A role for alternative splicing in circadian control of exocytosis and glucose homeostasis - Genes Dev
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
A role for alternative splicing in circadian
control of exocytosis and glucose
homeostasis
Biliana Marcheva,1,5 Mark Perelis,1,2,5 Benjamin J. Weidemann,1,5 Akihiko Taguchi,1 Haopeng Lin,3
Chiaki Omura,1 Yumiko Kobayashi,1 Marsha V. Newman,1 Eugene J. Wyatt,4 Elizabeth M. McNally,4
Jocelyn E. Manning Fox,3 Heekyung Hong,1 Archana Shankar,2 Emily C. Wheeler,2 Kathryn
Moynihan Ramsey,1 Patrick E. MacDonald,3 Gene W. Yeo,2 and Joseph Bass1
1
Department of Medicine, Division of Endocrinology, Metabolism, and Molecular Medicine, Northwestern University Feinberg
School of Medicine, Chicago, Illinois 60611, USA; 2Department of Cellular and Molecular Medicine, University of California at
San Diego, La Jolla, California 92093, USA; 3Department of Pharmacology, Alberta Diabetes Institute, University of Alberta,
Edmonton, Alberta T6G 2E1, Canada; 4Center for Genetic Medicine, Northwestern University, Chicago, Illinois 60611, USA
The circadian clock is encoded by a negative transcriptional feedback loop that coordinates physiology and behavior
through molecular programs that remain incompletely understood. Here, we reveal rhythmic genome-wide alter-
native splicing (AS) of pre-mRNAs encoding regulators of peptidergic secretion within pancreatic β cells that are
perturbed in Clock −/− and Bmal1 −/− β-cell lines. We show that the RNA-binding protein THRAP3 (thyroid hormone
receptor-associated protein 3) regulates circadian clock-dependent AS by binding to exons at coding sequences
flanking exons that are more frequently skipped in clock mutant β cells, including transcripts encoding Cask
(calcium/calmodulin-dependent serine protein kinase) and Madd (MAP kinase-activating death domain).
Depletion of THRAP3 restores expression of the long isoforms of Cask and Madd, and mimicking exon skipping in
these transcripts through antisense oligonucleotide delivery in wild-type islets reduces glucose-stimulated insulin
secretion. Finally, we identify shared networks of alternatively spliced exocytic genes from islets of rodent models of
diet-induced obesity that significantly overlap with clock mutants. Our results establish a role for pre-mRNA
alternative splicing in β-cell function across the sleep/wake cycle.
[Keywords: circadian clock; insulin secretion; exocytosis; alternative splicing; RNA sequencing; transcriptomics;
THRAP3; MADD; CASK; SNAP25]
Supplemental material is available for this article.
Received March 5, 2020; revised version accepted June 10, 2020.
The circadian clock is an endogenous timing system that Tong et al. 2010; Nakabayashi et al. 2013). RNA sequenc-
coordinates organismal physiology in anticipation of the ing analyses have shown that CLOCK/BMAL1 drive
sleep–wake cycle (Bass and Takahashi 2010; Asher and rhythmic expression of 10%–25% of all mRNAs within
Schibler 2011; Bass 2012). At the molecular level, the diverse mammalian tissues (Panda et al. 2002; Lowrey
clock is composed of basic helix–loop–helix transcription and Takahashi 2004; Hughes et al. 2009; Vollmers et al.
factors (TFs), in which activators in the forward limb 2009). Unique sets of genes oscillate across tissues due
(CLOCK and BMAL1) induce the transcription of their to interactions between CLOCK/BMAL1 and lineage-
own repressors (PER1/2/3 and CRY1/2) in a cycle that re- determining factors (Lee et al. 2005; Perelis et al. 2015).
peats itself every 24 h (Balsalobre et al. 1998; Preitner et al. Surprisingly, however, recent reports in liver have
2002; Liu et al. 2008; Bugge et al. 2012; Cho et al. 2012). indicated that ∼70% of genes that display mRNA rhyth-
The REV-ERB and ROR proteins form a reinforcing loop micity do not display oscillations in their corresponding
that modulates Bmal1 transcription. In turn, the core intron-containing pre-mRNAs (Koike et al. 2012). Addi-
TFs induce robust amplitude in the rhythmic production tionally, the zenith of CLOCK/BMAL1 chromatin binding
of downstream PAR-bZIP TFs in liver and β cells that play does not correspond to the peak phase of mRNA accumu-
key roles in mammalian metabolism (Gachon et al. 2006; lation for most target genes. These studies suggest that
posttranscriptional regulation is a crucial component of
5
These authors contributed equally to this work.
Corresponding author: j-bass@northwestern.edu
Article published online ahead of print. Article and publication date are
online at http://www.genesdev.org/cgi/doi/10.1101/gad.338178.120. Free- © 2020 Marcheva et al. This article, published in Genes & Development,
ly available online through the Genes & Development Open Access is available under a Creative Commons License (Attribution 4.0 Interna-
option. tional), as described at http://creativecommons.org/licenses/by/4.0/.
GENES & DEVELOPMENT 34:1–17 Published by Cold Spring Harbor Laboratory Press; ISSN 0890-9369/20; www.genesdev.org 1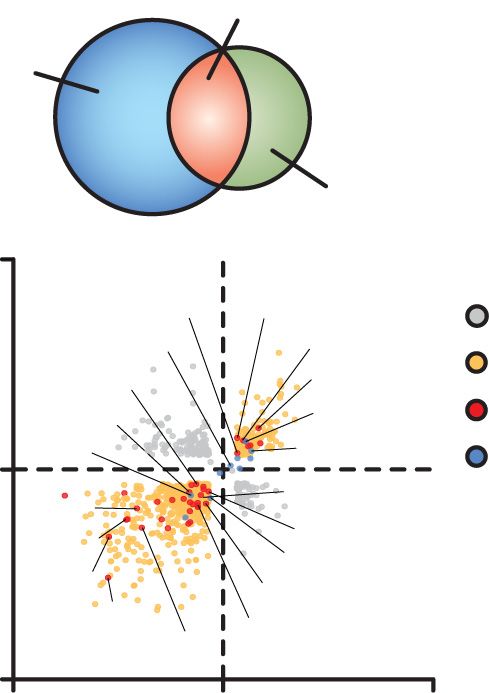
Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Marcheva et al.
circadian gene regulation (Koike et al. 2012; Menet et al. live cell capacitance measurements to determine the level
2012; Green 2018). of impairment in exocytosis in new CRISPR-generated
Mounting evidence suggests that posttranscriptional clock mutant β-cell lines as well as primary β cells from
RNA-processing events such as methylation, polyadeny- pancreas-specific Bmal1 −/− mice. Last, we identify an
lation, and alternative splicing (AS) are linked to the circa- overlap between AS events that are disrupted in clock mu-
dian clock (Kojima et al. 2012; McGlincy et al. 2012; tant β cells and those identified in diet-induced obesity.
Fustin et al. 2013). AS, which enables a single genomic lo- Thus, we propose that RNA missplicing in β cells contrib-
cus to produce multiple functionally distinct mRNAs in a utes to the etiology of diabetes mellitus in conditions of
tissue-, cell type-, and developmental stage-specific man- circadian disruption.
ner (Young et al. 1981; Vuong et al. 2016; Kim et al. 2018),
is initiated upon the recruitment of RNA-binding proteins
(RBPs) to consensus RNA motifs within regulatory ele- Results
ments of introns and exons of pre-mRNAs. RBPs then
Circadian control of alternative splicing in pancreatic
guide the oligomeric spliceosome complex to the correct
islets throughout the day
sites for subsequent inclusion, exclusion, or skipping of
exon cassettes to generate alternatively spliced tran- To determine whether posttranscriptional alternative pre-
scripts. Rhythmic production of unique alternative mRNA splicing in pancreatic islets is influenced by circadi-
mRNA splice isoforms from pre-mRNA has been shown an time, we first assessed genome-wide AS events from WT
to occur in plants, Drosophila, and mice (Sanchez et al. mouse islets cultured ex vivo and sampled in triplicate at 4-
2010; Hughes et al. 2012; McGlincy et al. 2012; Wang h intervals for 48 h following synchronization by a 1-h 10
et al. 2018). Oscillating AS events have also been shown μM forskolin shock (Perelis et al. 2015) (GSE69889) (see
to be driven by both the internal core circadian clock schematic in Fig. 1A). We used rMATS (replicate multivar-
and environmental timing cues such as temperature, iate analysis of transcript splicing) (Shen et al. 2014) to iden-
which drives the rhythmic phosphorylation and function tify and quantify the following types of AS events: skipped
of serine–arginine (SR)-containing spliceosomal proteins exons (SE), alternative 3′ and 5′ splice sites (A3SS and
as well as the splicing efficiency and stabilization of A5SS), and mutually exclusive exons (MXE) (Fig. 1A). To
cold-inducible RNA-binding protein (Cirp) (Morf et al. compare AS events across time, we first collected RNA se-
2012; Gotic et al. 2016; Preußner et al. 2017). quencing (RNA-seq) reads in our data that uniquely aligned
Evidence suggests that the clock regulates AS in neu- to exon–exon junctions annotated within the reference ge-
rons, where neurotransmitter release varies across the nome that were commonly assigned as AS events in all
day/night cycle (Shinohara et al. 2002; Barber et al. 2016; time points that had changes in percent-spliced in (ψ) values
Gizowski et al. 2016; Wang et al. 2018), raising the possi- reflecting exon inclusion in at least half of all averaged time
bility that rhythmic AS may play a role in physiology. Ge- points. The ensemble of AS events in our data included
netic studies show that selective ablation of the molecular 11,400 SEs, 2251 A3SSs, 1297 A5SSs, and 1689 MXEs across
clock within pancreatic β cells of adult mice induces dia- 6520 unique genes. To establish the trajectories of AS
betes due to severely impaired insulin secretion (Mar- events throughout the time series to assess rhythmic AS
cheva et al. 2010; Sadacca et al. 2011; Perelis et al. 2015; across the day, we calculated ψ for all common AS events
Petrenko et al. 2017). Clues to a role for rhythmic AS in from each of the three replicate samples for each time point
β-cell function originated in RNA and ChIP sequencing and identified statistically significant (adjusted P-value <
studies in mouse and human pancreatic islets, which re- 0.05) oscillations in ψ using JTK_cycle (Hughes et al.
vealed that as many as 80% of rhythmic mRNAs and 2010), allowing for period lengths of 20, 24, or 28 h. This
82% of mRNAs differentially expressed in Bmal1 −/− islets analysis revealed a total of 257 oscillatory AS events, con-
are not directly controlled by CLOCK/BMAL1, and are sisting of 172 SE, 44 A3SS, 20 A5SS, and 21 MXE events
therefore regulated either transcriptionally or posttran- across 241 unique genes (Fig. 1A; Supplemental Table S1),
scriptionally (Perelis et al. 2015). which collectively displayed a bimodal distribution in
Since the circadian transcriptome in islet cells is en- peak phases occurring ∼44 and 52 h after shock (Fig. 1B).
riched for mRNAs encoding synapse-associated proteins Sashimi plots, which show reads mapping across splice
that are established targets of AS, we sought here to test junctions to quantify exon inclusion levels, are shown
whether AS events are regulated by the clock through cir- across the circadian day to demonstrate rhythmic AS for
cadian RBP expression in islet cells. We used genome- Golgb1, which encodes a Golgi-associated and vesicle teth-
wide transcriptomics to quantify changes in AS events ering factor (Fig. 1C; Supplemental Fig. S1A). Rhythmic AS
and expression in circadian controlled RNAs. We found for other representative genes is shown by plotting inclu-
that wild-type (WT) islets display rhythmic expression sion level differences for cassette exons across the circadian
of RBPs and AS of mRNAs encoding protein regulators day in the splicing factor Srrm2 and the synaptic vesicle cy-
of peptidergic secretion, and that clock nullizygous β cells cle-associated GTPase Dnm1 (Supplemental Fig. S1A).
display significantly increased exon skipping events in These data establish for the first time that the molecular
genes that control insulin secretion. We report a novel clock regulates rhythmic AS throughout the day in the pan-
role for the circadian RBP THRAP3 in isoform selection creatic islet, a peripheral cell type.
within misspliced transcripts associated with insulin Given that the islet circadian transcriptome is enriched
secretion in clock-deficient cells. In addition, we used for mRNAs encoding trafficking and synapse-associated
2 GENES & DEVELOPMENT
Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Clock control of β-cell alternative splicing
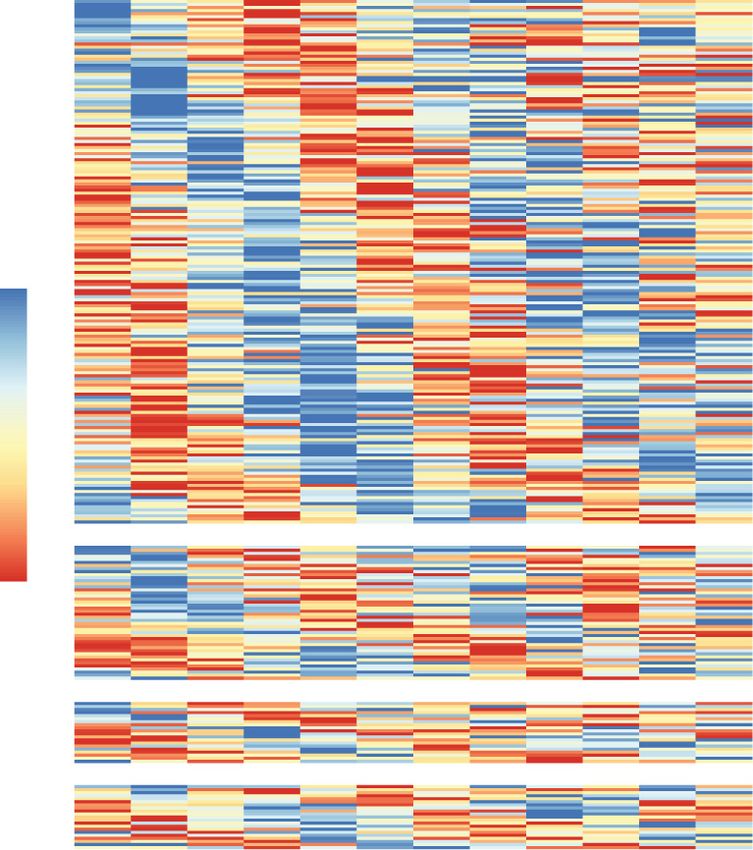
A B Figure 1. Circadian control of alternative splicing in
pancreatic islets throughout the day. (A) Schematic of
ex vivo experimental design for RNA isolation in for-
skolin-synchronized mouse islets. RNA was isolated
every 4 h for 48 h starting 40 h after forskolin shock.
TPS (hours), time after shock (hours) (n = 3/time-
point). Heatmap representing rhythmically spliced
genes (skipped exons [SE], alternative 3′ and 5′ splice
sites [A3SS and A5SS], and mutually exclusive exons
[MXE]) every 4 h over the course of 48 h in forskolin-
synchronized oscillating WT islets. (B) Radial histo-
D gram showing the number of splicing events of each
type with peak psi occurring within each period
length-adjusted phase, with the radius corresponding
to 40 splicing events. (C ) Representative sashimi
plots show time of day-dependent alternative splicing
of Golgb1 every 4 h across 24 h (starting 40 h after for-
skolin shock), with the exon inclusion level indicated
at each time point. (D) Pathway analyses reveal signif-
C icant enrichment of trafficking and secretion path-
ways within alternatively spliced gene sets. (E) Peak
phase in expression of cycling RNA-binding proteins
within the GO ontology term “RNA splicing.”
E
proteins known to be regulated by AS in neuronal tissues, mRNAs, we next interrogated the islet transcriptome
we hypothesized that similar pathways might emerge for rhythmically expressed RBPs and identified 296 oscil-
within circadian-controlled alternatively spliced tran- lating transcripts encoding RBPs (Supplemental Fig. S1C;
scripts. Indeed, interrogation of functional pathways in- Supplemental Table S2). We identified oscillation of RBPs
cluded in KEGG (Kyoto Encyclopedia of Genes and such as Nono and Adarb1, which have been implicated in
Genomes), GO (gene ontology), and reactome databases regulating circadian RNA transport and editing, respec-
using Metascape (Tripathi et al. 2015) revealed that genes tively (Terajima et al. 2017; Benegiamo et al. 2018). Of
with oscillatory splicing events are over-represented in note, of the 296 rhythmic RBPs, 99 have shown to been as-
trafficking and synapse pathways (Fig. 1D), including en- sociated with splicing (Fig. 1E; Supplemental Table S2),
docytosis (i.e., Dnm1, Pip5k1a, Vps25, Rab11a, and including Thrap3 (Lande-Diner et al. 2013). We further ob-
Smap2), spontaneous synaptic transmission (i.e., Rims2, served that rhythmic RBPs, including the splicing-associ-
Mtmr2, and App), and exocytic processes (i.e., Grik5, ated RBPs, displayed peak phases of expression ∼44–48 h
Snap47, Vps11, and Synj1). Several pathways and gene tar- after shock (Fig. 1E; Supplemental Fig. S1C), which over-
gets regulating transcription and posttranscriptional pro- lapped with the first peak in splicing (Fig. 1B) and the ze-
cessing were also identified (Fig. 1D), including the nith in insulin secretion (Perelis et al. 2015). Conversely,
transcript for splicing factor SRRM2 (Supplemental Fig. the trough of RBP expression (Fig. 1E; Supplemental Fig.
S1A). Interestingly, comparison of rhythmically spliced S1C) coincided with the second peak in splicing ∼52–
transcripts (from Fig. 1A) and rhythmically expressed 56 h after shock (Fig. 1B) and was in anticipation of the na-
transcripts (from Perelis et al. 2015) revealed that while dir in insulin secretion (Perelis et al. 2015).
only 34% of rhythmically spliced transcripts overlapped
with rhythmically expressed mRNAs, both were enriched
for gene sets important for synaptic and secretory path- Generation of cell models for analysis of BMAL1
ways (Supplemental Fig. S1B). and CLOCK alternative splicing and function
Since AS occurs as a consequence of interactions be-
Given that alternative splicing of mRNAs encoding key
tween RBPs and splicing regulatory elements in pre-
exocytotic and trafficking factors varies across the day in
GENES & DEVELOPMENT 3Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Marcheva et al.
islet cell clusters (Fig. 1D), and that both clock-controlled mental Fig. S3E). Together, these findings in vivo and in
expression and AS are cell type-specific (Young et al. 1981; cell lines are the first to show that both BMAL1 and
Vuong et al. 2016), we next sought to determine the specif- CLOCK TFs are required for appropriate control of stimu-
ic role of the β-cell molecular circadian clock in regulating lus-secretion coupling within β cells and prompted us to
the synthesis and processing of protein-coding messenger interrogate genome-wide transcription and processing of
RNAs. To do so, we first generated clonal isogenic β-cell mRNAs underlying the circadian control of insulin secre-
lines lacking a functional clock (Bmal1 −/− or Clock −/−) tion in pancreatic β cells versus in a heterogeneous islet
that recapitulate physiologic features of glucose-respon- population. Our results demonstrate a cell-autonomous
sive insulin secretion. We used CRISPR/Cas9 gene editing requirement for molecular clock components in β-cell
and homology-directed repair (HDR) in mouse β-TC6 cells function.
to establish Bmal1 and Clock nullizygous β cells harbor-
ing premature stop codons that lead to disruption of exons
BMAL1 and CLOCK control shared exocytic networks
encoding the bHLH DNA-binding domains (Supplemen-
regulated by alternative splicing
tal Fig. S2A). Quantitative real-time PCR and Western
blot screening of individual clones identified desired cell To first identify the repertoire of genes specifically regu-
lines lacking functional Bmal1 and Clock mRNA and pro- lated by both BMAL1 and CLOCK in β cells and to ulti-
tein, as well as decreased expression of their downstream mately determine whether circadian regulation of RNA
target Rev-erbα (Supplemental Fig. S2B,C). To confirm processing may also contribute to secretory defects in
functional loss of the β-cell core molecular clock, we clock mutant β cells, we initially performed RNA se-
transduced Bmal1 −/−, Clock −/−, and control β-cell lines quencing (RNA-seq) comparing polyadenylated mRNAs
with a lentivirus encoding a Per2 promoter fragment con- isolated from Bmal1 −/−, Clock −/−, and WT control β cells.
taining consensus binding sites for core circadian TFs im- We identified altered expression of 3035 genes in
mediately upstream of the firefly Luciferase (Luc) gene Bmal1 −/− cells and 1606 genes in Clock −/− cells (log2 FC
that is sufficient to drive circadian bioluminescence > 1, FDR < 0.01) (Fig. 2A). Among these differentially ex-
rhythms in mammalian cells (Supplemental Fig. S2D; pressed genes, we focused on the 815 mRNAs that were
Liu et al. 2008). Whereas control β-cell lines transduced differentially expressed in both Clock −/− and Bmal1 −/−
with the Per2-dLuc reporter displayed robust circadian cells, as they are more likely to underlie the common
bioluminescence rhythms following synchronization by secretory phenotypes observed in these two lines (Supple-
exposure to a 24-h temperature cycle that mimics the en- mental Table S3). Importantly, 79% (640/815) of the com-
dogenous body temperature rhythm (Buhr et al. 2010; Saini monly differentially expressed genes displayed common
et al. 2012), PER2-dLUC failed to oscillate in Bmal1 −/− and directionality (up-regulated or down-regulated in both),
Clock −/− β cells, indicating a loss of the core circadian net- including a large number of genes related to secretory pro-
work function in these cells (Supplemental Fig. S2D). Con- cesses (Fig. 2A), suggesting a high degree of overlap of tar-
sistent with β-cell-specific loss of clock function, these get genes regulated by both core clock activators, as
Bmal1 −/− and Clock −/− β-cell lines had impaired nutrient opposed to being CLOCK or BMAL1 targets independent
and nonnutrient secretagogue-induced insulin secretion of the core clock network. Among these commonly
following exposure to either glucose or potassium chlo- differentially expressed mRNAs, 21% (136/640) were up-
ride, respectively (Supplemental Fig. S2E; Marcheva et al. regulated while 79% (504/640) were down-regulated, con-
2010; Perelis et al. 2015). Importantly, these Bmal1 −/− sistent with inhibition of a transcription activation com-
and Clock −/− β-cell lines have secretory phenotypes that plex (Fig. 2A; Supplemental Fig. S5A). To identify the
are shared with pancreas-specific Bmal1 mutant mice functional β-cell gene networks regulated by both CLOCK
(Marcheva et al. 2010; Sadacca et al. 2011; Perelis et al. and BMAL1, we tested for over-representation of defined
2015). Here, we also show a similar loss of glucose-respon- KEGG, GO, and reactome pathways amongst the genes
sive insulin secretion in pancreas-specific Clock knockout that were commonly changed in both CLOCK and
mice. Ablation of CLOCK expression specifically in pan- BMAL1 mutant cell lines using Metascape (Tripathi
creatic islets was confirmed by immunofluorescent stain- et al. 2015). Consistent with inhibition of insulin secre-
ing (Supplemental Fig. S3A) and did not alter locomotor tion, the 504 down-regulated mRNAs common to both
activity, circadian period length, feeding rhythm, or body CLOCK and BMAL1 knockout cells were enriched for
weight (Supplemental Fig. S4). Similar to pancreas-specific genes with annotated functions related to G-protein and
loss of BMAL1, we observed significant hyperglycemia ion channel activation, synaptic processes, and regulation
without a concomitant increase in insulin (Supplemental of hormone secretion (Supplemental Fig. S5B). Among
Fig. S3B,C), and PdxCre;Clockflx/flx mice were glucose in- these were neurotransmitter and hormone receptors me-
tolerant and displayed reduced insulin secretion following diating responses to somatostatin (Sstr1, Sstr3, and
a glucose challenge compared with their littermate con- Sstr5), acetylcholine (Chrna3 and Chrna4), norepineph-
trols (Supplemental Fig. S3D). Finally, islets isolated rine (Adra2b), dopamine (Drd2), glycine (Glra1), and glu-
from pancreas-specific Clock KO mice also secreted signif- tamate (Gria4), as well as voltage-gated calcium (Cacng5
icantly less insulin in response to both glucose and KCl, an and Cacna2d4), sodium (Scn1b), and potassium channels
insulin secretagogue that causes direct β-cell depolariza- (Kcne1, Kcnh3, Kcnj12, Kcnk1, Kcnma1, and Kcnq2).
tion, despite no change in intracellular insulin content, mRNAs encoding proteins mediating the synthesis or
consistent with observations in CRISPR cell lines (Supple- downstream effectors of second messenger signals
4 GENES & DEVELOPMENTDownloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Clock control of β-cell alternative splicing
initiated by membrane-bound receptors such as calcium, vealing that exocytic genes in these two cell populations
3′ ,5′ cyclic adenosine monophosphate (cAMP), and diacyl within the islet oscillate in distinct phases (Petrenko
glycerol were also inhibited in clock mutant cells (Syt2, et al. 2017). To specifically test whether α- and β-cell types
Syt16, Adcy5, Rgs4, and Dgkb). Last, clock mutant cells display differences in peaks of AS events across the day, we
displayed altered expression of mRNAs encoding proteins examined rhythmic patterns of AS events in FACS-sorted
regulating organelle localization (Rab15 and Myrip) and α and β cells collected every 4 h across a 24-h circadian cy-
synaptic vesicle priming and exocytosis (Doc2b and cle by performing rMATS analyses on this published data
Cadps2). These changes in the transcriptomes of CLOCK- set (GSE95156), as we did with the synchronized islets in
and BMAL1-deficient cells suggest altered expression and Figure 1A. Indeed, we found that a majority of the tran-
function of proteins coupling nutrient sensing to the reg- scripts that displayed altered splicing in the clock-defi-
ulated release of insulin. Indeed, electrophysiologic analy- cient β cells (Fig. 2D), but that were not identified as
ses revealed that Bmal1 mutant β-cell lines and primary statistically rhythmic in whole islets, exhibited peak
mouse β cells from PdxCre;Bmal1flx/flx mice display re- exon inclusion at distinct times of day in α versus β cells
duced rates of exocytosis following direct depolarization (Fig. 2E). To independently determine whether the differ-
as indicated by reduced capacitance in the circadian mu- entially spliced transcripts identified in the clock mutant
tant β-cell lines and mouse β cells (Fig. 2B). The set of com- β-cell lines (Fig. 2D) displayed rhythmicity in a pure WT
monly differentially expressed genes also included β-cell population, we synchronized WT β-TC6 cells (Sup-
mRNAs encoding synaptic membrane-associated pro- plemental Fig. S5E) and performed targeted qPCR analysis
teins that are functionally regulated by AS in neurons of representative differentially spliced (SE) transcripts, in-
(Snap25, Nrxn2, and Erc2), which prompted us to next in- cluding the secretion-related genes Cask, Madd, Dock11,
terrogate the impact of disruption of the molecular clock and Dctn1. We observed patterns of rhythmic exon inclu-
on posttranscriptional pre-mRNA AS in β cells. sion in these key targets, which exhibit high degrees of AS
Given that synchronized WT islets display rhythmic AS under control of CLOCK/BMAL1 (Fig. 2F; Supplemental
(Fig. 1) and that clock control of peptide secretion in pan- Fig. S5F), indicating that splicing events that are perturbed
creatic β cells is similar to that in neurons, we next as- by clock mutation display rhythms in a pure population of
sessed genome-wide AS patterns in Clock and Bmal1 β cells. Together these data reveal a connection between
mutant β cells. We hypothesized that circadian control the activating limb circadian TFs and rhythmic splicing
of RNA processing may also contribute to cell type-specif- in pancreatic β cells.
ic circadian gene regulation and insulin secretion in the β
cell. We used rMATS to quantify AS events in Clock and
Thyroid hormone receptor associated protein 3
Bmal1 mutant cell lines relative to WT. Considering both
(THRAP3) regulates circadian clock-dependent
intron spanning and exonic reads, we detected 1453 AS
alternative splicing
events in Bmal1 mutant cells (FDR < 0.05, ψ > 0.1) consist-
ing of 1018 skipped exons (SE), 174 mutually exclusive ex- Given our observation that rhythmic splicing occurs un-
ons (MXE), 134 alternative 3′ splice sites (A3SS), and 127 der the control of core circadian transcription factors
alternative 5′ splice sites (A5SS). In Clock mutant cells, (TFs), we next sought to examine how the clock TFs regu-
we detected 710 AS events, of which 492 were SE, 105 late AS in pancreatic β cells by examining RBPs displaying
were MXE, 63 were A3SS, and 50 were A5SS events. Apply- rhythmic expression in islet cells (Supplemental Table S2).
ing the same filtering approach as for mRNA expression, Identification of rhythmic expression of the gene encoding
we identified 101 AS events within 95 annotated genes the RBP THRAP3 (thyroid hormone receptor-associated
that were common and displayed similar directionality protein 3) (Supplemental Fig. S6A ) was of particular inter-
in both Bmal1 −/− and Clock −/− cells (Fig. 2C,D; Supple- est since THRAP3 has been shown to regulate AS by inter-
mental Table S4). A majority of the common AS events acting with splicing factors (Nakatsuru et al. 1989; Yarosh
were SE (93), with the remaining as MXE (four) and A3SS et al. 2015). It also binds CLOCK/BMAL1 to regulate
(four) events (Fig. 2C). Only four of these 95 alternatively rhythmic transcription (Lande-Diner et al. 2013) and met-
spliced genes were included among the 640 differentially abolic TFs such as PPARγ recruit THRAP3 to chromatin
expressed mRNAs (Supplemental Fig. S5D), suggesting (Choi et al. 2014). To determine whether THRAP3 binds
that changes in AS and mRNA abundance are regulated to differentially spliced transcripts in clock mutant β-cell
by distinct mechanisms downstream from the molecular lines, we performed enhanced UV cross-linking and im-
clock. Differentially spliced mRNAs encoded proteins munoprecipitation (eCLIP), followed by sequencing analy-
known to mediate cellular events important for insulin sis (Van Nostrand et al. 2016) in two replicate WT and
vesicle transport and exocytosis, such as cytoskeletal Bmal1 −/− lysates. Briefly, THRAP3–RNA complexes
and motor proteins (Mapt, Arhgap21, Myo5a, Kif16b, were immunoprecipitated, and ribonucleoprotein (RNP)
and Dctn1), vesicle membrane constituents (Erc2, Ap1s2, complexes within the expected size range were isolated
and Sec31a), synaptic vesicle-associated proteins following electrophoresis and transfer to nitrocellulose
(Snap25), and kinases and GTPases responsible for post- membranes. For each IP replicate, a corresponding input
translational modifications to the secretory apparatus lysate was processed to isolate a size-matched input
(Cask and Madd) (Fig. 2D; Supplemental Fig. S5C). RNA sample, comprising RNAs within the background
Previous studies have used FACS-sorting to uncover dis- set of RNPs similarly sized to THRAP3 complexes. We
tinct circadian transcriptional profiles of α and β cells, re- note that size matching to input is necessary in order to
GENES & DEVELOPMENT 5Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Marcheva et al.
control for differences in RNA expression levels across the pared with input can also be visualized within individual
genome. Illumina sequencing libraries were then generat- genes containing clock-dependent SE events, including
ed from both WT and Bmal1 −/− β-cell-derived IP and input Cask, Madd, and Dctn1 (Fig. 3B; Supplemental Fig. S6C).
RNA (Fig. 3A, see Materials and Methods). This approach Given that THRAP3 displays increased binding at exons
allowed us to compare abundance of THRAP3–RNA com- flanking differentially spliced cassette exons in Bmal1 −/−
plexes relative to size-matched protein-bound RNA input β cells, we hypothesized that siRNA-mediated depletion
within each genotype, thus controlling for alterations in of Thrap3 in Bmal1 −/− β cells would lead to increased in-
total protein-bound RNA abundance that may occur be- clusion of the skipped exons. Indeed, we observed in-
tween genotypes. We found that THRAP3 binding in the creased retention of skipped cassette exons in Madd,
Bmal1 −/− cells was enriched relative to Bmal1 −/− size- Cask, and Dock11 by ∼1.5-fold in Bmal1 −/− cells express-
matched input specifically within coding exons flanking ing Thrap3 siRNA (Fig. 3C; Supplemental Fig. S6D), with-
exons that were skipped more frequently in Bmal1 −/− cells out altering cassette exon inclusion in WT cells
(Fig. 3A), but not in regions surrounding exons that were (Supplemental Fig. S6E). However, THRAP3 depletion
preferentially retained in clock mutant cells (Supplemen- did not restore inclusion of the cassette skipped in
tal Fig. S6B), as indicated by increased normalized se- Dctn1, which may be regulated by other clock-controlled
quencing reads in THRAP3 IP versus size-matched input RBPs (Supplemental Fig. S5G). This data indicates that in-
control at the exons immediately up-stream of and creased THRAP3 occupancy at these RNAs in Bmal1 −/−
down-stream from the skipped exon (Fig. 3A). In contrast, cells drives exon skipping.
we did not observe enrichment of THRAP3 IP sequencing Having established a role for THRAP3 in AS in Bmal1 −/−
reads relative to input in WT cells in these regions (Fig. 3A), cells, we next hypothesized that THRAP3 may contribute
suggesting that THRAP3 displays increased occupancy to the rhythmic AS in WT islets. To test whether THRAP3
along these regions in Bmal1 −/− cells compared with was enriched within exons flanking rhythmic cassette ex-
WT. This pattern of enrichment of THRAP3 IP reads com- ons in a time of day-dependent manner, we examined SE
A B Figure 2. Ablation of both BMAL1 and CLOCK dis-
Overlap of differentially Reduced exocytosis in rupts splicing within gene networks important in ves-
expressed genes in clock mutant β-cell lines
Bmal1-/- and Clock -/- β-cells and primary β-cells icle trafficking and exocytosis. (A, top) Venn diagram
of overlapping differentially expressed genes identi-
Bmal1-/- / WT 815 10 β-cell line
2220
WT fied by RNA sequencing in Bmal1 −/− and Clock −/−
Cumulative capacitance
Bmal1 -/-
-/-
Clock / WT 5 β-cell lines compared with control cell line. (Bottom)
791
15
Scatter plot depicts all differentially expressed genes,
(fF/PF)
Erc2 Rab15 0 highlighting secretion and circadian genes with a
Log2 fold change
Dbp Doc2b All genes
(Clock -/- / WT)
Similarly dysregulated 20 Mouse β-cells
Snap25 Cadps2
Adra2b Rorc Secretion Bmal1flx/flx strong correlation between directionality of gene ex-
pression changes between Bmal1 −/− and Clock −/− β-
Per3 Cry1 Circadian
0 PdxCre; Bmal1flx/flx
Chrna3
Sstr1 Cacng5 10
Glra1
Myrip
Rev-erbα
Syt2
cell lines. (B) Capacitance measurements in
-15
Kcnj12
Scn1b
Nrxn2 0 Bmal1 −/− β-cell lines (n = 13–17 cells per genotype)
0 2 4 6 8 10
-15 0 15 Depolarization number and β cells from PdxCre;Bmal1flx/flx mouse islets (n
Log2 fold change
(Bmal1-/- / WT ) = 4–5 mice per genotype, 7–16 cells per mouse) com-
pared with controls. (C ) Splicing analysis reveals dif-
C D
Disrupted splicing in Bmal1-/- Overlap in splicing events ferential alternative splicing events (skipped exons
and Clock -/- β-cells in Bmal1-/- and Clock -/- β-cells [SE], mutually exclusive exons [MXE], and alternative
3′ splice sites [A3SS]) common in both Bmal1 −/− and
SE (93)
1
A3SS (4) SE
Clock −/− β-cell lines. Representative genes are listed
Myo5a
Δψ Clock -/- / WT
(% spliced in)
Dock11 MXE
Madd Snap25
Dctn1 A3SS
Cask
Dock11
Pfkfb1 Madd below each category. (D) Scatter plot depicts alterna-
0
tively spliced genes common to both Bmal1 −/− and
Snrpn Erc2
Dctn1 Grina Mapt
Nudt13 Sec31a
Clock −/− β-cell lines, highlighting strong correlation
MXE (4) Zfyve21
Nfat5 Ap1s2 Arhgap21
Zfp945 Cask
Fkbp14
Asxl2 Snap25
-1
Kif16b
of directionality between Bmal1 −/− and Clock −/− β-
-1 0 1
Mapt
Creb1
Slc7a2
Ints10 Δψ Bmal1-/- / WT cell lines. (E) Zeitgeber time (ZT) of peak inclusion
Pigq (% spliced in) in sorted α and β cells from islets (left) (Petrenko
et al. 2017) and patterns of mean ψ scores across two
E F
Distinct peak phases of clock- Rhythmic splicing of events 24-h timescales for skipped exon events identified
dependent alternative splicing identified in clock mutant in clock mutants in synchronized islets (right). (F)
events in α vs β cells β-cells
Cell type Synchronized
Rhythmic splicing events identified via qPCR in for-
α β islets 1.05 Cask SE
WT skolin-synchronized WT and Bmal1 −/− β-cell lines
Relative mRNA expression
0
(skipped / upstream exon)
Bmal1-/-
Peak inclusion
(n = 3/timepoint). RNA was isolated every 4 h across
24 h (starting 24 h after forskolin shock). Skipped
(ZT)
0.6 P=1.60e-01
0.15
0 exon expression was normalized to a neighboring
20 0.3 Madd SE (nonspliced) exon, and evaluation for rhythmicity
max
performed by JTK_CYCLE (adjusted P-value shown).
Inclusion
0.15
level
P=4.12e-03
0.05
0
min Petrenko 40 48 56 64 72 80 24 28 32 36 40 44
et al Time post shock (hrs) Time post shock (hrs)
6 GENES & DEVELOPMENTDownloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Clock control of β-cell alternative splicing
A Increased THRAP3 binding to exons D Enrichment of common TF motifs among Figure 3. THRAP3 mediates clock-dependent alter-
flanking SE events in Bmal1-/- cells promoter regions of THRAP3 ChIP-seq native splicing. (A) Schematic of eCLIP sequencing
target and alternatively spliced genes
Crosslinking &
fragmentation
Immunoprecipitation High throuhgput
sequencing experiment (top) and normalized (reads per million)
eCLIP-seq
THRAP3
THRAP3
antibody
ACUG
THRAP3
targets
Not THRAP3
targets
THRAP3 eCLIP or input sequencing reads along indi-
mRNA
RNA
THRAP3
RNA cated regions of alternatively spliced mRNAs (bot-
UV + RNAase binding site target sites 533 409
tom). Solid line indicates mean for two replicates
Bmal1-/- AS genes and shaded area represents 95% confidence interval.
(B) UCSC genome browser tracks showing reads
Mean coverage (RPM)
Upstream Upstream Skipped Downstream Downstream
exon intron exon intron exon TF Bmal1-/- Oscillating THRAP3 Consensus
2
Bmal1-/-
motif AS AS enriched sequence
promoter promoter promoter from indicated THRAP3 IP and input samples along
USF2 GT
CACGTGGT regions within Cask and Madd RefSeq transcripts.
bHLH
AC
A G
AA TG
0
C T
C
T A
GT A
G
CG
T A C
T C
G CT A CA
2 TFE3 AT
G
CC
G
T
CACGTGAC
AG
TA
T
G
CG
A C
T
G
C
AC
TA TA
TA G
C G
CC
TA
TAG
TG
Skipped exons identified by rMATS and flanking ex-
E-BOX CCG
GGCGTCACGTGA G
WT
TTCC
T
A AAA TAGT AG CT
G T
G A C
T C
G A
A CT A C T
ZNF143ISTA ATTT
CA
GCGA
TGACCCAG AGTGC C C
GGT A GT A GT A GA
CT
C
TG
CG
A
TT CCA
AT
C
C T
A GT A G
ons are shaded in gray and track heights for each sam-
YY1 CAAGATGGCGGC
0 ple are standardized to equal height for row shown.
G C
ATGT
C G
CT TGT
C
A G A
C CT A CT A GT
A
C
A
TATCTG
A
0 100 SP5 G
C
GGGGGCGGAGC
A T
TA
CAA TC CT CT A CT A G
T
A CT A C A T G
CTT C
A
G
T
A
Distance in bins SP C
G CCGCCCCC
G CC
G A
TATAA CT AG
T GT GT A CT AGT A GT A G A
A
TT G
T
G A
(C ) Thrap3 siRNA knockdown restores exon inclu-
AACTACAA TCCCAG TGC
AG
G C A
RONIN C
sion in Bmal1 −/− β cells, assessed by qPCR (n = 3).
TT
THRAP3 THRAP3
C
T
CTGT
C GT A CGT
G A
C GT A
GGG
T
C
A
AC C GGT
A GT A GT
A CT GCGGAT ATCCT A
T
A
G
Input Input MAZ GGGGGGGG
CT ACT A
T AC
CTCAA
AA
C T T CT CT A
KLF9 GCCACACCCACT
G C
(D, top) Pie chart summarizing number of AS genes
C
TT
AA
GT A
G C
G
T GT A C TGT A GT A GT
A G
C TG
AG T A
ZF KLF6 CT
T
GGGCTGTGGCC
AG
A G T
T
A
in Bmal1 −/− β cells identified as THRAP3 ChIP-seq
G C CT A CT A CT A G A
CT A C
A C A TCTG AG A
GGGCTGGTGGC
A
B Increased THRAP3 binding to exons KLF5
KLF4
C
T
GA CT
GCCACACCCA
CT A CT A G A A
CTCCT
AACG
TT
A
A
flanking SE events in Cask and Madd A T
targets. (Bottom) Bubble plot indicating enrichment
CT
T
GGT A
A C
G
T GT A G
CT GT A GT A GT A C
G
GCCC
TG
ACGCCC
A TCGC
in Bmal1-/- β-cells KLF3 CA TG
TC
CG
A T
C
GT CT A GT A G A T GT GT A CT GT A GT A GT A G AAAA
KLF14 GGGGGCGG
G T
TG
GC
of indicated motifs within promoter regions of AS
TA
A T ACT
AG
CCA
T CTA
C CT A CT A G A CT AC
A C
TTA
exon 11 exon 26 KLF10 GGGGGTGTGTCC T C GGT
Cask Madd
A TG
5 kb 1 kb
CAT
TAT C C C A CT A G A CT A G
C A CT A C AAA
Long
EGR2 GGCGTGGGCGG
T
CCT
AAA
TGCGTGGGC
TGCT A GA
C A CT CT A CT A G
T
A CT
A C
TC
T
A
G
A
genes and promoter-localized THRAP3 peaks. Size
Short EGR1 T
G T
of bubbles reflects enrichment score for motifs within
G
CA
C
A T G
A
CT A G
C A CT ACT
A CT A GCT A A
GABPA GG
CCGA
AA
CCGGAAGT AC
THRAP3 1 TTA G AC TT
CG
T
T
GC
T
A
TC
G
CT A
TTCCG
Bmal1-/-
CAC
T GC
T
FLI1
each indicated set of promoters, and those called as
GGTA
TG TC A
TG
THRAP3 2 AC AGAT
C A
G
AA
G
CG T
A
C
C
A
ACAGGAAGT
AC
Input 1 EWS:FLI1 GG
TT C GT A C
GT CT A CT A GT
C GT
C CT G A
TTCCG
TGTT
statistically significant (Homer hypergeometric test
CCAC
GTG
CC
G
Input 2 ETV2 T T
AAC
G
GA
T AG
C G A
C GT A GT A
C
A
AC TGA
A
AGGAAG
CC
AA T
THRAP3 1 ETV1 G
CG
TG AG C
P < 0.05) are shown in blue.
TC
A TCA
ETS
A G T
TG
T
A
C
TC
T
G
CCT
GGAAGTG
ACA
WT
THRAP3 2 ETS1 GGC
C
TA T G
T CT A CT A GT
C G
C
C
A
TACT
CT G A
Input 1 ETS
AAC
CG
G CGGAAGT
TCGA TA
T GTCT ACT AC
GT C
G TC
T G
AAC
Input 2
ERG
ACA
G
CG
TAGGAAGTG T C
GT CT A CT A GT
C G
C TCT ACC
T
G
A
A
Upstream Skipped Downstream Upstream Skipped Downstream
ELK4
CAC
T
TTCCGG
GTGA
AG
C
TAG
C G
C AG AGT A
T
C
ATT
A
CG
C
T
A
ELK1
CAC
TG
TTCCGG
ACT
GTGA AG
C
G
C
T
G
AG
TA C
A
A
CA
TT
AG
C
T
ELF1
A C
GC
TG CGGAAGT
CTGA
A
T
A
G
TC
TAC
TT
CC
GT
AGT CA
G
A
C
C Thrap3 knockdown restores Cask CRE G
T
A T ACGTCAC
CGG G
TC
C AA TC
G A TA C G
CT GT
A CT AC
G A TA G G
CT A
G T
ATGACGTCATC
and Madd splicing in Bmal1-/- β-cells
C
T
c-JUN-CRE G GC
TA CTA
GGG
bZIP
CTC A
G C A TG
CTATC
G A GT AGT
C A A
A
TGACGTCAT
CG
GAG C
ATF7 AT TAA
expression (skipped /
TCC
T C
A
GA CCAT
TG
GT
G
T
GCC
TA
C
A G G
TGACGTCAT
CGA
GAG
2 ATF2 TCC
AT TGGCAAT
T
TAC C
G
upstream exon)
A
T AT G A
Relative mRNA
G
CCC
GA GTC
*** GA
TGACGTCA
*** ***
ATF1
C
TC
CG
AG
TA
G
C
A
GTTGCCATGGCAAC
TC
C G
C TATG AG
T
T
T
C
G TC
AC A G
RFX GT
AA CTAG
A
C
GA
A
CT TG T
TTCAAAA
G C G A G A GC CT CT GTG
CT CTA
G
CCGCGCATGCGC
GT
Other
1 NRF T A G A CT A GT A CT A GT A GT
C G A
C CT A GT A CT A GT A
GGCCAATGAG
A CG
NFY CC
T
T
A
GT A GT A GT
C GT
C G A
C TC
T T
A
C
A
GACTACA TCCCAGAAGTGC
A AT TC C
GFY
A
CT GT
C GT A C
G A GT
C GT A
G
C
TGGAT
CA
G
C G A GT A GT A GTT
CT
GG
C
C
T
ATT
CA
C
G
A
0 Enrichment (%) Significance
Cask Madd Dock11 Dctn1
n.s.
Control siRNA 0 5 10 15 20 PDownloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Marcheva et al.
Alternative splicing of Cask and Madd directly more often in the clock mutants, encodes 58 amino acids
modulates β-cell insulin secretion upstream of its death domain, which inhibits caspase ac-
tivation (Fig. 4C; Kurada et al. 2009). To examine the im-
Given that Cask and Madd display among the highest de-
pact of the increased abundance of the short isoforms of
grees of exon inclusion level difference in Bmal1 −/− and
Cask and Madd transcripts in Bmal1 and Clock mutant
Clock −/− cells compared with WT (Fig. 2C,D), and that
β cells, we generated antisense oligonucleotides (ASOs)
THRAP3 is enriched at exons flanking the skipped exon
targeting the exons that were skipped more frequently
in Cask and Madd in Bmal1 −/− cells (Fig. 3B), we sought
in the mutants. ASOs are short single-stranded segments
to determine whether missplicing of Cask and Madd con-
of DNA that can affect expression of alternatively spliced
tributes to impaired β-cell function in Bmal1 −/− cells.
protein isoforms by initiating exon skipping or inclusion
Cask had an ∼36% reduction in inclusion of exon 11 in
by masking silencing sites during splicing. We treated
both Bmal1 −/− and Clock −/− cells (Fig. 4A), and Madd dis-
WT islets with control and targeting ASOs against Cask
played a 28% reduction in exon 26 in both Bmal1 −/− and
and Madd and observed ∼81% and ∼90% reduction in in-
Clock −/− cells (Fig. 4C). CASK was of particular interest as
clusion of Cask exon 11 and Madd exon 26 mRNA, re-
a member of the membrane-associated guanylate kinase
spectively, relative to a nontargeting control ASO (Fig.
(MAGUK) protein family that helps initiate assembly of
4B,D), while the ASOs did not alter expression of up-
the presynaptic secretory machinery. CASK is involved
stream exons 1 and 3 in Cask and Madd, respectively
in exocytosis of neurotransmitters in the brain (Atasoy
(Fig. 4B,D). Notably, islets treated with targeting ASOs
et al. 2007) and of insulin in the endocrine pancreas
against both Cask and Madd displayed significantly re-
(Suckow et al. 2008; Wang et al. 2015). Furthermore, Uni-
duced glucose-stimulated insulin secretion compared
prot and Interpro annotations show that exon 11 of Cask,
with control ASO-treated islets (by ∼40%) (Fig. 4B,D), sug-
which is skipped more frequently in the circadian mu-
gesting that clock-regulated splicing of these transcripts
tants, encodes a domain that facilitates interactions
supports β-cell function. Thus, these results demonstrate
with synaptic membrane proteins (Fig. 4A; Chetkovich
that mimicking exon skipping through the use of ASOs,
et al. 2002; Lee et al. 2002; Roh et al. 2002). Madd, a splice
which promotes expression of the short isoforms of these
variant of the insulinoma-glucagonoma clone 20 (IG20)
two genes whose splicing is controlled by the circadian
gene, contains a single nucleotide polymorphism associ-
clock, reduces glucose-stimulated insulin secretion, sug-
ated with human type 2 diabetes, and Madd knockout
gesting that missplicing of these mRNAs contributes to
mice display hyperglycemia and impaired insulin secre-
secretory failure in the setting of circadian disruption.
tion (Li et al. 2014). Exon 26 of Madd, which is skipped
A Altered Cask splicing in Bmal1-/- C Altered Madd splicing in Bmal1-/- Figure 4. Skipped exons within Cask and Madd
-/-
and Clock β-cells -/-
and Clock β-cells in circadian mutant β cells impair insulin secre-
Refseq transcripts exon 11 Refseq transcripts exon 26 tion. (A) Refseq transcripts depict the “long” and
Cask-long
Cask-short
Madd-long
Madd-short “short” (i.e., skipping exon 11) isoforms of Cask.
25 kb
G339 L L A A E R345
5 kb
G1287 R D K G S M W D Q L E D A A M E T F S L S1308
Exon 11 encodes amino acids 339–345, which
Protein domains Protein domains InterPro annotates as part of the C-terminal L27
domain. Sashimi plots of the differentially spliced
Cask gene in Bmal1 −/− and Clock −/− β-cell lines.
Kinase L27 L27 SHP PDZ Guanylate kinase uDENN cDENN dDENN Death
(1-250) (311-369) (370-426) (557-627) (458-539) (684-856) (13-267) (288-428) (430-564) (1366-1411)
Cask IncLevel: Madd IncLevel: (B) Expression of Cask skipped exon 11 or nontar-
30 65%
WT
60 71%
WT geted exon 1 in WT (n = 7) and Bmal1 −/− (n = 6) β-
0 Bmal1 -/- 0 Bmal1 -/- TC6 cells and following control (n = 5) and target-
Clock -/- Clock -/-
RPKM
RPKM
30 29% 60 43%
ing (n = 3–5) ASO treatment in WT islets assessed
0 0 via qPCR. Insulin secretion in the targeted islets,
30 28% 60 43%
Cask ASO-treated (n = 17 mice), compared with
0 0 control ASO-treated (n = 11 mice). (C) RefSeq tran-
Cask Madd
exon 11
ASO
exon 26
ASO scripts depict the “long” and “short” (i.e., skip-
13600456 chrX “-” strand 13586846 91155648 chr2 “-” strand 91154646 ping exon 26) isoforms of Madd. Exon 26
encodes amino acids 1287–1308, which InterPro
B Disrupted Cask splicing alters D Disrupted Madd splicing alters annotates as part of a disordered region 58 amino
insulin secretion in WT islets insulin secretion in WT islets acids upstream of its C-terminal death domain.
Exon 11 Exon 11 Exon 1 Exon 26 Exon 26 Exon 3 Sashimi plots of the differentially spliced Madd
1.5 1.5
n.s. * n.s.
gene in Bmal1 −/− and Clock −/− β-cell lines. (D) Ex-
Relative mRNA
Relative mRNA
*** *** **
expression
expression
1 1 pression of Madd skipped exon 26 or nontargeted
0.5 0.5
exon 3 in WT (n = 7) and Bmal1 −/− (n = 6) β-TC6
cells and following control (n = 5) and targeting (n
0 0 = 4) ASO treatment in WT islets assessed via
2 2
qPCR. Insulin secretion in the targeted islets,
Insulin release
Insulin release
WT WT
from control)
(fold change
from control)
(fold change
Bmal1-/- * Bmal1-/- * Madd ASO-treated (n = 15 mice) compared with
1 1
Control ASO Control ASO control ASO-treated (n = 9 mice).
Cask exon 11 ASO Madd exon 26 ASO
0 0
8 GENES & DEVELOPMENTDownloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press
Clock control of β-cell alternative splicing
Alterations in both transcription and alternative splicing data from islets isolated from high-fat diet (HFD)-fed com-
of Snap25a in circadian mutant and WT β cells pared with regular chow-fed mice (CON; >100 million
reads per sample; GSE92602) (Motterle et al. 2017). Princi-
Analysis of RNA-seq by rMATS of circadian mutant cells
ple component analysis (PCA) clustering revealed that AS
revealed that Snap25 was one of the few genes that was
events in islets from HFD-fed mice segregate from con-
both differentially expressed (74% and 54% reduction in
trol-fed mice (Supplemental Fig. S9A). Furthermore, we
mRNA in Bmal1 −/− and Clock −/− cells, respectively,
detected 259 differential AS slice events, including 170
compared with controls) and spliced (MXE: 15% and
SEs, 30 MXEs, 27 A5SS, and 32 A3SS, among 228 unique
25% reduced inclusion of alternative exon 5b versus 5a
genes (Δψ > 0.10, P < 0.05) (Supplemental Fig. S9B; Supple-
in Bmal1 −/− and Clock −/− cells) (Fig. 2A,C,D; Supplemen-
mental Table S5), while there were 187 genes differen-
tal Figs. S5D, S8A). Since SNAP25 is a key component of
tially expressed (log2 FC > 1, FDR < 0.05). Interestingly,
the core SNARE complex that mediates stimulus-depen-
similar to the clock mutant islets, there was little overlap
dent release of insulin from pancreatic β cells and overall
between differentially spliced and differentially expressed
expression levels were reduced in circadian mutant β
genes in the islets from HFD-fed mice (2.6%) (log2 FC > 1,
cells, we first tested whether overexpression of either of
FDR < 0.05). Of particular interest, the ortholog of the re-
the two major splice-variant isoforms of SNAP25 could re-
cently identified type II diabetes risk gene PAM (Thomsen
store secretion in clock mutant β cells. We generated sta-
et al. 2018) in humans was one of the most differentially
ble Bmal1 −/− and WT β-TC6 cell lines overexpressing full-
spliced transcripts in the islets from HFD-fed mice (Sup-
length human Snap25a or Snap25b cDNAs, which were
plemental Fig. S9B) despite similar total Pam expression
validated by qPCR and Western blot (Supplemental Fig.
in these data. This further provides evidence toward splic-
S8B). Surprisingly, overexpression of the Snap25a isoform,
ing as an inherent regulatory mechanism through which
which was more abundant relative to Snap25b in clock
β-cell function is mediated. To elucidate how these diet-
mutant cells, restored glucose-stimulated insulin secre-
induced changes in RNA processing intersect with circa-
tion in Bmal1 −/− cells to levels similar to the ones ob-
dian control, we next examined the overlap between AS
served in WT controls (Supplemental Fig. S8B).
events significantly altered in islets following HFD com-
However, overexpression of the Snap25b did not rescue
pared with those altered in the Bmal1 −/− cell lines (Δψ >
insulin secretion in the Bmal1 −/− cells despite high levels
0.10, P < 0.05). Surprisingly, we found a 15.8% overlap
of overexpression (Supplemental Fig. S8B). Overexpres-
(36 out of 228 AS HFD genes) between the two groups, in-
sion of either Snap25a or Snap25b above endogenous lev-
cluding in the TATA-binding protein (Tbp), the guanine
els in WT cells did not alter insulin secretion, indicating
nucleotide-binding protein (G protein; Gnas), and myo-
that reduced transcription of Snap25, as opposed to re-
sin5 (Myo5), and pathway analyses revealed that the
duced AS-mediated selection of the Snap25b isoform in
most enriched pathways at the intersection of HFD-in-
Bmal1 −/− β cells, contributes to impaired insulin secre-
duced and circadian-regulated splicing act to control
tion. To next test whether loss of either the Snap25a or
membrane protein trafficking and vesicle transport (Sup-
Snap25b isoforms per se was associated with altered
plemental Fig. S9C,D). Overall, the differential AS events
GSIS independently from differences in underlying tran-
were over-represented in gene ontology terms related to
scription, we used ASOs to block inclusion of alternative
synaptic vesicle transport and peptide secretion (Supple-
exon 5a or 5b and observed 90% and 83% reduction in
mental Fig. S5B).
exons 5a and 5b, respectively (Supplemental Fig. S8C). Im-
portantly, ASO treatment did not alter the total transcript
abundance of either isoform. We observed that loss of the
Snap25a transcript resulted in a 78% reduction in insulin Discussion
secretion in WT islets, while loss of the Snap25b isoform
Mounting evidence from genomic analyses has estab-
did not decrease the insulin response (Supplemental Fig.
lished extensive circadian control in the expression of tis-
S8C). Thus, our studies suggest that reduced overall
sue-specific gene networks central to metabolism (Koike
Snap25 expression levels, as opposed to reduced inclusion
et al. 2012; Remsberg et al. 2017; Yeung et al. 2018).
of alternative exon 5b, contribute to impaired insulin
Most studies have focused on using mRNA- and nascent
secretion in clock mutant β cells.
RNA sequencing in tandem with assays for measuring
TF binding to understand the role of the molecular clock
in initiating RNA synthesis (Fang et al. 2014; Zhang
Diet-induced obesity and circadian disruption impair
et al. 2014). More recent ribosome-associated RNA se-
shared exocytic pathways that are controlled by AS
quencing and proteomics have demonstrated that rhyth-
Having established that rhythmic AS impacts key regula- micity and phase in circadian transcription does not
tors of exocytosis in circadian mutant conditions, we necessarily reflect final protein expression, particularly
sought to determine whether impaired AS might repre- outside of the core circadian activator and repressor pro-
sent a more general defect associated with β-cell failure teins (Deery et al. 2009; Jang et al. 2015; Janich et al.
in the setting of type 2 diabetes. To determine whether 2015). Emergence of clock function has been implicated
impaired alternative pre-mRNA splicing might occur un- in β-cell maturation (Alvarez-Dominguez et al. 2020),
der conditions of β-cell disruption in type 2 diabetes and/ and circadian control of splicing events are necessary for
or obesity, we analyzed existing deep RNA sequencing the proper assembly of synaptic machinery in developing
GENES & DEVELOPMENT 9Downloaded from genesdev.cshlp.org on November 22, 2020 - Published by Cold Spring Harbor Laboratory Press Marcheva et al. neurons (Song et al. 2017; Wang et al. 2018). Despite this islets from hyperglycemic animals reveal over-representa- link between the circadian clock and neuronal cell-specif- tion of genes encoding vesicle trafficking and synaptic ic splicing, whether the circadian clock mediates AS proteins. Furthermore, rhythmic- and CLOCK/BMAL1- across the 24-h cycle in peripheral tissues remains incom- dependent transcriptomes revealed a repertoire of dynam- pletely understood. ic RBPs that mediate AS. There is evidence for RBPs Here we identified pre-mRNA alternative splicing as an regulating circadian splicing events (Benegiamo et al. output of the molecular clock that contributes to insulin 2018; Wang et al. 2018) and insulin secretion (Juan-Mateu secretion in pancreatic β cells. We also showed that dis- et al. 2017), but little is known about how circadian RBPs rupted AS of key exocytic genes following circadian dis- and downstream rhythmic RNA processing regulates pe- ruption contributes to impaired β-cell function. Our ripheral tissue function. Here we demonstrated that the analyses of RNA sequencing coverage of splice junctions circadian RBP THRAP3 regulates cassette exon skipping demonstrated extensive modulation of the circadian in β cells at transcripts controlling insulin secretion, in- transcriptome in islets at the level of mRNA isoform se- cluding Cask (a scaffold for calcium-calmodulin kinase) lection, and that genes encoding proteins mediating exo- and Madd (a RAB3 GTPase-activating protein). We show cytosis, synaptic transmission, and transcription are that proper Thrap3 expression is necessary for maintain- rhythmically alternatively spliced (Fig. 1; Supplemental ing exon skipping in Cask and Madd in Bmal1 −/− cells. Fig. S1). Global splicing patterns further indicated that a Surprisingly, simply mimicking the SE events found in majority of splicing events involve retention or exclusion Bmal1 −/− cells in Cask and Madd transcripts was suffi- of individual cassette exons (SE), and that these splicing cient to suppress glucose-stimulated insulin secretion in events display a bimodal distribution across the circadian WT mouse islets, suggesting that missplicing of mRNAs cycle, peaking at times corresponding roughly to the ze- such as Cask and Madd may contribute to secretory fail- nith and nadir of endogenous clock-driven insulin secre- ure with circadian disruption (Fig. 4). It is intriguing to tion (Fig. 1; Supplemental Fig. S1). Our observations speculate that reduced inclusion of the Cask exon 11 in suggest that the molecular clock within pancreatic β cells the circadian mutants leads to altered interactions with regulates insulin secretion by ensuring appropriate key synaptic membrane proteins such as Postsynaptic mRNA isoform selection at specific phases throughout density protein 95 (PSD-95), Neurexin-1, and Munc-18- the day/night cycle. interacting protein (MINT) (Chetkovich et al. 2002; Since the functional consequences of clock gene manip- Mukherjee et al. 2008), in turn impeding vesicle traffick- ulation to date have largely been performed in primary ing (Gillespie and Hodge 2013). Further analyses will be murine and human pancreatic islets containing mixed en- required to determine the impact of reduced inclusion of docrine cell types, our analyses of clonal Bmal1 and Clock these Cask and Madd exons on vesicle trafficking and cal- knockout β-cell lines enabled the interrogation of clock- cium signaling within circadian mutant β cells. These dependent transcriptional and AS events associated with findings reveal the importance of circadian control of AS impaired insulin secretion in pure populations of insu- of key exocytotic-controlling genes in the maintenance lin-secreting cells (Fig. 2; Supplemental Fig. S2). Genetic of β-cell function. depletion of Bmal1 and Clock resulted in phenotypic Snap25 was one of only four genes that were both dif- changes comparable with those observed in pancreas-spe- ferentially spliced and reduced in expression in both cific knockout mouse models, such as refractory secretory Clock −/− and Bmal1 −/− β cells. Snap25 encodes a target responses to both glucose and nonnutrient insulin secre- membrane SNARE protein critical for synaptic vesicle tagogues (Fig. 2; Supplemental Figs. S2–S4). Moreover, exocytosis and expresses either of the mutually exclusive electrophysiologic analysis of Bmal1 mutant β-cell lines exons 5a or 5b, which arose as a consequence of an ances- and mouse β cells revealed a conserved defect in depolari- tral duplication event (Bark 1993). While both SNAP25a zation-evoked exocytosis (Fig. 2B), localizing the secretory and SNAP25b are capable of assembling functional defect to the final stages of insulin granule fusion and pep- SNARE complexes in pancreatic β cells (Daraio et al. tide release. 2017), the relative roles of the Snap25a and Snap25b iso- The transcriptomes of Bmal1 −/− and Clock −/− β cells re- forms are not well understood. We observed reduced ex- vealed perturbations in mRNAs involved with peptide pression of both the Snap25a and Snap25b isoforms in secretion and second messenger signals that trigger insu- the circadian mutant β cells, with a greater reduction in lin secretion (Fig. 2A). We detected significant changes the Snap25b isoform relative to the Snap25a isoform (Sup- in the AS of ∼100 genes, including vesicle membrane plemental Fig. S8). Our findings that Snap25a overexpres- components, kinases, GTPases responsible for posttrans- sion enhanced insulin secretion in the circadian mutant β lational modifications to the secretory apparatus, and cells, and that blocking inclusion of the Snap25a but not plasma membrane proteins that are also important regula- Snap25b isoform using ASOs inhibited insulin secretion, tors of neuronal maturation and synaptic function (Fig. together suggests the β-cell clock modulates both splice 2D). A majority of these missplicing events occurred in isoform selection and transcription of rhythmic genes. mRNAs that were not differentially expressed in the Our studies verify the observation that Snap25a, which circadian mutants, highlighting the importance of clock is a less functional fetal isoform in neurons, is coexpressed control of isoform selection throughout the day. Our anal- with Snap25b in β cells and enhances insulin secretion yses of oscillating RNA transcripts in synchronized WT (Gonelle-Gispert et al. 1999; Daraio et al. 2017). Finally, islets, synchronized WT and clock deficient β cells, and observations that (1) AS of Snap25 and other neuronal 10 GENES & DEVELOPMENT
You can also read

















































