Berberine inhibits the progression of renal cell carcinoma cells by regulating reactive oxygen species generation and inducing DNA damage
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Berberine inhibits the progression of renal cell
carcinoma cells by regulating reactive oxygen
species generation and inducing DNA damage
Yuwan Zhao ( zhaoyw201@126.com )
Affiliated Hospital of Guangdong Medical University
Xinghua Lin
Affiliated Hospital of Guangdong Medical University
Wenfeng Zeng
Affiliated Hospital of Guangdong Medical University
Xingzhang Qin
Affiliated Hospital of Guangdong Medical University
Bailiang Miao
Affiliated Hospital of Guangdong Medical University
Sheng Gao
Affiliated Hospital of Guangdong Medical University
Jianjun Liu
Affiliated Hospital of Guangdong Medical University
Zhuo Li
Affiliated Hospital of Guangdong Medical University
Research Article
Keywords: berberine, renal cell carcinoma, ROS, DNA damage
Posted Date: February 22nd, 2023
DOI: https://doi.org/10.21203/rs.3.rs-1884385/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Read Full License
Page 1/18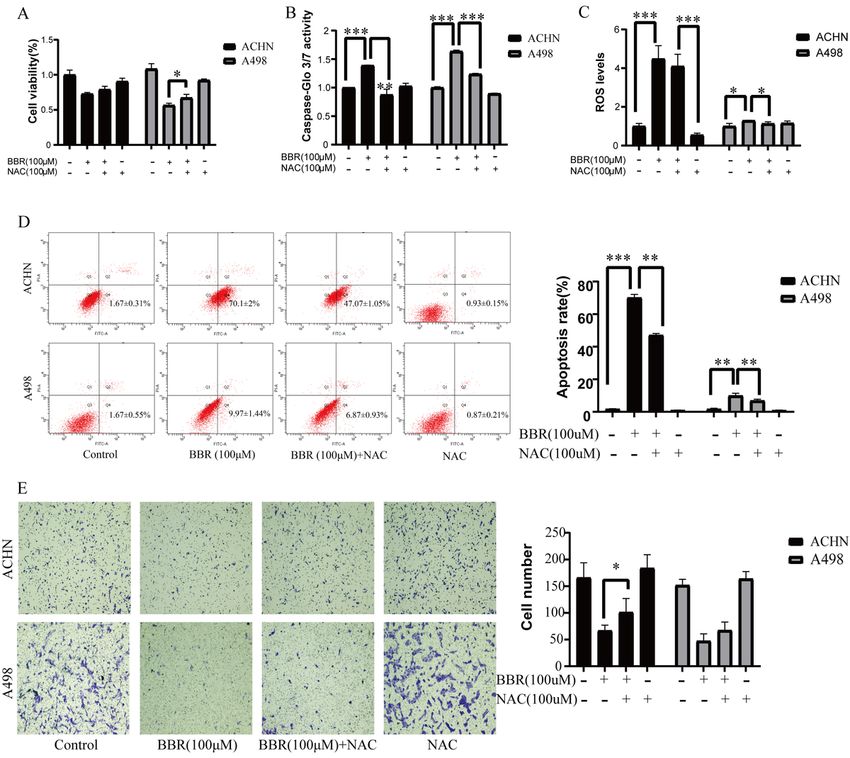
Abstract
Background Berberine is a natural isoquinoline alkaloid that has been shown to have antitumor properties
in a growing number of studies. However, its role in renal cell carcinoma remains unclear. This study
investigates berberine's effect and mechanism in renal cell carcinoma.
Methods The methyl-tetrazolium, colony formation, and lactate dehydrogenase assay were used to detect
proliferation and cytotoxicity, respectively. Flow cytometry, caspase-Glo 3/7 assay, and adenosine
triphosphate assay were used to detect apoptosis and the adenosine triphosphate levels. Wound healing
and transwell assay were used to examine the migration ability of renal cell carcinoma cells. Besides, the
level of reactive oxygen species (ROS) was explored using a DCFH-DA-based kit. Additionally, western
blot and Immunofluorescence assay was taken to determine the levels of relative proteins.
Results In vitro, our findings indicated that the proliferation and migration of renal cell carcinoma cells
treated with berberine in various concentrations were inhibited, while the level of ROS and apoptosis rate
were increased. Furthermore, The results of western blot showed that the expression of Bax, Bad, Bak,
Cyto c, Clv-Caspase 3, Clv-Caspase 9, E-cadherin, TIMP-1and γH2AX were up-regulated, while Bcl-2, N-
cadherin, Vimentin, Snail, Rad51 and PCNA were down-regulated after treating with berberine with various
concentration.
Conclusion The result of this study revealed that berberine inhibits renal cell carcinoma progression via
regulating ROS generation and inducing DNA break.
Introduction
Renal cell carcinoma (RCC), a group of cancers derived from renal tubular epithelial cells, accounts for
90% of renal malignancies and is the eighth most common cancer in men and the ten most common in
women[1-3]. Approximately 25-30% of patients present with various degrees of tumor invasion and
metastasis at the time of diagnosis. Metastatic RCC is among the most resistant malignancies, with an
average survival rate of less than a year [4]. A third of patients relapse and develop metastatic renal cell
carcinoma after surgery[5]. Chemotherapy and radiotherapy are currently the treatment options for
advanced RCC. However, their efficacy is not ideal due to RCC being insensitive to radiotherapy and
conventional chemotherapy drugs[6]. Therefore, it is urgent to discover new drugs with anti-renal cancer
effects.
Berberine is an isoquinoline alkaloid extracted from the herbal plant Rizoma Coptidis[7, 8]. It is well
known that berberine shows the effect on anti-diabetic, antimicrobial, anti-angiogenesis, anti-
inflammatory and cholesterol-lowering. However, more and more studies indicated that berberine has the
potential effect of anti-tumor in various cancer cells[9-11]. The function of cancer cells influenced by
berberine included proliferation, apoptosis, metastasis, invasion, autophagic cell death, and cell cycle
progression[12-15]. In mechanism, berberine-regulated signaling pathways included DNA damage and
Page 2/18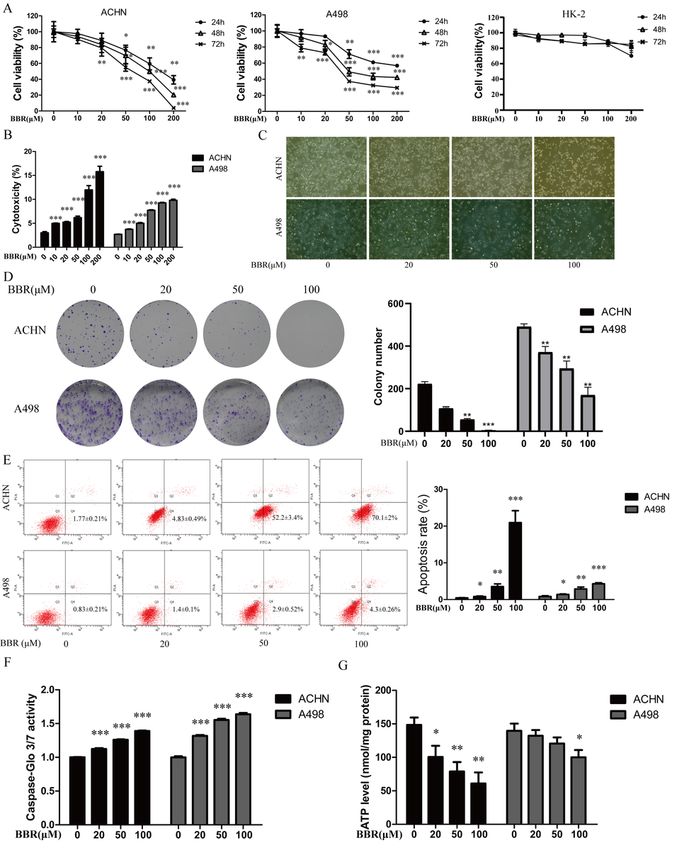
repair, ROS activation, Wnt/β-catenin, VEGFR2/ERK, epithelial-mesenchymal transition(EMT) pathways,
and so on[16-20]. Additionally, recent studies have shown that Berberine induces photoxicity in cancer cells
through ROS generation, mitochondrial dysfunction, and caspase activation[21].These studies showed
that berberine has a potentially anti-tumor effect via multiple mechanisms..
The production of ROS was from the reaction of electrons and oxygen, series of studies indicated that
ROS plays a crucial role in human health and disease[22]. The abnormal ROS level may promote cancer
progression via activating signaling pathways of the tumor microenvironment, DNA mutations,
metastasis, immune escape, angiogenesis, and extension of telomere[23]. In addition, the dysregulating of
DNA damage repair may regulate the progression of cancers through increasing mutagenesis and
genomic instability[24]. γH2AX, Rad51, and PCNA are the key molecules that can reflect the extent of DNA
damage and DNA repair[6].
This study explores how berberine affects proliferation, apoptosis, migration, and DNA damage of RCC
cell lines (ACHN and A498) and possible mechanisms. Our results indicated that berberine suppresses
proliferation, and migration and promotes apoptosis of RCC cells by increasing ROS generation and
inducing DNA breaks.
Materials And Methods
2.1 Cell culture and Reagents
Human RCC cells (ACHN and A498) and the normal renal cell line (HK-2) were obtained from Guangzhou
Jennio Biological Technology Co., Ltd. (Guangzhou, China). ACHN and HK-2 cells were grown in
Dulbecco’s modified Eagle medium (DMEM) (GIBCO, Thermo Fisher Scientific, Inc., Waltham, MA, USA).
A498 cells were cultured in RPMI 1640 medium (GIBCO, Thermo Fisher Scientific, Inc., Waltham, MA,
USA). All culture media were supplemented with 10% (v/v) fetal bovine serum (FBS; GIBCO, Thermo
Fisher Scientific, Inc., Waltham, MA, USA) at 37 °C in a humidified atmosphere that contained 5% CO2.
Berberine was purchased from Sigma (St. Louis, USA). DMSO was used to dissolve berberine to stock
solutions with a concentration of 100 mM. N-acetylcysteine (NAC) was purchased from the Beyotime
Institute of Biotechnology (Shanghai, China). PBS was used to dissolve them to 100 mM respectively.
These stock solutions were stored at -20 °C for experiment use.
2.2 Cell viability assay
Methyl-tetrazolium (MTT) assay was used to detect the viability of RCC cells. Cells were seeded in 96-well
plates (4 × 103 cell/well) (Nest Biotechnology, Wuxi, China), next day, various concentrations(0, 10, 20, 50,
100, 200 µM) of berberine were used to incubate these cells for 24 h, 48 h, and 72h, then, add 20 μL MTT
(Sigma Aldrich, St. Louis, MO, USA) in every which need to test. After 4 hours, a multiscan Ascent
microplate photometer (EnSpire 2300 Multilabel Reader, PE, USA) was used to measure the absorbance
of the solution at 492 nm.
Page 3/18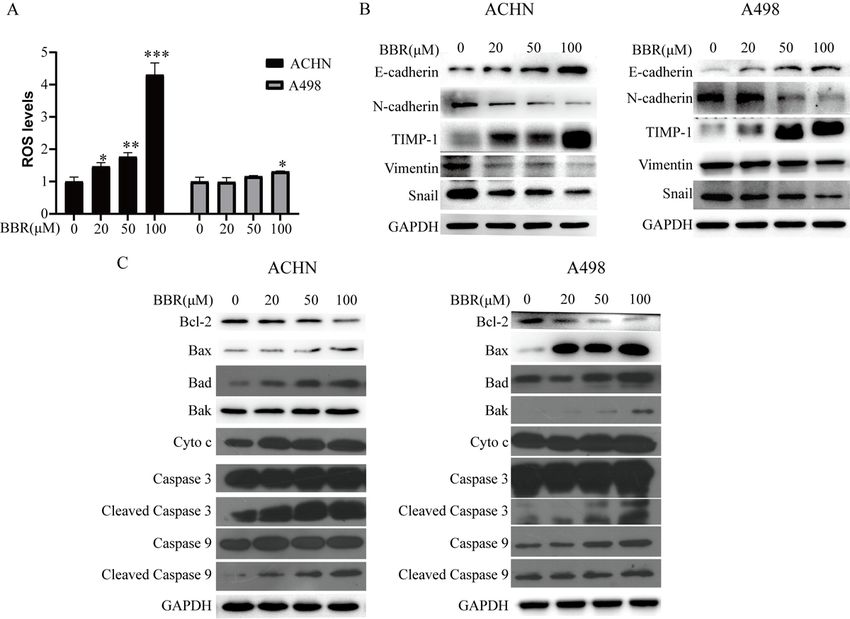
2.3 Colony formation assay
After being treated with various concentrations (0, 20, 50, 100 µM) of berberine for 24 h, ACHN and A498
cells were trypsinized and 2 × 103 cells were plated into 6-well plates (Nest Biotechnology) for another 2
weeks (ACHN) and 10 days (A498). The cells were stained with crystal violet (Beyotime, Shanghai, China)
for 20 min after being fixed with 4% paraformaldehyde for 15 min. Finally, the number of colonies formed
was counted.
2.4 Cytotoxicity assay
ACHN cells(4 × 103 cells/well) and A498 cells (4 × 103 cells/well) were seeded in 96-well plates and
treated with different BBR concentrations (0, 20, 50, 100 µM) for 24 h. After centrifuging at 400 × g for 5
min, the supernatants (120 μL/well) were transferred into new 96-well plates, and 60 μL of lactate
dehydrogenase(LDH) detection reagent was added to each well. The plates were incubated for 30 min at
room temperature in the dark, and the absorbance of the formazan was detected at 490 nm using a
reader (EnSpire 2300 Multilabel Reader, PE, USA).
2.5 Caspase-Glo 3/7 assays
ACHN cells(4 × 103 cells/well) and A498 cells (4 × 104 cells/well) were seeded in 96-well plates and
treated with BBR(0, 20, 50, 100 µM) and N-Acetyl-L-cysteine (NAC) (100 µM) for 24 h. Equal volumes (100
µL) medium and caspase-Glo 3/7 reagent (Promega Corporation, Madison, WI, USA) were added to each
well, and the cells were incubated at room temperature in the dark for 30 min. Luminescence was
measured by a luminometer (Berthold Sirius L; Titertek-Berthold, Pforzheim, Germany).
2.6 ATP levels
The adenosine triphosphate (ATP) levels were detected by the ATP Assay kit(Beyotime Institute of
Biotechnology) in ACHN and A498 cells. Cells (1 × 105 cells/well) were treated with different BBR
concentrations (0, 20, 50, 100 µM) for 12 h. Subsequently, cells were treated with 200 µL lysis buffer
(Beyotime Institute of Biotechnology) and collected by centrifugation at 12,000 × g for 5 min at 4 °C.
Subsequently, 50 µL supernatant and 100 µL ATP detection reagent were mixed. Firefly luciferase activity
was measured using a luminometer (Berthold Sirius L).
2.7 Apoptosis assay
The apoptotic rate of ACHN and A498 treated with various concentrations(0, 20, 50, 100 µM) of
berberine for 24 h were detected using the fluorescein isothiocyanate (FITC)-labeled Annexin V Apoptosis
Detection kit (BD Biosciences, Franklin Lakes, NJ, USA). Briefly, cells were digested with Trypsin Solution
without EDTA (Beyotime), then, washed and centrifugated according to the manufacturer’s instructions.
FITC-labeled annexin V and propidium iodide were used to stain cells for 15 min. The apoptotic death
was determined by annexin V/ PI staining. Then the apoptotic rate of cells was measured by Flow
Cytometer (BD FACSCelesta, Becton Dickinson USA) using flow cytometry BD FACSDiva 6.1 software.
Page 4/18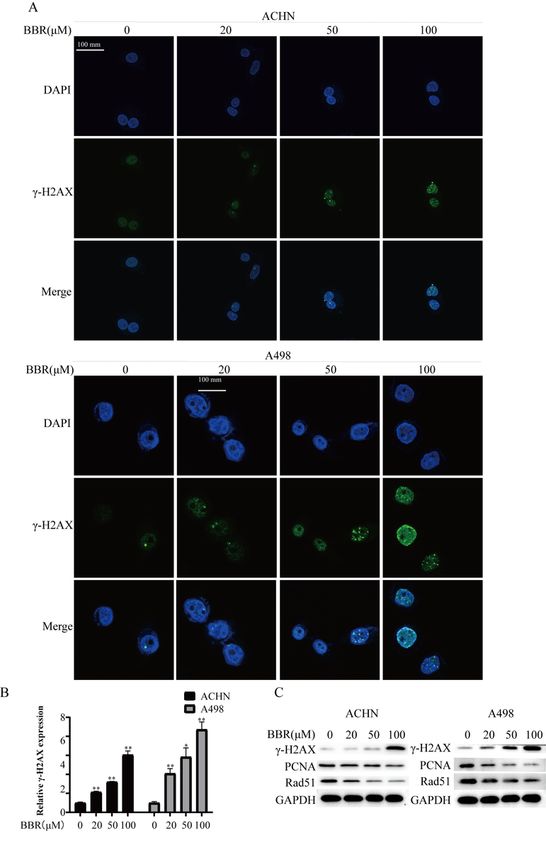
2.8 Wound healing assay
Cells were seeded into 6-well plates and scratches were made with a pipette tip after the confluence of
cells up to 100%. Then, washed cells with PBS, a medium without FBS were used to culture cells. After
being treated with various concentrations(0, 20, 50, 100 µM) of berberine for 24 h, the wound gaps and
cells were scanned under EVOS XL Core Imaging System (Life Technologies, USA). The areas of wounds
were assessed by ImageJ software. The wound healing rates were measured by ( ([original wound area of
drug group]-[wound area of drug group])/ ([original wound area of control group]-[wound area of control
group]) )× 100%.
2.9 Transwell assay
ACHN and A498 were harvested after being treated with various concentrations(0, 20, 50, 100 µM) of
berberine for 24 h. Then, the cells were resuspended with serum-free medium and then 200 μL cells
suspension(2 × 104 cells) were plated in the upper chamber of the insert (membrane pore size, 8 μm;
Corning) with Matrigel (BD Biosciences, Billerica, MA, USA). 750 μL medium with 20% FBS was added to
the bottom chambers. After incubating for 24 hours at 37°C, the cells were fixed with 4%
paraformaldehyde and stained with crystal violet. Cells adhering to the base membrane of the inserts
were scanned in 5 random fields with EVOS XL Core Imaging System. The migration cell numbers were
then counted with ImageJ software.
2.10 Intracellular ROS detection
The level of ROS in cells was explored using the Reactive Oxygen Species Assay Kit (Beyotime). Cells
were gathered and suspended with a medium (without FBS) that contained DCFH-DA (10 µM) after being
treated with various concentrations(0, 20, 50, 100 µM) of berberine. Then, cells were incubated at 37 °C
for 20 min in dark and shaken every 3-5 min. Serum-free mediums were used to wash cells three times to
clear intracellular DCFH-DA. The intensity of intracellular ROS was determined by the Fluorescence
intensity of DCF which was measured by Flow Cytometer (BD FACSCelesta, Becton Dickinson USA) using
flow cytometry BD FACSDiva 6.1 software.
2.11 Western blot analysis
Proteins were extracted from ACHN and A498 treated with various concentrations of berberine for 24 h.
In brief, cells were lysed in the mixture of RIPA Lysis Buffer (Beyotime) and PMSF
((phenylmethanesulfonyl fluoride) (Beyotime), and the concentrations of proteins were detected using
Bradford Protein Assay Kit (Beyotime, Shanghai, China). Then, the proteins were separated by SDS‐PAGE
(SDS-poly-acrylamide gel electrophoresis) and transferred onto PVDF (polyvinylidene fluoride
membranes) (EMD Millipore, Billerica, MA, USA). Next, 5% non-fat milk in TBST (Tris-buffered saline and
Tween 20) was used to block the membranes for 1 h, and the membranes were blotted with primary
antibodies at 4 °C overnight. Primary antibodies (dilution 1: 1,000) for Caspase-3 (#9665S), Caspase-9
(#9502S), Bad (#9239), Bax (#2772S), Bak (#12105S), Cyto-c(#4272S), Bcl-2 (#2876S), TIMP-1
Page 5/18(#8946S), E-cadherin (#3195S), Vimentin (#5741S), N-cadherin (#4061S), Snail (#3879S), γH2AX (#2577S), Rad51 (#8875S), PCNA (#2586S) (all Cell Signaling Technology, Inc., Danvers, MA, USA) were diluted with Primary Antibody Dilution Buffer (Beyotime). IgG-HRP secondary antibody (EarthOx, USA) was used to incubate the membranes for 1 h at room temperature. Proteins were observed in Tanon 5200 chemiluminescent imaging system (Shanghai, China). 2.12 Immunofluorescence analysis Cells were treated with various concentrations(0, 20, 50, 100 µM) of berberine for 48 h, then washed with PBS and fixed with 4% paraformaldehyde, Triton X-100 was used to incubate cells for 30 min, next, the cells were treated with PBS containing 1% BSA (Solarbio, Beijing, China) for 1 h. Cells were incubated with 1% BSA containing anti-γH2AX polyclonal Ab (dilution 1:100) (CST) at 4 °C overnight. In addition, Alexa Fluor 488-conjugated anti-rabbit IgG (dilution 1:100 ) (Thermo Fischer Scientific, Carlsbad, CA, USA) was used to treat the cells for 2 h at room temperature in dark. Then, cells were washed with PBS three times and stained with the Mounting Medium, antifading (including DAPI) for 3 min. Finally, cells were observed using a confocal laser scanning microscope. (Olympus FV3000, Tokyo, Japan). 2.13 Statistical analysis The analysis of all data was performed with SPSS 19.0 software (SPSS, Inc., Chicago, IL, USA). A one- way ANOVA test was used to determine the significance. *p
cells were increased in a concentrations-dependent manner (Fig.1E and F). It is known that apoptosis
could alter the intracellular levels of ATP. The ATP kit was used to examine ATP levels in ACHN and A498
cells after being treated with different BBR concentrations (0, 20, 50, 100 µM) for 12 h. It shows that BBR
could significantly decrease the ATP levels in ACHN cells, but the ATP levels on A498 cells only decreased
by BBR at 100µM (Fig.1G). These data indicated that berberine inhibited growth and promoted apoptosis
of ACHN and A498 in a concentration-dependent manner.
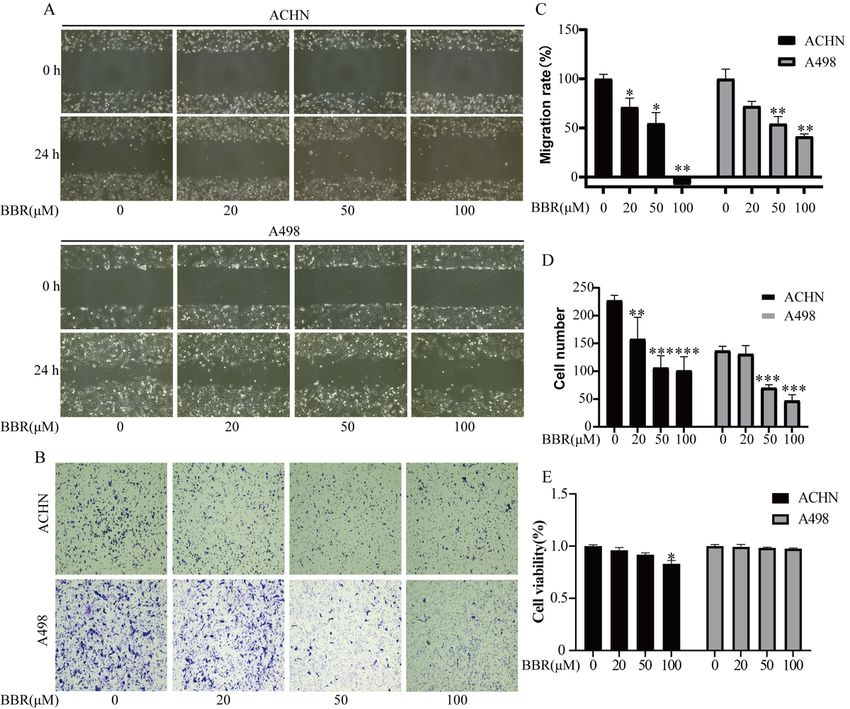
3.2 Berberine suppressed migration of RCC cells
The ability of migration of ACHN and A498 was determined by a wounding heal assay (Fig.2A、C) and
transwell assay (Fig.2B、D). The result showed that after being treated with various concentrations of
berberine (0, 20, 50, 100 μM), the migration of cells was suppressed gradually. A recent study reminds us
that cytotoxicity is minimal at high cell density in a medium that is low in serum[25]. Under this condition,
MTT assay was used to detect cell proliferation. The results show that Berberine(100 μM, 24 h) treatment
caused only little cell death in ACHN cells. More of all, the death of A498 cells is virtually nonexistent at
any concentration(Fig.2E). The above data suggest that Berberine treatment inhibits renal cancer cell
migration.
3.3 Berberine increased the level of ROS and regulated the expression of related molecules
The level of ROS was detected using the DCFH-DA-based kit. The result showed that the production of
ROS increased after being treated with various concentrations of berberine (0, 20, 50, 100 μM) (Fig.3A).
The apoptotic and metastasis related protein proteins were measured by western blot assay. The result
showed that the expression of Bax, Bad, Bak, Cyto c, Caspase 3, Caspase 9, E-cadherin, and TIMP-1 were
up-regulated, whereas Bcl-2, N-cadherin, Vimentin, Snail were down-regulated (Fig.3B and C). These data
revealed that berberine increased the production of ROS and regulated the expression of apoptotic and
metastasis related proteins.
3.4 NAC reversed the anti-tumor activity of berberine on the RCC cells
The effect of berberine regulating by NAC was explored after ACHN and A498 were treated with control,
berberine (100 μM), berberine (100 μM) + NAC (100 μM) and NAC (100 μM) for 24 h. The results showed
that the NAC reversed the effect of berberine on growth(Fig.4A), caspase-3/7 activity (Fig.4B), ROS levels
(Fig.4C), apoptosis(Fig.4D), and migration(Fig.4E). These data proved that NAC reversed the anti-tumor
effect of berberine in ACHN and A498. Berberine inhibit the proliferation, invasion, migration and
promotes apoptosis of renal cell carcinoma cells partly by promoting the generation of ROS.
3.5 Berberine increased DNA breaks and inhibited DNA repairs on the RCC cells
The expression of γH2AX (reflecting the level of DNA damage) was performed by the
immunofluorescence analysis assay and western blot assay. ACHN and A498 were treated with various
concentrations of berberine (0, 20, 50, 100 μM) for 24 h. The result showed that berberine-induced DNA
Page 7/18breaks in a concentration-dependent manner (Fig.5A and B). In addition, we also explored the expression
of PCNA and Rad51 (reflect the level of DNA repair). The data revealed that berberine decreased the
expression of PCNA and Rad51 (Fig.5C). These data showed that berberine aggravates DNA breaks and
inhibited DNA repair in the RCC cells.
Discussion
In recent years, the incidence of RCC is steadily rising at a rate of approximately 2–4% each year which
caused great harm to human health[26]. Patients with metastatic RCC show resistance to common
treatments of radiotherapy and chemotherapy, meanwhile, the effect of immunotherapy is poor[27]. It is
urgent to discover new effective drugs due to the existing treatment being poor for RCC. More and more
reports proved that berberine has an essential effect on anti-cancer[28]. Here, we investigated the effect of
berberine on RCC and its possible mechanism.
In this study, our data suggested that the growth and migration of RCC cells were significantly inhibited
after being treated with various concentrations of berberine, while the apoptosis of cells was intensified.
In addition, the activity of anti-tumor of berberine was reversed by NAC. Apoptosis, an ordered and
orchestrated cellular process, plays a key regulator in the human physiological and pathological
process[29]. The apoptosis process is usually induced by the extrinsic death receptor pathway and the
intrinsic mitochondrial apoptosis pathway[30]. The proteins of the B-cell lymphoma-2 (Bcl-2) family plays
cytochrome C (Cyto c) an essential role in the intrinsic mitochondrial apoptosis pathway[31], meanwhile,
caspase-9 is an initiator of the intrinsic mitochondrial apoptosis pathway, which then activates caspase-3
(executioner) and initiates a caspase cascade[30]. By contrast, NAC reduced the level of ROS which
reduced apoptosis significantly. In addition, the ability of cell migration and invasion is an important
indicator to evaluate the aggressiveness of solid cancer[32]. The expression of N-cadherin and TIMP-1
promotes metastasis and invasion in various cancers[33, 34], Snail induced migration of cancer cells via
the Epithelial-mesenchymal transition (EMT) pathway[35]. Besides, vimentin is required for the migration
of cancer cells that have undergone EMT[36]. In this study, we detected the expression of the above
proteins and proved that berberine regulated the biological behavior of RCC in a dose-dependent manner.
The mechanism of berberine inhibited growth and migration, but promoted apoptosis of RCC cells has
been investigated. We measured the level of ROS and proteins related DNA damage and DNA repair after
being treated with various concentrations of berberine. The result showed that with the rise of berberine
concentrations, the level of ROS was decreased. A previous study also showed that berberine increase the
level of ROS which is consistent with our results[37]. The generation of ROS induced intrinsic
mitochondrial apoptosis[38], the excess ROS cause an increase in mitochondrial membrane permeability
which further decreased the level of mitochondrial membrane potential and regulated the expression of
Bcl-2 family proteins. Then, induced the release of Cyto c. Finally, a caspase cascade has been
developed[30]. Elevated intracellular ROS level plays crucial roles in the regulation of Ethe MT process and
regulates the expression of related proteins[39]. In addition, the excessive generation of ROS can induce
Page 8/18DNA damage in cancer cells through compromise the DNA backbone[40, 41], γH2AX is an significant
molecular marker of DNA damage and aging whis ich associated with the level of DNA damage
positively[42], PCNA and Rad51 are important markers of DNA replication and repair[43, 44]. In this study,
we measured the expression of the above proteins, the data revealed that after being treated with various
concentrations of berberine, the expression of γH2AX was up-regulated, whereas PCNA and Rad51 were
down-regulated. Thus, these result suggested that berberine play its effect of anti-tumor on RCC via
producing ROS, inducing DNA damage, and inhibiting DNA repair.
In summary, this study investigated the effect of berberine in growth, apoptosis, and migration on RCC
cells and its possible mechanism. Our data showed that berberine inhibited growth and migration, but
promoted apoptosis of RCC cells via promoting the generation of ROS and inducing DNA damage. These
findings may provide a new treatment option for RCC.
Declarations
Funding
The present study was supported by the National Natural Science Foundation of China (grant no.
81272833).
Author contributions
Zhao Yuwan: Conceptualization, Methodology, Data Curation. Lin Xinghua: Validation, Writing - Original
Draft, Writing - Review & Editing. Zeng Wenfeng and Qin Xingzhang: Validation, Formal analysis.Miao
Bailiang: Investigation. Gao Sheng: Visualization. Liu Jianjun: Resources, Funding acquisition. Li Zhuo:
Project administration, Supervision.
Ethical standards
The manuscript does not contain clinical studies or patient data.
Conflict of interest
The authors declare that they have no conflict of interest.
References
1. Hsieh J J, Purdue M P, Signoretti S, et al. Renal cell carcinoma[J]. Nat Rev Dis Primers, 2017,3:17009.
2. Low G, Huang G, Fu W, et al. Review of renal cell carcinoma and its common subtypes in
radiology[J]. World J Radiol, 2016,8(5):484-500.
3. Doehn C, Grunwald V, Steiner T, et al. The Diagnosis, Treatment, and Follow-up of Renal Cell
Carcinoma[J]. Dtsch Arztebl Int, 2016,113(35-36):590-596.
Page 9/184. Gupta K, Miller J D, Li J Z, et al. Epidemiologic and socioeconomic burden of metastatic renal cell
carcinoma (mRCC): a literature review[J]. Cancer Treat Rev, 2008,34(3):193-205.
5. Bosma N A, Warkentin M T, Gan C L, et al. Efficacy and Safety of First-line Systemic Therapy for
Metastatic Renal Cell Carcinoma: A Systematic Review and Network Meta-analysis[J]. Eur Urol Open
Sci, 2022,37:14-26.
6. Zhao Y, Luo Q, Mo J, et al. Metformin in combination with JS-K inhibits growth of renal cell
carcinoma cells via reactive oxygen species activation and inducing DNA breaks[J]. J Cancer,
2020,11(13):3701-3712.
7. Li W, Hua B, Saud S M, et al. Berberine regulates AMP-activated protein kinase signaling pathways
and inhibits colon tumorigenesis in mice[J]. Mol Carcinog, 2015,54(10):1096-1109.
8. Pan Y, Shao D, Zhao Y, et al. Berberine Reverses Hypoxia-induced Chemoresistance in Breast Cancer
through the Inhibition of AMPK- HIF-1alpha[J]. Int J Biol Sci, 2017,13(6):794-803.
9. Okuno K, Garg R, Yuan Y C, et al. Berberine and Oligomeric Proanthocyanidins Exhibit Synergistic
Efficacy Through Regulation of PI3K-Akt Signaling Pathway in Colorectal Cancer[J]. Front Oncol,
2022,12:855860.
10. Xu M, Ren L, Fan J, et al. Berberine inhibits gastric cancer development and progression by regulating
the JAK2/STAT3 pathway and downregulating IL-6[J]. Life Sci, 2022,290:120266.
11. Zhong X D, Chen L J, Xu X Y, et al. Berberine as a potential agent for breast cancer therapy[J]. Front
Oncol, 2022,12:993775.
12. Nie Q, Peng W W, Wang Y, et al. beta-catenin correlates with the progression of colon cancers and
berberine inhibits the proliferation of colon cancer cells by regulating the beta-catenin signaling
pathway[J]. Gene, 2022,818:146207.
13. Yang L, Cao J, Wei J, et al. Antiproliferative activity of berberine in HepG2 cells via inducing
apoptosis and arresting cell cycle[J]. Food Funct, 2021,12(23):12115-12126.
14. Huang D, Wang W, Feng Z, et al. Berberine inhibits the invasion and metastasis of nasopharyngeal
carcinoma cells through Ezrin phosphorylation[J]. Zhong Nan Da Xue Xue Bao Yi Xue Ban,
2011,36(7):616-623.
15. Zhang Q, Wang X, Cao S, et al. Berberine represses human gastric cancer cell growth in vitro and in
vivo by inducing cytostatic autophagy via inhibition of MAPK/mTOR/p70S6K and Akt signaling
pathways[J]. Biomed Pharmacother, 2020,128:110245.
16. Hou D, Xu G, Zhang C, et al. Berberine induces oxidative DNA damage and impairs homologous
recombination repair in ovarian cancer cells to confer increased sensitivity to PARP inhibition[J]. Cell
Death Dis, 2017,8(10):e3070.
17. Chen Q, Hou Y, Li D, et al. Berberine induces non-small cell lung cancer apoptosis via the activation of
the ROS/ASK1/JNK pathway[J]. Ann Transl Med, 2022,10(8):485.
18. Li S Y, Shi C J, Fu W M, et al. Berberine inhibits tumour growth in vivo and in vitro through
suppressing the lincROR-Wnt/beta-catenin regulatory axis in colorectal cancer[J]. J Pharm
Pharmacol, 2022.
Page 10/1819. Jin F, Xie T, Huang X, et al. Berberine inhibits angiogenesis in glioblastoma xenografts by targeting
the VEGFR2/ERK pathway[J]. Pharm Biol, 2018,56(1):665-671.
20. Du H, Gu J, Peng Q, et al. Berberine Suppresses EMT in Liver and Gastric Carcinoma Cells through
Combination with TGFbetaR Regulating TGF-beta/Smad Pathway[J]. Oxid Med Cell Longev,
2021,2021:2337818.
21. Bhattacharyya R, Saha B, Tyagi M, et al. Differential modes of photosensitisation in cancer cells by
berberine and coralyne[J]. Free Radic Res, 2017,51(7-8):723-738.
22. Brieger K, Schiavone S, Miller F J, et al. Reactive oxygen species: from health to disease[J]. Swiss
Med Wkly, 2012,142:w13659.
23. Kirtonia A, Sethi G, Garg M. The multifaceted role of reactive oxygen species in tumorigenesis[J]. Cell
Mol Life Sci, 2020.
24. Chatterjee N, Walker G C. Mechanisms of DNA damage, repair, and mutagenesis[J]. Environ Mol
Mutagen, 2017,58(5):235-263.
25. Saha B, Pai G B, Subramanian M, et al. Resveratrol analogue, trans-4,4'-dihydroxystilbene (DHS),
inhibits melanoma tumor growth and suppresses its metastatic colonization in lungs[J]. Biomed
Pharmacother, 2018,107:1104-1114.
26. Chen S, Wang Q, Wang L, et al. REGgamma deficiency suppresses tumor progression via stabilizing
CK1epsilon in renal cell carcinoma[J]. Cell Death Dis, 2018,9(6):627.
27. Choueiri T K, Motzer R J. Systemic Therapy for Metastatic Renal-Cell Carcinoma[J]. N Engl J Med,
2017,376(4):354-366.
28. Rauf A, Abu-Izneid T, Khalil A A, et al. Berberine as a Potential Anticancer Agent: A Comprehensive
Review[J]. Molecules, 2021,26(23).
29. Wong R S. Apoptosis in cancer: from pathogenesis to treatment[J]. J Exp Clin Cancer Res,
2011,30:87.
30. Goldar S, Khaniani M S, Derakhshan S M, et al. Molecular mechanisms of apoptosis and roles in
cancer development and treatment[J]. Asian Pac J Cancer Prev, 2015,16(6):2129-2144.
31. Tomek M, Akiyama T, Dass C R. Role of Bcl-2 in tumour cell survival and implications for
pharmacotherapy[J]. J Pharm Pharmacol, 2012,64(12):1695-1702.
32. van de Merbel A F, van der Horst G, Buijs J T, et al. Protocols for Migration and Invasion Studies in
Prostate Cancer[J]. Methods Mol Biol, 2018,1786:67-79.
33. Cao Z Q, Wang Z, Leng P. Aberrant N-cadherin expression in cancer[J]. Biomed Pharmacother,
2019,118:109320.
34. Meng C, Yin X, Liu J, et al. TIMP-1 is a novel serum biomarker for the diagnosis of colorectal cancer:
A meta-analysis[J]. PLoS One, 2018,13(11):e207039.
35. Wang Y, Shi J, Chai K, et al. The Role of Snail in EMT and Tumorigenesis[J]. Curr Cancer Drug
Targets, 2013,13(9):963-972.
Page 11/1836. Usman S, Waseem N H, Nguyen T, et al. Vimentin Is at the Heart of Epithelial Mesenchymal
Transition (EMT) Mediated Metastasis[J]. Cancers (Basel), 2021,13(19).
37. Lopes T Z, de Moraes F R, Tedesco A C, et al. Berberine associated photodynamic therapy promotes
autophagy and apoptosis via ROS generation in renal carcinoma cells[J]. Biomed Pharmacother,
2020,123:109794.
38. Tripathi S K, Rengasamy K, Biswal B K. Plumbagin engenders apoptosis in lung cancer cells via
caspase-9 activation and targeting mitochondrial-mediated ROS induction[J]. Arch Pharm Res,
2020,43(2):242-256.
39. Chatterjee R, Chatterjee J. ROS and oncogenesis with special reference to EMT and stemness[J]. Eur
J Cell Biol, 2020,99(2-3):151073.
40. Yang H L, Thiyagarajan V, Shen P C, et al. Anti-EMT properties of CoQ0 attributed to
PI3K/AKT/NFKB/MMP-9 signaling pathway through ROS-mediated apoptosis[J]. J Exp Clin Cancer
Res, 2019,38(1):186.
41. Chatterjee N, Walker G C. Mechanisms of DNA damage, repair, and mutagenesis[J]. Environ Mol
Mutagen, 2017,58(5):235-263.
42. Siddiqui M S, Francois M, Fenech M F, et al. Persistent gammaH2AX: A promising molecular marker
of DNA damage and aging[J]. Mutat Res Rev Mutat Res, 2015,766:1-19.
43. Gonzalez-Magana A, Blanco F J. Human PCNA Structure, Function and Interactions[J]. Biomolecules,
2020,10(4).
44. Laurini E, Marson D, Fermeglia A, et al. Role of Rad51 and DNA repair in cancer: A molecular
perspective[J]. Pharmacol Ther, 2020,208:107492.
Figures
Page 12/18Figure 1
BBR inhibits cell proliferation and induces apoptosis in different renal cancer cell lines. (A): The effect of
BBR on the proliferation of human proximal tubule epithelial cells and renal cancer cells. ACHN、A498 and
HK-2 cells were treated with different BBR concentrations, and an MTT assay was used to determine cell
viability. (B): BBR increased cytotoxicity in RCC cells in a concentration-dependent manner. ACHN and
A498 cells were treated with different BBR concentrations for 24 h. Supernatants were harvested and
Page 13/18analyzed by lactate dehydrogenase kit. (C): ACHN and A498 cells were treated with different BBR
concentrations for 24 h, Besides, Morphologic changes were examined by phase contrast microscopy.
(D):BBR inhibits the clonogenic ability of different renal cancer cell lines. ACHN and A498 cells were
treated with different BBR concentrations for 24 h and then cultured with medium without BBR for
another 10 d. Cells were stained with crystal violet. (E): BBR induced apoptosis of ACHN and A498 cells in
a dose‐dependent manner. Cells were treated with BBR at the indicated concentration for 24 h, and cell
death was measured with flow cytometry. (F): BBR increase the activity of caspase-3/7.ACHN and A498
cells were treated with different BBR concentrations for 24 h. The apoptosis of cells was detected by
Caspase 3/7 activity assay. (G): BBR decreases ATP levels. ACHN and A498 cells were treated with
different BBR concentrations for 12 h. The ATP levels were detected by the ATP Assay kit. More than three
independent experiments were performed for each group, and the results are mean ± SD of three different
experiments. *** p-value < 0.001, ** p-value < 0.01, * p-value < 0.05. ns = nonsignificant.
Figure 2
Page 14/18Berberine suppressed migration of RCC cells. (A): ACHN and A498 cells were treated with different BBR
concentrations for 24 h. The inhibitory effect of BBR on renal cancer cell migration was detected using
wound healing assay in ACHN and A498 cells. (B): ACHN and A498 cells were treated with different BBR
concentrations for 24 h. The inhibitory effect of BBR on Renal cancer cell migration was detected by
Transwell chambers assay in ACHN and A498 cells. (C)The closure of area due to cell migration in Fig.2A
was quantified and shown. (D) The cells passing through membrane due to cell migration in Fig.2B were
counted and shown. (E) The ACHN and A498 cells at high cell density in low serum medium were treated
with different BBR concentrations for 24 h. MTT assay was used to determine cell viability. More than
three independent experiments were performed for each group, and the results are mean ± SD of three
different experiments. *** p-value < 0.001, ** p-value < 0.01, * p-value < 0.05. ns = nonsignificant.
Figure 3
BBR promotes reactive oxygen species generation and regulates the expression of related proteins. (A):
ACHN and A498 cells were treated with different BBR concentrations for 24 h, and ROS levels were
measured with flow cytometry. (B): The expression levels of E-cadherin、N-cadherin、TIMP-1、Vimentin and
Snail were determined by Western blotting analysis in ACHN and A498 cells after BBR treatment. (C): The
Page 15/18expression levels of Bcl-2、Bax、Bad、Bak、Cyto c、Caspase 3、Cleaved Caspase 3、Caspase 9 and Cleaved
Caspase 9 were determined by Western blotting analysis in ACHN and A498 cells after BBR treatment.
More than three independent experiments were performed for each group, and the results are mean ± SD
of three different experiments. *** p-value < 0.001, ** p-value < 0.01, * p-value < 0.05. ns = nonsignificant.
Figure 4
NAC reversed the anti-tumor activity of berberine on the RCC cells. ACHN and A498 were treated with
control, berberine (100 μM), berberine (100 μM) + NAC (100 μM) and NAC (100 μM) for 24 h. (A): The cell
viability was determined by MTT assay. (B): The caspase-Glo 3/7 activity was determined by Caspase-
Glo 3/7 assays. (C): The ROS levels was measured with flow cytometry. (D): The apotosis rate was
measured with flow cytometry. (E): The migration of Renal cancer cell was detected by Transwell
Page 16/18chambers assay. More than three independent experiments were performed for each group, and the
results are mean ± SD of three different experiments. *** p-value < 0.001, ** p-value < 0.01, * p-value <
0.05. ns = nonsignificant.
Figure 5
Page 17/18BBR induces DNA breaks and suppresses DNA repair. (A): Dose dependence course of γH2AX focus
formation. ACHN and A498 cells were treated with different BBR concentrations for 24 h, Besides, the
γH2AX focus were scanned by confocal microscopy. (B):Quantification plot of the immunofluorescence
intensity in Figure A. The immunofluorescence intensities were measured using the image J software,
Besides, The Immunofluorescence intensity ratio was normalized by comparing the data with the control
group and is represented as relative Immunofluorescence intensity. (C):The expression levels of γH2AX、
PCNA and Rad51 were determined by Western blotting analysis in ACHN and A498 cells after BBR
treatment. More than three independent experiments were performed for each group, and the results are
mean ± SD of three different experiments. *** p-value < 0.001, ** p-value < 0.01, * p-value < 0.05. ns =
nonsignificant.
Page 18/18You can also read

















































