Reserve (Fig. 1), all of which were surveyed
in the original bird study. We did this to allow
for direct comparison in our analysis. We
repeated the protocol used in the earlier
surveys as follows: Upon arriving at a site, we
waited 5 minutes before surveying to
account for possible disturbance caused by
our arrival. We recorded habitat type
(grassland, man-made grassland, oak
woodland, chaparral, cypress chaparral, or
chamise chaparral), burn year (1999, 2015,
or not burned) and serpentine (present or
absent). We observed for 9 minutes and
recorded all birds sighted. We took note of
distance (less than 50 meters, 50–100
meters and flyovers) from the point to Figure 1, 1A, 1B. Map of sites. Map of McLaughlin
account for possible edge effects from Natural Reserve. 1: the entire reserve; 1A: sites on the
differing habitats that neighbored each site. west side of the reserve; 1B: sites on the east side of
During these point counts, we used both the reserve. Each map point represents the site of a
point count survey with fixed-distance circular plots
binoculars and our naked eye. We
(radius = 100 m). At each point count, species and
referenced The Sibley Field Guide to Birds of abundance of all birds were recorded. Sites visited in
Western North America for identifications 2021 follow surveys conducted in the early 2000s.
(Sibley 2003).
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 3/9
2.3 Data Analysis 3.1 Changes Over Time
We used JMP Pro 15 and JMP Student 14 Bird richness declined from the 2000s to
for all statistical analyses. 2021 (n =189, F = 12.4487, P < 0.01), with 27
To test for changes in bird communities, fewer bird species in the present day than in
we included all available bird data to run an earlier surveys. The majority of differences
ANOVA test on the effect of time (2000s or in species richness by 2021 were due to a
present) on bird communities. We also used reduction in resident species (Table 1).
all available bird data to run an ANOVA on There was no significant difference in bird
the effect of burn year on bird communities. abundance from the 2000s to the present.
We used ANOVA tests to analyze the effect However, we observed a 50% lower average
of habitat and serpentine on bird total abundance in 2021 than in the 2000s
communities. To better understand the (n= 189, F = 2.1234, P = 0.1467). The greatest
effects of habitat on various bird dietary declines in count were observed in three
niches, we categorized species as carnivores, resident bird species: the lesser goldfinch
scavengers, herbivores, omnivores, and (Spinus psaltria), American robin (Turdus
insectivores based on life history migratorius), and lark sparrow (Chondestes
information obtained through Cornell’s All grammacus) (Fig. 2). All three species were
About Birds (Cornell Lab of Ornithology observed in the early 2000s; however, they
2019). We then used ANOVA tests to analyze were absent from the 2021 counts.
the effects of habitat on communities of
specific dietary niches as listed above.
RESULTS
Across all survey years (2004, 2006, 2007,
and 2021), 57 bird species were observed. 47
species were observed in the early 2000s
and 28 species were observed in 2021. 8
species were exclusive to the latter survey.
Bird species new to the 2021 dataset
consisted of the Cooper’s hawk (Accipiter
cooperii), American kestrel (Falco
sparverius), white-tailed kite (Elanus Figure 2. Relationship between species richness and
leucurus), killdeer (Charadrius vociferus), time. The average richness of bird species across two
red-winged blackbird (Agelaius phoeniceus), different time periods at McLaughlin Natural Reserve
based on all available bird data. Species richness was
rufous hummingbird (Selasphorus rufus),
two times greater in the 2000s than in the present
Canada goose (Branta canadensis), and an day. This decline was mainly a result of decreased
unidentified duck (no ducks were sighted in richness of resident bird species (Table 1). Vertical
the early 2000s data) (Table 1). bars represent means ± 1 S.E.
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 4/9
Table 1. Species that differ between early 2000s and present. All species that were recorded only in the early 2000s
(2004-2007) or only in 2021. 8 species were exclusive to the present-day data (one was a pair of unidentified ducks
and was omitted from the table). n represents the total abundance of a species across the entire survey. Migratory
habit indicates the species’ level of residency at the reserve. Resident species should be present year-round,
wintering species should be present during the winter season; breeding and migrant species are indicated with the
appropriate season. Migratory habit information was sourced from Cornell’s All About Birds (Cornell Lab of
Ornithology 2019). McLaughlin Natural Reserve’s species richness decreased from the early 2000s to 2021 (Fig. 2).
Most species absent in the present-day survey were resident birds.
Species Migratory Habit n Early 2000s Present
Cooper’s hawk Accipiter cooperii Resident 1 Absent Present
Merlin Falco columbarius Wintering 2 Present Absent
Peregrine falcon Falco peregrinus Resident 1 Present Absent
White-tailed kite Elanus leucurus Resident 1 Absent Present
Northern harrier Circus hudsonius Resident 1 Present Absent
Common kestrel Falco tinnunculus Resident 3 Absent Present
Bewick’s wren Thryomanes bewickii Resident 11 Present Absent
Polioptila caerulea Breeding
Blue-grey gnatcatcher 2 Present Absent
(April to June)
Bushtit Psaltriparus minimus Resident 21 Present Absent
California thrasher Toxostoma redivivum Resident 6 Present Absent
Hermit thrush Catharus guttatus Wintering 6 Present Absent
Hutton’s vireo Vireo huttoni Resident 2 Present Absent
Killdeer Charadrius vociferus Resident 1 Absent Present
Oak titmouse Baeolophus inornatus Resident 6 Present Absent
Red-breasted sapsucker Sphyrapicus ruber Wintering 1 Present Absent
Ruby-crowned kinglet Regulus calendula Wintering 12 Present Absent
Red-winged blackbird Agelaius phoeniceus Resident 30 Absent Present
Song sparrow Melospiza melodia Resident 2 Present Absent
White-breasted nuthatch Sitta carolinensis Resident 5 Present Absent
Western bluebird Sialia mexicana Resident 30 Present Absent
Yellow-rumped warbler Setophaga coronata Resident 4 Present Absent
Varied thrush Ixoreus naevius Wintering 6 Present Absent
Northern mockingbird Mimus polyglottos Resident 1 Present Absent
Cedar waxwing Bombycilla cedrorum Wintering 12 Present Absent
Anna’s hummingbird Calypte anna Resident 3 Present Absent
Selasphorus rufus Migrant
Rufous hummingbird 1 Absent Present
(spring)
California quail Callipepla californica Resident 5 Present Absent
Dark-eyed junco Junco hyemalis Resident 26 Present Absent
Haemorhous
House finch Resident 17 Present Absent
mexicanus
Chondestes
Lark sparrow Resident 108 Present Absent
grammacus
Canada goose Branta canadensis Resident 2 Absent Present
Lesser goldfinch Spinus psaltria Resident 108 Present Absent
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 5/93.2 Habitat Impacts
Habitat affected overall species richness,
as well as the richness of specific dietary
niches. The overall species richness in oak
woodland habitats was higher than
grasslands, man-made grasslands, cypress
chaparral, and chaparral habitats (Fig. 3, P <
0.01). The species richness of insectivores Figure 3. Species richness across habitats. The
(Fig. 4, P < 0.0001) and herbivores (Fig. 4, P < average richness of bird species across six different
0.001) were also impacted by habitat. habitats at McLaughlin Natural Reserve based on
Insectivore richness followed the same trend early 2000s (2004–2007) and recent (2021) data.
as was seen in overall richness. Herbivore Birds seen >50 meters away, >100 meters away, and
flyovers were excluded to control for direct
richness was similar in chamise chaparrals interaction with the site’s habitat. Habitat impacted
and oak woodlands; greater in chamise species richness. Richness was greater in areas of
chaparral when compared to grassland, more complex vegetation, as shown through greater
man-made grassland, and cypress chaparral; richness in oak woodlands, compared to grassland,
and greater in oak woodlands than in man- man-made grassland, chaparral, and cypress
chaparral. Vertical bars represent means ± 1 S.E.
made grasslands. We found habitat had no
effect on the species richness of carnivores,
omnivores, and scavengers. There was no
effect of habitat on total abundance or
abundance of specific dietary niches. There
were also no differences in the distributions
of bird communities across habitats
between the early 2000s and 2021.
3.3 Serpentine and Burning
Burn history and presence of serpentine Figure 4. Species richness across habitats based on
had no effect on species richness or dietary niche. The average richness of insectivores
abundance. and herbivores across six different habitats at
McLaughlin Natural Reserve based on early 2000s
DISCUSSION (2004-2007) and recent (2021) data. Birds seen >50
meters away, >100 meters away, and flyovers were
excluded to control for direct interaction with the
4.1 Changes Over Time
site’s habitat. Habitat affected the average richness of
insectivores and herbivores. Insectivores followed
We speculate that the decline in average the same trend as overall richness (Figure 3).
species richness may be a response to Herbivore richness was similar in chamise chaparrals
gradual, site-level environmental shifts such and oak woodlands; greater in chamise chaparral
as climate change (Rodenhouse et al. 2009, when compared to grassland, man-made grassland,
and cypress chaparral habitats; and greater in oak
Lehikoinen and Virkkala 2017). Changes in
woodlands than in man-made grasslands. Vertical
the timing of breeding, as a result of earlier bars represent means ± 1 S.E.
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 6/9warming, has been shown to impact the 4.2 Varying Habitats
reproductive success of residents and
migrants differently, indicating potential for Greater richness at oak woodlands could
changes in competitive dynamics (Ahola et be due to the complex vegetation present;
al. 2007). There is evidence that the increased complexity leads to increased
distribution of resident bird species (who richness (MacArthur and MacArthur 1961).
represent the majority of the decline in This increased complexity likely provides the
richness) shows greater sensitivity towards vegetation necessary for providing shelter
our shifting climate (Lehikoinen and Virkkala and more abundant foraging opportunities
2017), suggesting that altered ambient for birds (DeGraaf 1991). The high
temperatures over the last 14 years may insectivore richness at oak woodlands and
have affected bird richness at McLaughlin high herbivore richness at both chamise
Natural Reserve. chaparral and oak woodland habitats could
Differences in abundance were not be attributed to the way in which these
significant across years. However, resident dietary niches rely on habitat. This trend in
species, such as the American robin, lesser insectivores and herbivores may indicate
goldfinch, and lark sparrow, were abundant that their diets tie them to particular
in December to January of 2004 to 2007, but habitats, likely because their dietary needs
absent from our 2021 survey (Table 1). As are best met in those habitats. However, this
the 2021 survey was conducted in late trend was not found for carnivores,
February, it is possible these differences omnivores, or scavengers. This suggests that
were due to phenological shifts unrelated to these three dietary niches exploit resources
migratory habits. The aforementioned more opportunistically and, as such, can
resident birds, who account for a majority of utilize a greater variety of habitats to fulfill
the observed decline in abundance, form their dietary requirements (Wiens &
large feeding flocks in winter, but become Rotenberry 1979).
territorial and travel in mated pairs during
spring (Adamson 2000). As such, it is possible Implications
that the observed decrease in total
From the early 2000s to 2021, the richness
abundance in February 2021 was due to the
of McLaughlin’s wintering birds declined by
resident species beginning to travel in
over half, while total abundance of observed
mating pairs as spring approaches, rather
birds was not impacted by time, indicating
than the large flocks observed in the initial
spatial and temporal shifts in population
studies, which were conducted earlier in the
dynamics. Declines in species richness at this
winter season. Additionally, decreased
location could be indicative of how
abundance in 2021 may have been caused,
disturbance may be impacting avian
in part, by the high winds; birds, particularly
communities of neighboring regions.
those that migrate, will avoid flight during
Greater richness in oak woodlands suggests
windy periods (Åkesson and Hedenström
that these habitats should be prioritized in
1999).
the management of land and the
conservation of bird populations in the Lake
County area.
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 7/9The impacts of intense and frequent Cornell Lab of Ornithology. 2019. All about birds.
disturbance in Lake County habitats on avian Cornell Lab of Ornithology, Ithaca, New York.
https://www.allaboutbirds.org. Accessed on
communities is important to survey because March 2, 2021.
of the high species richness in this area.
Considering the gap in data since the early Crick, H. Q. P. 2004. The impact of climate change on
2000s, we advocate for more regular, long- birds. Ibis 146:48–56.
term monitoring in order to gain a clearer
Daniau, A.-L. et al. 2012. Predictability of biomass
picture of the effects of recent disturbances, burning in response to climate changes. Global
such as the impacts of the 2020 SCU Biogeochemical Cycles 26.
Lightning Complex fire, as well as more
gradual mechanisms, such as climate DeGraaf, Richard M. Forest and rangeland birds of the
change, which may be driving shifts in United States: natural history and habitat use. No.
688. United States Government Printing, 1991.
biodiversity.
Diffenbaugh, N. S., D. L. Swain, and D. Touma. 2015.
ACKNOWLEDGMENTS Anthropogenic warming has increased drought risk
in California. Proceedings of the National Academy
This work was performed at the University of Sciences 112:3931–3936.
of California’s McLaughlin Natural Reserve,
eBird. (n.d.). Lake, California.
doi:10.21973/N3W08D.
https://ebird.org/region/US-CA-033. Accessed on
March 2, 2021.
REFERENCES
Gibbs, J. P., and A. R. Breisch. 2001. Climate warming
Adamson, J. J. 2000. Final report to City of Boulder and calling phenology of frogs near Ithaca, New
Open Space Department testing female restriction York, 1900–1999. Conservation Biology 15:1175–
of male mating success: The suitability of lark 1178.
sparrows.
Grinnell, J., Jr, and A. H. Miller. 1986. The distribution
Ahola, M. P., T. Laaksonen, T. Eeva, and E. Lehikoinen. of the birds of California. The Condor 88:482–482.
2007. Climate change can alter competitive
relationships between resident and migratory Hayhoe, K., D. Cayan, C. B. Field, P. C. Frumhoff, E. P.
birds. Journal of Animal Ecology 76:1045–1052. Maurer, N. L. Miller, S. C. Moser, S. H. Schneider, K.
N. Cahill, E. E. Cleland, L. Dale, R. Drapek, R. M.
Avery, M. L., and C. van R. III. 1989. Seasonal changes Hanemann, L. S. Kalkstein, J. Lenihan, C. K. Lunch,
in bird communities of the chaparral and blue-oak R. P. Neilson, S. C. Sheridan, and J. H. Verville. 2004.
woodlands in central California. The Condor Emissions pathways, climate change, and impacts
91:288–295. on California. Proceedings of the National Academy
of Sciences 101:12422–12427.
Beebee, T. O. 1995. Talking maps: region and
revolution in Juan Vincent Benet and Euclides da Jenkins, D., and A. Watson. 2000. Dates of first arrival
Cunha. Comparative Literature 47:193–214. and song of birds during 1974–99 in mid-Deeside,
Scotland. Bird Study 47:249–251.
Callizo, J. 1992. Serpentine habitats for the rare plants
of Lake, Napa and Yolo Counties, California. In: Knopf, F. L., and B. J. Miller. 1994. Charadrius
Baker, A. J. M., J. Proctor and R. D. Reeves, The montanus: Montane, Grassland, or Bare-Ground
vegetation of ultramafic (serpentine) soils. pp. 35- Plover? The Auk 111:504–506.
51. Intercept Ltd., Andover, Hants., England.
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 8/9Lindenmayer, D. B., G. E. Likens, A. Andersen, D. Sibley, D. 2003. The Sibley field guide to birds of
Bowman, C. M. Bull, E. Burns, C. R. Dickman, A. A. western North America. 1st ed. Alfred A. Knopf,
Hoffmann, D. A. Keith, M. J. Liddell, A. J. Lowe, D. J. New York.
Metcalfe, S. R. Phinn, J. Russell-Smith, N. Thurgate,
and G. M. Wardle. 2012. Value of long-term Virkkala, R. and A. Lehikoinen. 2017. Birds on the
ecological studies. Austral Ecology 37:745–757. move in the face of climate change: High species
turnover in northern Europe. Ecology and Evolution
MacArthur, R. H., and J. W. MacArthur. 1961. On bird 7:8201–8209.
species diversity. Ecology 42:594–598.
Wiens, John A. and John T. Rotenberry. 1979. Diet
Rodenhouse, N. L., L. M. Christenson, D. Parry, and L. niche relationships among North American
E. Green. 2009. Climate change effects on native grassland and shrubsteppe birds. Oecologia 42, no.
fauna of northeastern forests. In: NE Forests 2100: 3:253–292.
A synthesis of climate change impacts on forests of
the northeastern US and eastern Canada. Canadian Wiens, J. A. and J. T. Rotenberry. 1979. Diet niche
Journal of Forest Research 39:249–263. relationships among North American grassland and
shrubsteppe birds. Oecologia 42:253–292.
Rosén, M., A. Hedenström, A. Badami, F. Spina, and S.
Åkesson. 1999. Hunting flight behaviour of the Williams, A. P., R. Seager, J. T. Abatzoglou, B. I. Cook,
Eleonora’s falcon Falco eleonorae. Journal of Avian J. E. Smerdon, and E. R. Cook. 2015. Contribution of
Biology 30:342–350. anthropogenic warming to California drought
during 2012–2014. Geophysical Research Letters
Şekercioğlu, Çağan H., Gretchen C. Daily, and Paul R. 42:6819–6828.
Ehrlich. 2004. Ecosystem consequences of bird
declines. Proceedings of the National Academy of
Sciences 101(52):18042-18047.
|
CEC Research https://doi.org/10.21973/N33088 Winter 2021, Vol. 5 Issue 1 9/9You can also read

















































