A new Norwegian Lemming subspecies from Novaya Zemlya, Arctic Russia
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ecologica Montenegrina 40: 93-117 (2021)
This journal is available online at: www.biotaxa.org/em
http://dx.doi.org/10.37828/em.2021.40.8
https://zoobank.org/urn:lsid:zoobank.org:pub:102DFA47-5952-437B-A41E-F1D2D82A1B80
A new Norwegian Lemming subspecies from Novaya Zemlya,
Arctic Russia
VITALY M. SPITSYN1,*, IVAN N. BOLOTOV1, ALEXANDER V. KONDAKOV1,
ANNA L. KLASS1, IVAN A. MIZIN2, ALENA A. TOMILOVA1, NATALIA A. ZUBRII1
& MIKHAIL Y. GOFAROV1
1
N. Laverov Federal Center for Integrated Arctic Research of the Ural Branch of the Russian Academy of Sciences,
Northern Dvina Emb. 23, 163000 Arkhangelsk, Russia
2
Russian Arctic National Park, Northern Dvina Emb. 36, 163051 Arkhangelsk, Russia
*Corresponding author: spitsyn.v.m.91993@yandex.ru
Received 25 January 2021 │ Accepted by V. Pešić: 16 March 2021 │ Published online 17 March 2021.
Abstract
Norwegian Lemming Lemmus lemmus is a remarkable population cycling species having a number of aposematic traits
in coloration and behavior. This species was thought to be the only mammal endemic to Fennoscandia. Here, we report
on the discovery of a distinct lineage of this species from Novaya Zemlya. This lineage is described here as the new
subspecies Lemmus lemmus chernovi ssp. nov. that morphologically differs from the nominate subspecies by having a
cryptic coloration. Our time-calibrated phylogeny revealed that this insular subspecies was isolated there since the
Eemian interglacial (mean age 93 Ka). Hence, bright coloration and antipredator behavior of the nominate subspecies
are novel aposematic traits that evolved since its isolation in a European refugium. This new discovery indicates that
Novaya Zemlya supported a cryptic polar refugium for cold-adapted terrestrial fauna during the Pleistocene. Finally,
our findings highlight that allopatric ranges of the true lemmings (Lemmus) reflect stepwise range expansions in cold
climatic episodes with subsequent isolation in refugia during interglacial events.
Key words: Arctic Ocean islands, cryptic polar refugium, Eemian interglacial, integrative taxonomy, Lemmus lemmus,
Lemmini, phylogeography, Pleistocene glaciations.
Introduction
Norwegian Lemming Lemmus lemmus (Linnaeus, 1758) is an iconic population cycling species, the density
of which explosively increases every 3-5 years, with subsequent mass movements mystifying people at least
since the Middle Ages (Kausrud et al. 2008; Lagerholm et al. 2017; Le Vaillant et al. 2018). The range of
Norwegian Lemming covers Norway, eastern Sweden, northern Finland, and the Kola Peninsula in Russia
(Abramson & Petrova 2018), and this species was thought to be one of a few animals endemic to
Fennoscandia (Lagerholm et al. 2014, 2017). Norwegian Lemming shares a remarkable suit of aposematic
Ecologica Montenegrina, 40, 2021, 93-117
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
traits that includes contrasting coloration with black, yellow and white colors, antipredator calls, and threat
postures (Andersson 2015).
It was thought that two lemming species occur on Novaya Zemlya: Siberian Lemming Lemmus
sibiricus (Kerr, 1792) and Arctic Collared Lemming Dicrostonyx torquatus (Pallas, 1778) (Wilson et al.
2017; Wilson & Reeder 2005). Conversely, we found that the Lemmus population from Novaya Zemlya is
phylogenetically distant from the Siberian Lemming, and is more closely related to the Norwegian Lemming.
This remarkable population is isolated on the Southern Island of Novaya Zemlya, a large mountainous land
with the maximum altitude of 1291 m. The Southern Island is separated from the Vaigach Island by the Kara
Strait, a 56 km wide marine channel. Palynological data suggests that the Southern Island was free of large
glacial shields during the entire Pleistocene epoch (Serebryanny & Malyasova 1998), and this hypothesis
was supported by a growing body of phylogeographic research (Potapov et al. 2018). The archipelago was a
closed nuclear test site for a half of century, and the current knowledge of its fauna is far from being
complete (Coulson et al. 2014).
This paper aims to (1) report on the first record of a Norwegian Lemming lineage outside
Fennoscandia; (2) describe this lineage as a new insular subspecies; and (3) discuss the origin, and
biogeographic patterns of the Norwegian Lemming subspecies in a broader taxonomic, phylogenetic and
evolutionary context.
Materials and methods
Data sampling
New samples of Lemmus specimens were obtained from the following areas: Southern Island, Novaya
Zemlya (n = 23; type series of L. lemmus chernovi ssp. nov.), Kola Peninsula (n = 7 [RMBH Lem035 –
Lem041]; L. lemmus lemmus), and Taymyr Peninsula (n = 1; L. sibiricus). The samples were collected using
series of mouse traps that were checked daily. The coordinates of localities are presented in Table 1. All
applicable international, national, and/or institutional guidelines for the care and use of animals were
followed. The sampling procedures were approved by the Ethics Committee on Animal Research of the
Institute of Ecological Problems of the North of Ural Branch of the Russian Academy of Sciences,
Arkhangelsk, Russia (order no. 001-05/15 of May 10, 2015).
Specimen preparation and morphometric study
The body length and weight of lemmings were measured using freshly dead specimens obtained from killing
mouse traps. The body length was measured using a tapeline (±1 mm). A tissue snip was taken from each
specimen and was preserved in 96% ethanol for subsequent molecular analyses. Skin samples were obtained
from 9 specimens of Lemmus lemmus chernovi ssp. nov., 7 specimens of L. l. lemmus, and 1 specimen of L.
sibiricus, while their heads and 14 whole specimens of L. lemmus chernovi ssp. nov. were fixed in 96%
ethanol. Heads were processed in 8% potassium hydroxide (KOH) for 2 hours and skulls were bleached in
3% hydrogen peroxide solution. Craniodental measurements were taken to the nearest 0.01 mm using a
digital caliper (Christoff et al. 2016).
Laboratory protocols and sequence alignment
This study includes new molecular data for 12 specimens of Lemmus species (Table 1). Total DNA was
extracted from ethanol-preserved soft tissue snips using the NucleoSpin® Tissue Kit (Macherey-Nagel
GmbH & Co. KG, Germany). For molecular analyses we obtained partial sequences of two mtDNA markers
as follows: the cytochrome c oxidase subunit I (COI) and the cytochrome b (cytb) gene fragments.
Additionally, we sequenced two nuclear markers as follows: the growth hormone receptor (GHR) and the
recombination activating protein 1 (RAG1) gene fragments. The PCR mix was as follows: total cell DNA
(200 ng), each primer (10 pmol), each dNTP (200 μmol), PCR buffer with 20 mmol MgCl 2 (2.5 μl), Taq
DNA polymerase (SibEnzyme Ltd., Russia) (0.8 units), and H2O (up to 25 μl of the total volume). Primer
sequences and thermocycling settings for PCR are shown in Table 2. Forward and reverse sequence reactions
were generated on purified PCR products using ABI PRISM® BigDye™ Terminator v. 3.1 reagents kit and
run on an ABI PRISM® 3730 DNA analyzer (Thermo Fisher Scientific Inc., Waltham, MA, USA). The
sequences were checked with BioEdit v. 7.2.5 (Hall 1999). Altogether 69 additional cytb sequences were
obtained from NCBI GenBank, and one sequence was digitized from a published paper (Lopatin et al. 2019)
94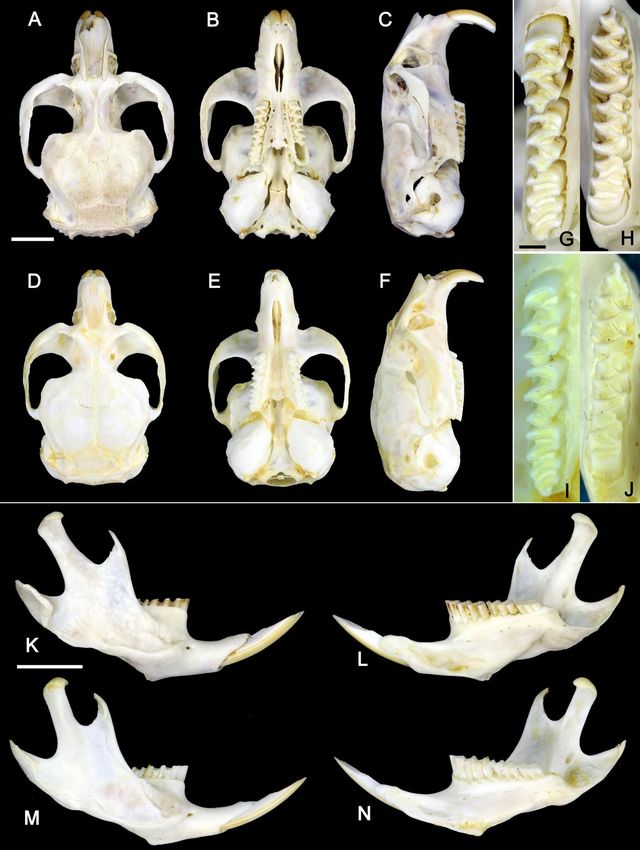
SPITSYN ET AL.
(Table 3). The alignment of the cytb, COI, GHR and RAG1 sequence data sets was performed directly using
the ClustalW algorithm of MEGA7 (Kumar et al. 2016).
Table 1. List of new molecular sequences of Lemmus taxa obtained in this study. Materials are deposited in the Russian
Museum of Biodiversity Hotspots (RMBH), N. Laverov Federal Center for Integrated Arctic Research of the Ural
Branch of the Russian Academy of Sciences (Arkhangelsk, Russia).
NCBI's GenBank acc. no. Specimen
Taxon Locality Latitude Longitude
Cytb COI GHR RAG1 Voucher
L. lemmus MK895056 N/A N/A N/A Lem005 Russia: Novaya 72.3822 52.7506
chernovi ssp. Zemlya, Malye
nov. Karmakuly
L. lemmus MK895057 MK895068 MK895075 MK895082 Lem008 Russia: Novaya 72.3933 52.7567
chernovi ssp. Zemlya, Malye
nov. Karmakuly
L. lemmus MK895058 MK895069 MK895076 MK895083 Lem020 Russia: Novaya 72.4058 52.8286
chernovi ssp. Zemlya, Malye
nov. Karmakuly
L. lemmus MK895059 MK895070 MK895077 MK895084 Lem029 Russia: Novaya 72.8500 52.6500
chernovi ssp. Zemlya,
nov. Bezymyannaya Bay
L. lemmus MK895061 MK895072 MK895079 MK895086 Lem035 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895062 MK895073 MK895080 MK895087 Lem036 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895063 N/A N/A N/A Lem037 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895064 MK895074 MK895081 MK895088 Lem038 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895065 N/A N/A N/A Lem039 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895066 N/A N/A N/A Lem040 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. lemmus MK895067 N/A N/A N/A Lem041 Russia: Kola 67.6633 33.7064
lemmus Peninsula, Khibiny
Mts.
L. sibiricus MK895060 MK895071 MK895078 MK895085 Lem034 Russia: Taymyr 69.4533 88.6697
Peninsula
Phylogenetic and phylogeographic analyses
For phylogenetic calculations, we used the cytb data set with 67 unique haplotype sequences of 834 bp long,
including four haplotypes of outgroup taxa, i.e. Dicrostonyx torquatus (Pallas, 1778), D. hudsonius (Pallas,
1778), Synaptomys cooperi (Baird, 1857), and Myopus schisticolor (Lilljeborg, 1844) (Tables 1 and 3). The
GTR+G+I evolutionary model was applied based on the corrected Akaike Information Criterion (AICc) of
MEGA7 (Kumar et al. 2016). The maximum likelihood phylogenetic analyses were computed with W-IQ-
TREE (Nguyen et al. 2015) through an online web server (http://iqtree.cibiv.univie.ac.at) (Trifinopoulos et
al. 2016). Node support values were calculated using an ultrafast bootstrapping algorithm implemented in
the web server (Hoang et al. 2017). Bayesian inference analyses were performed in MrBayes v. 3.2.6
(Ronquist et al. 2012) at the San Diego Supercomputer Center through the CIPRES Science Gateway (Miller
et al. 2010). Two runs, each with three heated (temperature = 0.1) and one cold Markov chain, were
conducted for 30 million generations. Trees were sampled every 1000th generation. After completion of the
MCMC analysis, the first 15% of trees were discarded as burn-in, and the majority rule consensus tree was
calculated from the remaining trees. Convergence of the MCMC chains was checked visually based on the
plotted posterior estimates using an MCMC trace analysis tool (Tracer v. 1.7) (Rambaut et al. 2018). For
Ecologica Montenegrina, 40, 2021, 93-117 95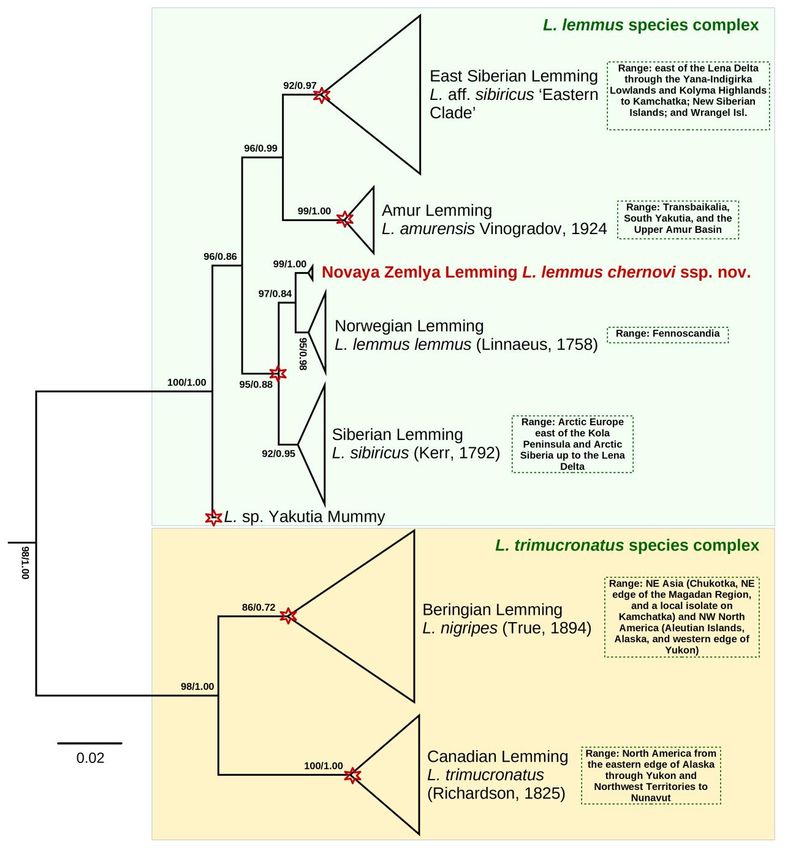
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
phylogeographic analyses, we used the cytb data set of Lemmus lemmus and L. sibiricus (length of sequences
= 792 bp). A median joining network approach was applied using Network v. 5.0.0.1 software with default
settings (Bandelt et al. 1999).
Table 2. Primer sequences for PCR amplification and sequencing.
Gene Primer’s name Thermocycling settings
Direction Sequence (5ʹ-3ʹ) References
fragment
cytb LemF Forward gmtcccthctdggmcthtgcc 4 min at 95°C, 28 cycles (50 This study
LemR Reverse gtwagbcctcgttgyttkgag seconds) at 94°C, 50 seconds at This study
60°C, 50 seconds at 72°C, and a
5 min final extension at 72°C
COI LoboF1 Forward kbtchacaaaycayaargayathgg 4 min at 95°C, 24 cycles (50 Lobo et al. (2013)
LoboR1 Reverse taaacytcwggrtgwccraaraayca seconds) at 94°C, 50 seconds at Lobo et al. (2013)
51°C, 50 seconds at 72°C, and a
5 min final extension at 72°C
GHR GHREXON10 Forward ggraarttrgaggaggtgaacacmatctt 4 min at 95°C, 30 cycles (50 Adkins et al. (2001)
GHREND Reverse ctactgcatgattttgttcagttggtctgtgctcac seconds) at 94°C, 50 seconds at Adkins et al. (2001)
67°C, 50 seconds at 72°C, and a
5 min final extension at 72°C
RAG1 S278F Forward gagcagtctccagtagttccaga 4 min at 95°C, 29 cycles (50 Schenk et al. (2013)
S281R Reverse ggtgcttacaactggtctcca seconds) at 94°C, 50 seconds at Justiniano et al.
58°C, 50 seconds at 72°C, and a (2015)
5 min final extension at 72°C
Divergence dating
Divergence times were estimated in BEAST v. 1.10.4 using a random local clock algorithm with the
Constant Coalescent model as the tree prior (Drummond & Rambaut 2007; Drummond et al. 2012). As an
input data, we used the restricted sequence data set with up to five cytb haplotypes per every species-level
taxon. We specified HKY+G+I substitution model to three partitions (3 codons of cytb) as suggested based
on the AICc of MEGA7 (Kumar et al. 2016). To dating the phylogeny, one published fossil calibration
(Abramson & Petrova 2018) was used with some modifications as follows: stem lineage of the Lemmus
clade; absolute age estimate 2.6 Ma; 95% soft upper bound 5.2 Ma (twice the age of the fossil); BEAST’s
prior settings: exponential distribution, mean (lambda) = 0.7, MRCA priors: Lemmus lemmus chernovi ssp.
nov. and L. nigripes (True, 1894). Additionally, one tip date was assigned as follows: Lemmus sp. ‘Yakutia
Mummy’; absolute age estimate 0.04 Ma (radiocarbon dating); Upper Pleistocene, Yana–Indigirka Lowland,
Yakutia (Lopatin et al. 2019). Three replicate searches were computed, each with 50 million generations.
The trees were sampled every 1,000th generation. Calculations were performed at the San Diego
Supercomputer Center through the CIPRES Science Gateway (Miller et al. 2010). The log files were
checked visually with Tracer v. 1.7 for an assessment of the convergence of the MCMC chains and the
effective sample size (ESS) of parameters (Rambaut et al. 2018). All ESS values were recorded as >2,000;
the posterior distributions were similar to the prior distributions. The resulting tree files from three
independent analyses were compiled with LogCombiner v. 1.10.4 using an additional resampling at every
5,000th generation (Drummond et al. 2012). The first 10% of trees were discarded as an appropriate burn-in.
The maximum clade credibility tree was obtained with TreeAnnotator v. 1.10.4 (Drummond et al. 2012).
Ancestral area and ancestral coloration reconstructions
The ancestral area patterns were estimated using two different approaches, i.e., Statistical Dispersal-
Vicariance Analysis (S-DIVA), Bayesian inference for discrete areas (BayArea) implemented in RASP v.
3.2 (Yu et al. 2015). For the ancestral area reconstruction, we used the set of 27,000 post-burn-in binary trees
that were combined from three runs of BEAST v. 1.10.4 (see above). As a condensed tree, we used the user-
specified consensus tree, which was calculated based on this set of trees with TreeAnnotator v. 1.10.4 (see
above). Out-group sequences were removed using the appropriate option of RASP v. 3.2 (Yu et al. 2015).
We coded four possible distribution areas of the Lemmus taxa as follows: (a) Europe, (b) North Asia (Siberia,
Russian Far East, New Siberian Archipelago, and Wrangel Island), (c) North America (with the Pribilof and
Aleutian Islands), and (d) Novaya Zemlya. The S-DIVA models were computed with the following priors:
max areas = 3; allow reconstruction with max value = 100; max reconstructions for final tree = 1000; and
96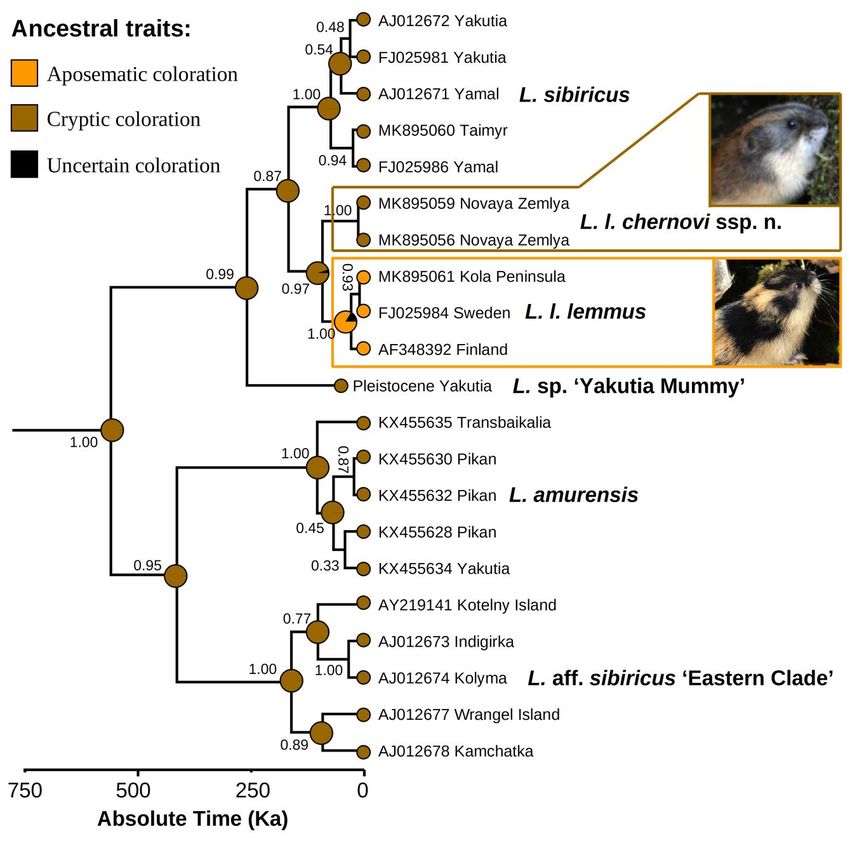
SPITSYN ET AL.
allowing extinctions. The BayArea analyses were run with default settings. In addition to the evaluations
obtained from each analysis separately, we used generalized results of the two modeling approaches, which
were combined using an algorithm implemented in RASP v. 3.2 (Yu et al. 2015).
Table 3. Additional cytb sequences of lemmings and related taxa used in this study.
Species Cytb GenBank acc. no. Locality References
Lemmus lemmus lemmus (Linnaeus, AF348392 Finland Fedorov & Stenseth
1758) (2001)
L. lemmus lemmus (Linnaeus, 1758) AF348391 Norway Fedorov & Stenseth
(2001)
L. lemmus lemmus (Linnaeus, 1758) FJ025984 Sweden: Västerbotten Abramson et al. (2008)
L. lemmus lemmus (Linnaeus, 1758) AY219145 Russia: Kola Peninsula Fedorov et al. (2003)
L. lemmus lemmus (Linnaeus, 1758) FJ025985 Russia: Kola Peninsula Abramson et al. (2008)
L. sibiricus (Kerr, 1792) AF348389 Russia: Yamal Peninsula Fedorov & Stenseth
(2001)
L. sibiricus (Kerr, 1792) AF348390 Russia: Yamal Peninsula Fedorov & Stenseth
(2001)
L. sibiricus (Kerr, 1792) AJ012672 Russia: Yakutia, Olenyok Gulf Fedorov et al. (1999)
L. sibiricus (Kerr, 1792) AY219140 Russia: Taymyr Peninsula Fedorov et al. (2003)
L. sibiricus (Kerr, 1792) [topotype AJ012671 Russia: Yamal Peninsula Fedorov et al. (1999)
of this taxon]
L. sibiricus (Kerr, 1792) AY219143 Russia: Taymyr Peninsula Fedorov et al. (2003)
L. sibiricus (Kerr, 1792) [topotype FJ025981 Russia: Yakutia, Lena River Delta Abramson et al. (2008)
of L. obensis bungei Vinogradov,
1926]
L. sibiricus (Kerr, 1792) FJ025980 Russia: Yakutia, Lena River Delta Abramson et al. (2008)
L. sibiricus (Kerr, 1792) FJ025986 Russia: Yamal Peninsula Abramson et al. (2008)
L. sp. “Yakutia Mummy” N/A Russia: Yakutia, right bank of the Tirekhtyakh River, Lopatin et al. (2019)
Indigirka River drainage [Pleistocene; calibrated 14C
age = 41305–41885 B.P.]
L. aff. sibiricus ‘Eastern Clade’ AJ012673 Russia: Yakutia, Indigirka River Delta Fedorov et al. (1999)
L. aff. sibiricus ‘Eastern Clade’ KY754011 Russia: Yakutia, near Kolyma River Delta Steppan & Schenk
(2017)
L. aff. sibiricus ‘Eastern Clade’ AJ012674 Russia: Yakutia, west of Kolyma River Delta Fedorov et al. (1999)
L. aff. sibiricus ‘Eastern Clade’ AY219144 Russia: Yakutia, near Kolyma River Fedorov et al. (2003)
L. aff. sibiricus ‘Eastern Clade’ AY219141 Russia: New Siberian Islands, Kotelny Island Fedorov et al. (2003)
L. aff. sibiricus ‘Eastern Clade’ AY219142 Russia: New Siberian Islands, Fadeyevsky Island Fedorov et al. (2003)
L. aff. sibiricus ‘Eastern Clade’ FJ025977 Russia: Kamchatka Peninsula, Uzon Caldera Abramson et al. (2008)
L. aff. sibiricus ‘Eastern Clade’ FJ025979 Russia: Kamchatka Peninsula, Uzon Caldera Abramson et al. (2008)
L. aff. sibiricus ‘Eastern Clade’ AJ012678 Russia: Kamchatka Peninsula, Uzon Caldera Fedorov et al. (1999)
L. aff. sibiricus ‘Eastern Clade’ KX455623 Russia: Magadan Region, Khinikanumsa River Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455624 Russia: Magadan Region, Khinikanumsa River Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455625 Russia: Magadan Region, Khinikanumsa River Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455626 Russia: Magadan Region, Yama River Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455627 Russia: Magadan Region, Yama River Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455621 Russia: Kamchatka Peninsula, Ust’-Kamchatsk Abramson & Petrova
(2018)
L. aff. sibiricus ‘Eastern Clade’ KX455622 Russia: Yakutia, Verhoyansky District, Nelgekhe Abramson & Petrova
[holotype of L. amurensis ognevi River (2018)
Vinogradov, 1933]
L. aff. sibiricus ‘Eastern Clade’ AJ012677 Russia: Wrangel Island Fedorov et al. (1999)
[topotype of L. portenkoi
Chernyavskii, 1967]
L. amurensis Vinogradov, 1924 KX455628 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
[holotype of this taxon] (2018)
L. amurensis Vinogradov, 1924 KX455629 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
(2018)
Ecologica Montenegrina, 40, 2021, 93-117 97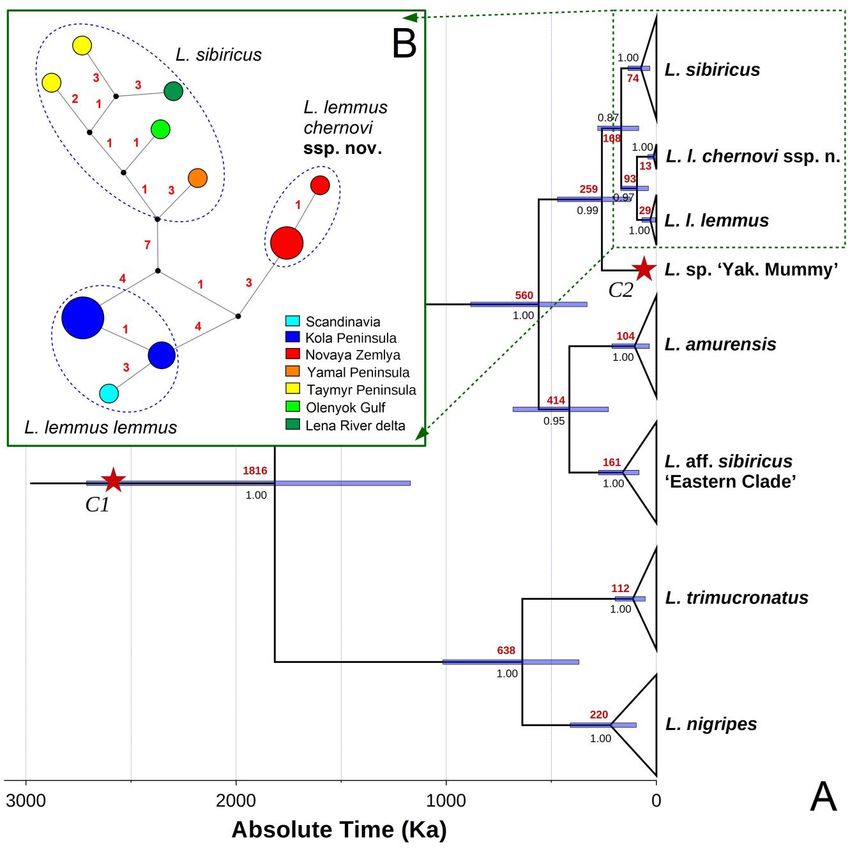
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
Species Cytb GenBank acc. no. Locality References
L. amurensis Vinogradov, 1924 KX455630 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455631 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455632 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455633 Russia: Amur Region, Zeysky District, Pikan Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455636 Russia: Yakutia, Neryungri District, Nagorny Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455634 Russia: Yakutia, Neryungri District, Chulman Abramson & Petrova
(2018)
L. amurensis Vinogradov, 1924 KX455635 Russia: Transbaikalia, Chita River upstream Abramson & Petrova
(2018)
L. nigripes (True, 1894) [topotype of AY219164 USA: Alaska, Pribilof Islands, Saint George Island Fedorov et al. (2003)
this taxon]
L. nigripes (True, 1894) [lectotype KX455619 Russia: Kamchatka Peninsula, south-western coast, Abramson & Petrova
of L. flavescens Vinogradov, 1926] probably Ust’-Bolsheretsk (2018)
L. nigripes (True, 1894) [topotype of KT877362 Russia: Gichiga, west coast of Okhotsk Sea, Abramson et al. (2018)
L. obensis chrysogaster Allen, 1903] Magadan Region
L. nigripes (True, 1894) KX455620 Russia: Kamchatka Peninsula, south-western coast, Abramson & Petrova
probably Ust’-Bolsheretsk (2018)
L. nigripes (True, 1894) KX455618 Russia, Chukotka Peninsula, Khatyrka River Abramson & Petrova
(2018)
L. nigripes (True, 1894) AJ012675 Russia, east of Kolyma River Delta, north-western Fedorov et al. (2003)
edge of the Chukotka Peninsula
L. nigripes (True, 1894) AY219158 Canada: Yukon Fedorov et al. (2003)
L. nigripes (True, 1894) AJ012676 USA: West Alaska Fedorov et al. (1999)
L. nigripes (True, 1894) AY219146 USA: Alaska, Seward Peninsula Fedorov et al. (2003)
L. nigripes (True, 1894) [topotype of AY219147 USA: Alaska, Point Barrow Fedorov et al. (2003)
L. alascensis Merriam, 1900]
L. nigripes (True, 1894) AY219148 USA: Alaska, Lake and Peninsula Fedorov et al. (2003)
L. nigripes (True, 1894) AY219149 USA: Alaska Fedorov et al. (2003)
L. nigripes (True, 1894) AY219152 USA: Alaska Fedorov et al. (2003)
L. nigripes (True, 1894) [topotype of AY219153 USA: Alaska, Goodnews Bay Fedorov et al. (2003)
L. minusculus Osgood, 1904]
L. nigripes (True, 1894) AY219154 USA: Alaska Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219162 Canada: Nunavut, Kugluktuk Fedorov et al. (2003)
[topotype of this taxon]
L. trimucronatus (Richardson, 1825) AY219161 Canada: Nunavut Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219155 Canada: Nunavut Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219159 Canada: Nunavut Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219163 Canada: Northwest Territories Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219156 Canada: Northwest Territories Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219157 Canada: Northwest Territories Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219167 USA: Alaska Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219150 USA: Alaska Fedorov et al. (2003)
L. trimucronatus (Richardson, 1825) AY219151 USA: Alaska Fedorov et al. (2003)
Dicrostonyx torquatus (Pallas, AJ131441 Russia: Yakutia, near Kolyma River Fedorov (1999)
1778)*
D. hudsonius (Pallas, 1778)* KY753975 Canada: Nunavut, Belcher Island Steppan & Schenk
(2017)
Synaptomys cooperi (Baird, 1857)* KY754162 USA: West Virginia, Allegheny Mountains Steppan & Schenk
(2017)
Myopus schisticolor (Lilljeborg, AF119263 N/A Conroy & Cook (1999)
1844)*
*These haplotypes were used as outgroup. N/A – not available.
To reconstruct ancestral coloration in Lemmus taxa, we applied Bayesian Binary MCMC (BBM)
analysis implemented in RASP v. 3.2 (Yu et al. 2015) using the same input data as for the ancestral area
reconstructions (see above). Two variants of coloration were coded as follows: (a) cryptic, and (b)
98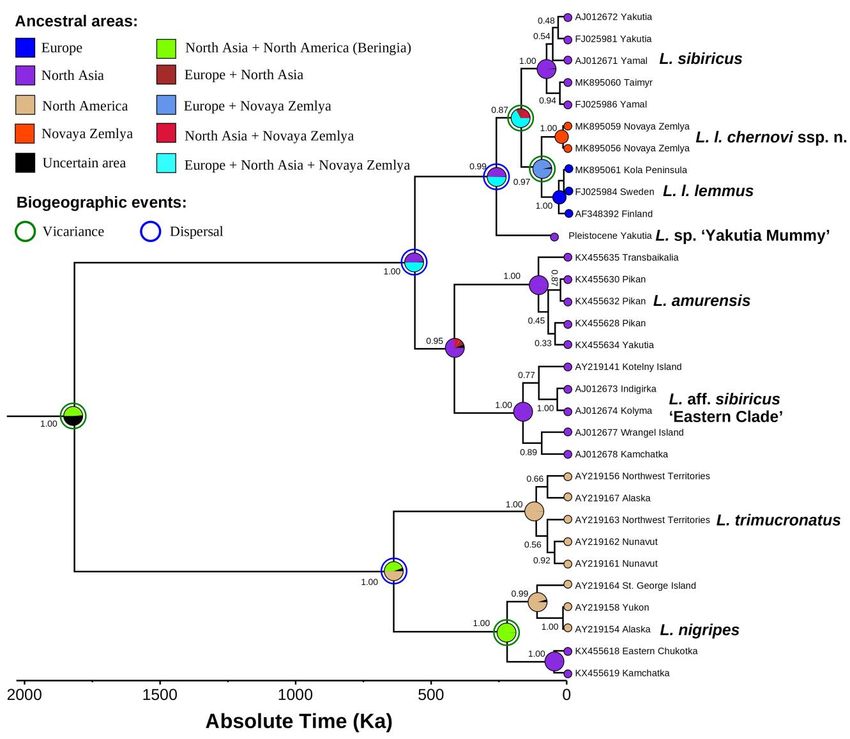
SPITSYN ET AL.
aposematic. The analyses were run under JC+G evolutionary model with the following parameters: 10
MCMC chains, 500,000 generations, sampling every 100th generation, and temperature = 0.1.
Species delimitation and molecular diagnostics
Molecular Operational Taxonomic Units (MOTUs) were delineated using two different approaches. First, we
applied the Bayesian Poisson Tree Process (bPTP) model thorough online bPTP server (http://species.h-
its.org/ptp) with 500,000 Markov Chain Monte Carlo (MCMC) generations and 15% burn-in (Zhang et al.
2013). Second, we computed the single-rate Poisson Tree Process (sPTP) model (p < 0.001) with online
mPTP server (http://mptp.h-its.org) (Kapli et al. 2017). As an input tree, we used the maximum likelihood
consensus phylogeny obtained from IQ-TREE analyses (see above). All outgroup taxa were removed from
the input tree using an appropriate option of the servers.
The molecular diagnosis of the new subspecies was designed using fixed nucleotide differences,
which were estimated for each gene separately (cytb, COI, GHR, and RAG1) using a Toggle conserved sites
tool of MEGA7 at 50 % level (Kumar et al. 2016). An alignment of haplotype sequences for the nominate
subspecies and Novaya Zemlya Lemming was performed using the ClustalW algorithm implemented in
MEGA7 (Kumar et al. 2016). Additionally, a mean p-distance to the nearest neighbor of each taxon under
discussion based on the cytb and COI haplotypes (if available) was calculated in MEGA7 (Kumar et al.
2016).
Data availability
The new sequences generated under this study are available from GenBank. Accession numbers for each
specimen are presented in Table 1. The voucher specimens are available in the RMBH, Russian Museum of
Biodiversity Hotspots, N. Laverov Federal Center for Integrated Arctic Research of the Ural Branch of the
Russian Academy of Sciences (Arkhangelsk, Russia).
Results
Phylogenetic position of Novaya Zemlya Lemming
Our molecular analyses of newly collected samples of the ‘Siberian’ lemming from Novaya Zemlya (Table
1, Fig. 1) indicate that it is a divergent phylogenetic lineage of the Norwegian Lemming (Figs 2-3), which
does not share aposematic traits and has cryptic gray coloration. The mean uncorrected p-distance between
the Fennoscandian and Novaya Zemlya lineages of Lemmus lemmus based on the cytb and COI gene
fragments is 1.2% and 1.0%, respectively (Table 4). Furthermore, the Novaya Zemlya Lemming shares two
diagnostic nucleotide substitutions in the GHR and RAG1 gene fragments compared with the nominate
subspecies (Table 5).
Table 4. Genetic divergences (mean uncorrected p-distances, %) between Lemmus taxa (L. lemmus and L. amurensis
species complexes) based on sequences of the mitochondrial cytb (below diagonal) and COI (above diagonal) genes.
Taxa LLC LLL LS LM* LSEC LA LT LN
L. lemmus chernovi ssp. nov. [LLC] 1.0 2.2 N/A N/A N/A N/A N/A
L. lemmus lemmus [LLL] 1.2 2.1 N/A N/A N/A N/A N/A
L. sibiricus [LS] 2.0 2.3 N/A N/A N/A N/A N/A
L. sp. “Yakutia Mummy” [LYM]* 1.7 2.0 2.2 N/A N/A N/A N/A
L. aff. sibiricus ‘Eastern Clade’ [LSEC] 4.5 4.9 4.5 2.8 N/A N/A N/A
L. amurensis [LA] 4.4 4.8 4.4 3.2 4.5 N/A N/A
L. trimucronatus [LT] 10.6 11.8 11.4 9.4 11.0 10.5 N/A
L. nigripes [LN] 8.7 9.8 10.1 7.8 8.9 8.4 7.2
*A frozen lemming mummy (14C age = 0.04 Ma) was collected from the Upper Pleistocene deposits in the Yana–
Indigirka Lowland, Yakutia, and its tissue snip was sequenced for a cytb gene fragment (Lopatin et al. 2019). N/A – not
available.
Ecologica Montenegrina, 40, 2021, 93-117 99A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
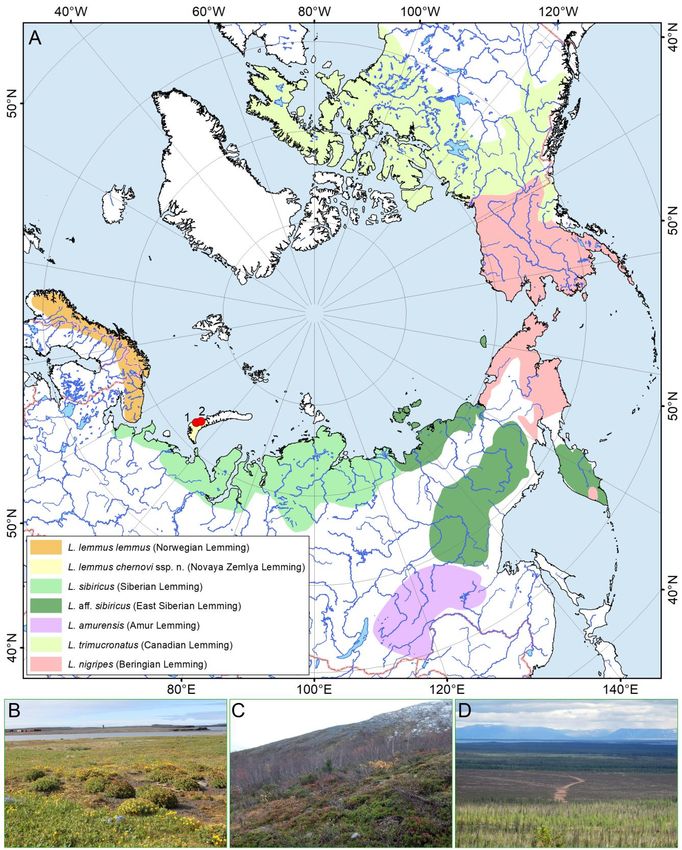
Figure 1. Ranges and habitats of the true lemmings (Lemmus). (A) Distribution map of Lemmus taxa. Ranges were
digitized based on published data and our records (Table 1). Red circles indicate sampling localities of Novaya Zemlya
Lemming (L. lemmus chernovi ssp. nov.): 1 – Malye Karmakuly Polar Station (type locality), 2 – Bezymyannaya Bay
(Table 1). The topographic base of the map was created with Natural Earth Free Vector and Raster Map Data
(www.naturalearthdata.com). The map was created using ESRI ArcGIS 10 software (www.esri.com/arcgis). (B, C, D)
Habitats of Lemmus taxa: (B) Novaya Zemlya Lemming (type locality, Novaya Zemlya), (C) Norwegian Lemming
(Kola Peninsula), and (D) Siberian Lemming (Taimyr Peninsula). (Photos: Vitaly M. Spitsyn [B, C] and Natalia A.
Zubrii [D])
100SPITSYN ET AL. Figure 2. Maximum likelihood (IQ-TREE) phylogeny of Lemmus taxa based on the cytb gene haplotypes. The Novaya Zemlya Lemming (Lemmus lemmus chernovi ssp. nov.) is colored red. The numbers near nodes are ultrafast bootstrap support values/Bayesian posterior probabilities. The red asterisks indicate the putative species-level clades supported by bPTP and sPTP species-delimitation models. The scale bar indicates the branch length. Outgroup taxa (Dicrostonyx torquatus, D. hudsonius, Synaptomys cooperi, and Myopus schisticolor) are not shown. Origin of Novaya Zemlya Lemming and its coloration patterns The fossil-calibrated phylogeny (Fig. 3A) revealed that the subspecies from Novaya Zemlya has been isolated from the Fennoscandian populations since the Eemian interglacial (mean age = 93 Ka, 95% HPD = 38–170 Ka). The generalized ancestral area reconstruction (S-DIVA + BayArea models) (Fig. 4, Table 6) suggested that the most recent common ancestor (MRCA) of the two subspecies of Lemmus lemmus has been widely distributed from Western Europe to Novaya Zemlya, most likely throughout the drying Arctic Shelf during a glacial event (probability = 96.6%). The Novaya Zemlya lineage was likely originated through a vicariance event associated with isolation of the insular population (probability = 95.9%). Ecologica Montenegrina, 40, 2021, 93-117 101
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
Figure 3. Phylogeny and phylogeography of Lemmus taxa. (A) Fossil-calibrated ultrametric chronogram of Lemmus
obtained with BEAST v. 1.10.4 for the cytb haplotype data set. Bars indicate 95% confidence intervals of the estimated
divergence times (Ka). Red and black numbers near nodes are mean ages (Ka) and BPP of BEAST, respectively. Red
stars indicate time calibrations: a stem lineage of the Lemmus clade with absolute age estimate 2,600 Ka and 95% soft
upper bound 5,200 Ka (C1); and Lemmus sp. ‘Yakutia Mummy’ with absolute age estimate 40 Ka (C2). Outgroup taxa
(Dicrostonyx torquatus, D. hudsonius, Synaptomys cooperi, and Myopus schisticolor) are not shown. (B) Median-
joining haplotype network of Novaya Zemlya, Norwegian, and Siberian lemmings based on the cytb sequences (N =
17). Circle size is proportional to the number of available sequences belonging to a certain haplotype (smallest circle =
one sequence). Small black dots indicate hypothetical ancestral haplotypes. Red numbers near branches indicate the
number of nucleotide substitutions between haplotypes.
Our ancestral trait modeling (Fig. 5) suggests that the MRCA of the two subspecies of Lemmus
lemmus has had a cryptic coloration (probability = 94.6%), and that bright coloration and specific
antipredator behavior of the Fennoscandian populations are novel aposematic traits that have been originated
since their isolation in a European refugium.
102SPITSYN ET AL.
Figure 4. Ancestral area reconstruction (combined scenario: S-DIVA + BayArea models) of Lemmus taxa based on our
fossil-calibrated BEAST v. 1.10.4 phylogeny. Black numbers near nodes are BEAST’s BPP.
Table 5. Polymorphism in the nuclear GHR and RAG1 gene sequences of Lemmus spp. Numbers in the top row refer to
nucleotide positions within a condensed sequence alignment, letters are FASTA codes for nucleotides but with
additional polymorphisms shown explicitly. Dots indicate a nucleotide matching the first sequence.
GHR RAG1
Taxon
268 377 439 719 812 860 294 332 399 436 440 546 599 707
L. lemmus lemmus A G C C C T G C G A C C C C
L. lemmus chernovi ssp. · · · · · C · · · · · · T ·
nov.
L. sibiricus · · C/T C/T C/T C · C/T · A/G C/T C/T T ·
L. aff. sibiricus ‘Eastern A/C A/G C/T · C/T · A · A · · · T T
Clade’
*GenBank data: MF074889 (GHR) and MF097872 (RAG1) [Russia: Yakutia, near Kolyma River Delta] (Steppan &
Schenk 2017).
Ecologica Montenegrina, 40, 2021, 93-117 103A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
Figure 5. Ancestral coloration reconstruction (Bayesian Binary MCMC analysis) of Lemmus taxa based on our fossil-
calibrated BEAST v. 1.10.4 phylogeny. Black numbers near nodes are BEAST’s BPP. The Lemmus trimucronatus + L.
nigripes clade (cryptic coloration) is omitted. (Photos: Vitaly M. Spitsyn).
Species delimitation and evolutionary biogeography of the true lemmings
Our Bayesian Poisson Tree Process (bPTP) and single-rate Poisson Tree Process (sPTP) species delimitation
modeling based on cytb haplotypes returned identical results with five Molecular Operational Taxonomic
Units (MOTUs) (Fig. 2; Appendix 1). Four of these MOTUs corresponded to the following species: L. aff.
sibiricus ‘Eastern Clade’, L. amurensis, L. nigripes, and L. trimucronatus (Richardson, 1825) (see Appendix
1 and discussion section for detail). Conversely, L. lemmus (with its subspecies) and L. sibiricus were treated
as a single MOTU.
Our fossil calibrated phylogeny (Fig. 3A) indicated that the genus Lemmus most likely originated in
the Early Pleistocene (mean age = 1.8 Ma; 95% HPD = 1.2-2.7 Ma) (Fig. 3A). The combined ancestral area
model failed to reconstruct the place of its origin with certainty (Fig. 4), while the S-DIVA reconstruction
suggested the Beringian origin of the genus (probability = 97.7%) (Table 6). The origin of the Lemmus
trimucronatus complex’s MRCA was placed in the Middle Pleistocene (mean age = 0.64 Ma; 95% HPD =
104SPITSYN ET AL.
0.37-1.02 Ma). It seems likely that this group has had the Nearctic origin with full support from the S-DIVA
reconstruction but a rather low probability based on the combined model (50.0%). Lemmus trimucronatus is
an entirely Nearctic species, while two isolates of L. nigripes on the both sides of the Bering Strait represent
fragments of its former continuous range throughout the Beringian Land Bridge (probability = 99.8%) in the
Middle Pleistocene (mean age = 0.22 Ma; 95% HPD = 0.10-0.41 Ma).
The Lemmus lemmus complex and a clade containing L. lemmus, L. sibiricus, and the extinct L. sp.
‘Yakutia Mummy’ have had a North Asian origin by the S-DIVA reconstruction (probabilities = 97.0-
97.7%), although the BayArea model failed to resolve ancestral areas for those groups with certainty,
suggesting the northern edge of Eurasia with the surrounding Arctic Shelf as a possible place (Fig. 4 and
Table 6). The L. amurensis + L. aff. sibiricus clade was originated in North Asia with a subsequent intra-area
radiation (probability = 73.4% by the combined scenario).
Table 6. The most probable ancestral areas of Lemmus clades inferred from two different statistical modeling
approaches and the combined reconstruction. High support values (probability ≥70 %) are highlighted in bold.
Type of Probability of ancestral areas (%)
Clades Ancestral areas* biogeographic Combined
S-DIVA BayArea
events* results
Lemmus N Asia & N America Vicariance 97.7 uncertain 48.9
[P = 12.4%] [all four areas:
100.0]
L. trimucronatus complex N America Dispersal 100.0 88.8 50.0
[P = 49.8%] [N Asia &
N. America]
L. nigripes North America – N Asia & N America Vicariance (Beringian 100.0 99.6 99.8
L. nigripes Asia Land Bridge)
[P = 95.0%]
L. lemmus complex Europe & Novaya Dispersal 97.7 98.0 49.5
Zemlya & N Asia [P = 18.4%] [N Asia]
L. amurensis + L. aff. N Asia Intra-area radiation 100.0 46.9 73.4
sibiricus ‘Eastern Clade’ [P = 72.7%]
L. lemmus + L. sibiricus + Europe & Novaya Dispersal 97.0 99.9 50.6
L. sp. ‘Yakutia Mummy’ Zemlya & N Asia [P = 33.8%] [N Asia]
L. lemmus + L. sibiricus Europe & Novaya Vicariance 33.7 100.0 66.8
Zemlya & N Asia [P = 63.7%]
L. lemmus lemmus – L. Europe & Novaya Vicariance 100.0 93.2 96.6
l. chernovi ssp. nov. Zemlya [P = 95.9%]
*Combined scenario.
Taxonomic description of Novaya Zemlya Lemming
Based on an integrative analysis of mitochondrial, nuclear (Figs 2-3, Tables 4-5) and morphological data
(Figs 6-7, Table 7), the Novaya Zemlya lineage of Lemmus lemmus is considered a new subspecies, which is
described here.
Lemmus lemmus chernovi Spitsyn, Bolotov & Kondakov ssp. nov.
Figs 6A, 7A-C, 7G-H, 7K-L, Table 7
Common name: Novaya Zemlya Lemming
Holotype: Male RMBH Lem005: type locality: Russia, Novaya Zemlya, Southern Island, near Malye
Karmakuly Polar Station, 72.3822°N, 52.7506°E, altitude 17 m, wet saxifrage-roseroot communities,
23.vii.2015, Spitsyn leg.
Paratypes: Russia, Novaya Zemlya, Southern Island: type locality, 27.vii–01.viii.2015, 11 specimens
(RMBH Lem001 – Lem004, Lem006, Lem007, Lem011, Lem019, Lem022, Lem023, Lem027), Spitsyn
leg.; near Malye Karmakuly Polar Station, 72.3575°N, 52.7799°E, altitude 27 m, dry lichen-moss rocky
tundra, 1.viii.2015, 1 specimen (RMBH Lem028), Spitsyn leg.; near Malye Karmakuly Polar Station,
72.3934°N, 52.7566°E, altitude 40 m, dry lichen-moss rocky tundra, 23.vii.2015, 2 specimens (RMBH
Lem008, Lem009), Spitsyn leg.; near Malye Karmakuly Polar Station, 72.4058°N, 52.8285°E, altitude 60 m,
dry tundra with Dryas octopetala and Astragalus alpinus, 27.vii.2015, 2 specimens (RMBH Lem020,
Ecologica Montenegrina, 40, 2021, 93-117 105A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
Lem021), Spitsyn leg.; near Malye Karmakuly Polar Station, 72.3797°N, 52.7525°E, altitude 20 m, wet
moss-sedge-willow tundra, 28–31.vii.2015, 4 specimens (RMBH Lem017, Lem018, Lem024, Lem025),
Spitsyn leg.; near Malye Karmakuly Polar Station, 72.3739°N, 52.7167°E, altitude 28 m, stinkweed
(Artemisia tilesii) assemblages, 5.viii.2015, 1 specimen (RMBH Lem031), Spitsyn leg.; Bezymyannaya Bay,
72.8°N, 52.5°E, 2017, 1 specimen (RMBH Lem029), local collector leg.
Molecular diagnosis: The novel subspecies differs from Lemmus lemmus lemmus by fixed nucleotide
substitutions in the mitochondrial cytb (12A, 36C, 189T, 441T, 573A, 726G, 765T) and COI (198C, 200A,
302T, 542A, 602C), and the nuclear GHR (860C) and RAG1 (599T) gene fragments (Tables 4-5).
Differential diagnosis: The Novaya Zemlya Lemming differs from the nominate subspecies by
having a cryptic coloration of dorsal surface that is dark gray with yellow and reddish hairs in adults (Fig.
6A) to reddish in juveniles, with an unclear longitudinal black stripe in anterior part from neck to nose (vs
bright yellow coloration with black anterior part of the trunk and large black spots on head). The new
subspecies is morphologically similar to Lemmus sibiricus but adult specimens from Novaya Zemlya could
be distinguished from it by dorsal coloration with yellow and reddish hairs (vs brown and reddish hairs).
Figure 6. Dorsal view of lemming specimens: (A) Novaya Zemlya Lemming (Lemmus lemmus chernovi ssp. nov.)
[holotype RMBH Lem005], and (B) Norwegian Lemming (L. lemmus lemmus) [RMBH Lem040, Kola Peninsula].
Scale bar = 10 mm. (Photos: Vitaly M. Spitsyn).
106SPITSYN ET AL. Figure 7. Comparison of crania, molar rows, and lower jaws of Novaya Zemlya (Lemmus lemmus chernovi ssp. nov.) [holotype RMBH Lem005] and Norwegian (L. l. lemmus) lemmings [specimen RMBH Lem040, Kola Peninsula]. (A, B, C) Crania of Novaya Zemlya Lemming: (A) dorsal view, (B) ventral view, and (C) lateral view. (D, E, F) Crania of Norwegian Lemming: (D) dorsal view, (E) ventral view, and (F) lateral view. Scale bar = 5 mm. (G, H) Molar rows of Novaya Zemlya Lemming: (G) maxillary, and (H) mandibular. (I, J) Molar rows of Norwegian Lemming: (I) maxillary, and (J) mandibular. Scale bar = 1 mm. (K, L) Lower jaw of Novaya Zemlya Lemming: (K) outer lateral view, and (L) inner lateral view. (M, N) Lower jaw of Norwegian Lemming: (M) outer lateral view, and (N) inner lateral view. Scale bar = 5 mm. (Photos: Vitaly M. Spitsyn). Ecologica Montenegrina, 40, 2021, 93-117 107
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
Description: Holotype male (Figs 6A, 7A-C, 7G-H, 7K-L, Table 7). Body weight 74 g; body length 130 mm,
tail 13 mm. Length of hind foot 15 mm. Length of upper molar row 8.5 mm, lower molar row 8.3 mm. Color
of dorsal surface grayish yellow, with an unclear longitudinal black stripe in anterior part from neck to nose.
Cheeks dark gray, nose brownish gray. Ventral surface light-colored, creamy-yellow. Tail creamy. Vibrissae
up to 33 mm long, black with yellow apex. Rostrum shorter than 2/3 greatest skull length; nasals do not
proceed beyond the frontal edge of the incisors, rear edge rounded; gnathic process not pronounced;
premaxilla rounded. Lacrimals with small ledge on the back side. Zygomatic arches parallel, jugal bone
broad. Zygomatic plate broad, with slightly concave frontal margin. Zygomatic notch shallow. Nasolacrimal
foramen with irregular shape, broad. Interorbital region narrow. Braincase region broad, almost square, with
concavities behind the zygomatic arches. Lambdoidal ridge well developed. Postglenoid foramen elongated.
Diastema long, concave; incisive foramina long and narrow. Mesopterygoid fossa broad, biconcave;
parapterygoid fossa large; auditory bulla large, drop-shaped. Lower jaw large, angular process, condylar
process и coronoid process large, well developed. Maxillary incisors yellowish-orange. Morphological
variability. Mean body length of the type specimens (±s.e.m.) 110.9±19.6 mm (min-max = 85-160 mm; n =
21). Color of dorsal surface grayish yellow in adults to reddish in juveniles, with an unclear longitudinal
black stripe (Fig. 6A).
Table 7. Body and cranial measurements (mm) of Novaya Zemlya and Norwegian lemmings.
Lemmus lemmus chernovi ssp. nov. (Novaya Zemlya, type series, Lemmus lemmus lemmus (Kola Peninsula, adults,
adults, N = 4) N = 3)
Parameters
Holotype Paratype Paratype Paratype Mean ± Mean ±
Lem036 Lem035 Lem038
Lem005 Lem028 Lem019 Lem029 s.e.m. s.e.m.
Body length 130 125 145 125 131±5 100 110 110 107±3
Tail length 13 12 15 13 13.3±0.6 11 12 12 11.7±0.3
Hind foot length 15 14 16 15 15.0±0.4 14 16 15 15.0±0.6
Greatest skull length 31.4 31.7 32.7 30.4 31.6±0.5 25.4 27.2 27.0 26.5±0.6
Interorbital breadth 4.1 4.0 N/A 4.2 4.1±0.1 4.0 4.4 4.0 4.1±0.1
Zygomatic breadth 21.8 21.4 23.4 21.2 22.0±0.5 18.4 19.4 18.4 18.7±0.3
Length of rostrum 8.7 8.7 9.3 8.3 8.8±0.2 N/A 7.3 6.9 7.1±0.2
Breadth of rostrum 6.5 6.5 6.9 6.4 6.6±0.1 5.5 5.5 5.6 5.5±0.0
Breadth of braincase 16.0 14.9 16.6 15.0 15.6±0.4 14.1 14.1 14.1 14.1±0.0
Height of braincase 10.5 10.1 11.5 10.2 10.6±0.3 9.5 9.5 9.1 9.4±0.1
Length of diastema 11.1 11.1 11.2 10.8 11.1±0.1 9.4 9.4 9.3 9.4±0.0
Length of incisive 6.2 6.6 6.6 6.2 6.4±0.1 5.2 5.9 6.4 5.8±0.3
foramina
Breadth of incisive 1.3 1.3 1.5 1.4 1.4±0.0 1.0 1.1 1.2 1.1±0.1
foramina
Palatal length 7.8 7.5 8.0 7.8 7.8±0.1 7.8 7.6 7.8 7.7±0.1
Postpalatal length 9.1 9.0 9.7 8.8 9.2±0.2 8.3 9.0 8.4 8.6±0.2
Breadth of 3.9 4.4 4.6 4.3 4.3±0.1 4.2 4.2 4.1 4.2±0.0
mesopterygoid fossa
Length of bulla 9.9 9.1 9.9 8.2 9.3±0.4 7.9 7.9 7.9 7.9±0.0
Crown length of 8.5 8.6 8.9 8.2 8.6±0.1 7.2 7.2 7.2 7.2±0.0
maxillary molar row
Crown length of 8.3 8.1 8.5 7.9 8.2±0.1 7.0 7.0 7.1 7.0±0.0
mandibular row
Etymology: This subspecies is named in memory of the late Academician of the Russian Academy
of Sciences Yury I. Chernov (1934-2012), a prominent Russian zoologist, biogeographer, and Arctic
explorer.
Distribution and habitat: This subspecies is endemic to the Southern Island of Novaya Zemlya, while
its reliable records from the Northern Island of this archipelago are unknown (Fig. 1A). It inhabits a broad
range of meadow-like (Fig. 1B) and tundra habitats.
Comments: The species delimitation modeling (Fig. 2) suggests that Lemmus lemmus and L.
sibiricus may belong to one polymorphic species, although molecular distances between these taxa
correspond to an interspecific difference (Tables 4-5) supporting the modern concept of two young allopatric
species (Lagerholm et al. 2014, 2017; Abramson & Petrova 2018; Wilson & Reeder 2005; Wilson et al.
2017; Abramson et al. 2018).
108SPITSYN ET AL. Discussion Discovery of a new Lemmus lemmus subspecies on Novaya Zemlya Here, we report on the unexpected discovery of a Norwegian Lemming lineage from Novaya Zemlya. This lineage shares a shallow genetic divergence from the nominate subspecies of Lemmus lemmus from Fennoscandia (1.2% and 1.0% by the cytb and COI gene fragments, respectively). Our fossil-calibrated phylogeny indicates that the Novaya Zemlya lineage was separated from Fennoscandian populations since the Eemian interglacial (mean age = 93 Ka). These novel results agree with the paleogenetic and DNA-based data that the Norwegian Lemming originated from a small population surviving the Last Glacial Maximum in an ice-free Scandinavian refugium (Fedorov & Stenseth 2001; Lagerholm et al. 2014) but we present the first evidence that other isolate of this species evolved simultaneously and independently on Novaya Zemlya. Hence, the MRCA of the Lemmus lemmus clade was widely distributed throughout the drying Arctic Shelf from Western Europe to Novaya Zemlya before the Eemian interglacial, as suggested based on our generalized ancestral area reconstruction (S-DIVA + BayArea models; probability = 96.6%). This broad range could have been originated during a cold event corresponding to the marine isotope stage (MIS) 6 [the penultimate glaciation], when lemmings abundantly occurred in Western (Lemmus lemmus) and Eastern (L. lemmus and L. sibiricus) Europe (Markova & Puzachenko 2017). The rising of the sea level during the subsequent Eemian interglacial (MIS 5) (Markova & Puzachenko 2017) was the most likely driver of isolation of Novaya Zemlya from the mainland followed by a vicariance event in the MRCA of L. lemmus. Origin of aposematic traits in Norwegian Lemming Our ancestral trait modeling results reveal that aposematic coloration and behavior of the Norwegian Lemming are secondary adaptive traits, which were lacking in the MRCA of the Fennoscandian and Novaya Zemlya lineages. It seems likely that these traits rapidly evolved in the nominate subspecies of Lemmus lemmus in the Late Pleistocene (mean age = 93 Ka, 95% HPD = 38–170 Ka). A well-preserved frozen lemming mummy (Lemmus sp. ‘Yakutia Mummy’) was discovered recently from the Upper Pleistocene deposits in the Yana–Indigirka Lowland, Yakutia (Lopatin et al. 2019). This ancient lemming (calibrated 14C age = 40 Ka) represents a phylogenetic lineage sister to the L. lemmus + L. sibiricus clade (Fig. 3A and Table 4), and it shares gray coloration with yellowish hairs (Lopatin et al. 2019). Externally, it is rather similar to cryptically colored lemmings, e.g. Lemmus lemmus chernovi ssp. nov., L. sibiricus, and L. aff. sibiricus ‘Eastern Clade’. These findings indicate that our reconstruction is correct and that cryptic coloration should be considered an ancestral trait for the entire Lemmus clade. Cryptic polar refugium on Novaya Zemlya Our discovery of an endemic insular subspecies of Lemmus lemmus aligns with a small body of earlier research indicating that Novaya Zemlya supported a cryptic glacial refugium for terrestrial fauna and flora during the Pleistocene epoch (Serebryanny & Malyasova 1998; Potapov et al. 2018). Dicrostonyx torquatus ungulatus Baer, 1841 was described as a subspecies endemic to Novaya Zemlya (Baranova & Gromov 2003), and needs future DNA-based studies as a possible additional example of a distant insular lineage in small rodents. The bumble bee species Bombus glacialis Friese, 1902 is known to occur on Novaya Zemlya, and, probably, on Wrangel Island (Potapov et al. 2018). This species did not cross the Kara Strait, a 56 km wide marine barrier, separating Novaya Zemlya from the Vaigach Island (Potapov et al. 2019). The Arctic charr Salvelinus alpinus (Linnaeus, 1758) populations in land-locked lakes on Novaya Zemlya share unique mtDNA haplotypes that may reveal their long-term (pre-glacial) existence there (Makhrov et al. 2019). Palynological data indicate the presence of large ice-free refugia with species-rich plant assemblages on Novaya Zemlya during the entire Pleistocene (Serebryanny & Malyasova 1998). Conversely, several mammal species can migrate to Novaya Zemlya from the mainland during the winter months, as it was shown for the reindeer Rangifer tarandus (Linnaeus, 1758) (Kvie et al. 2016), and the mountain hare Lepus timidus (Linnaeus, 1758) (Spitsyn & Bolotov 2020). It is clear that molecular genetic studies on animal groups with lower dispersal abilities such as sawflies (Hymenoptera: Symphyta) (Taegeret et al. 2006) and tadpole shrimps (Crustacea: Notostraca) could contribute to future understanding of the role of Novaya Zemlya as a cryptic polar refugium. Preliminary hypothesis on evolutionary biogeography of the true lemmings Based on our time-calibrated phylogeny, ancestral area reconstruction, and available published data, the Ecologica Montenegrina, 40, 2021, 93-117 109
A NEW NORWEGIAN LEMMING SUBSPECIES FROM NOVAYA ZEMLYA
following evolutionary scenario for Lemmus taxa can be proposed. The basal split of the genus Lemmus into
the two species complexes indicates that the MRCA of this group has had a trans-Beringian range.
Therefore, it could have been originated in Beringia approximately 2,000 Ka ago or so. This date aligns well
with an earlier fossil-calibrated phylogeny of the true lemmings and available fossil records (Abramson &
Petrova 2018). The Eurasian group (Lemmus lemmus species complex) dispersed throughout the Far East and
Siberia to the west approximately 560 Ka ago, during a short and mild glacial epoch (MIS 14) (Hao et al.
2015). Later, this continuous range was subdivided into several fragments in East Siberian mountains
(approximately 400 Ka ago, L. amurensis and L. aff. sibiricus ‘Eastern Clade’ in Transbaikalia and Kolyma
Highlands, respectively), Yana-Indigirka Lowland (approximately 260 Ka ago, L. sp. ‘Yakutia Mummy’),
and an area west of the Lena River basin (the stem lineage of L. sibiricus and L. lemmus). The first split
corresponds to a global interglacial event during MIS 11 stage (Melles et al. 2012) while the second event
can be linked to a warm climatic episode at MIS 7 stage (Berger et al. 2016). After that, lemmings colonized
Western Siberia, the Urals, Novaya Zemlya, and Europe (approximately 170 Ka ago, the MRCA of Lemmus
sibiricus and L. lemmus). As it was noted above, this massive westward dispersal event coincides with the
penultimate glacial maximum (MIS 6) 190-130 Ka ago, and is traced well in paleontological records
(Markova & Puzachenko 2017).
It is clear that the Beringia could have played a critical role in the evolutionary history of the true
lemmings, as the basal split in the genus and at least one further divergence event (i.e. the intraspecific split
in Lemmus nigripes) were likely associated with the opening and closure of this land bridge. Moreover, the
emerged Bering Land Bridge and surrounding shelf areas most likely served as a corridor for migration of L.
nigripes individuals with Nearctic haplotypes to the Chukotka Peninsula and of L. aff. sibiricus ‘Eastern
Clade’ from the Kolyma Highland to the New Siberian Archipelago and Wrangel Island during the Last
Glacial Maximum (approximately 18-25 Ka ago) (Elias & Brigham-Grette 2013).
Taxonomy and species richness of the true lemmings (Lemmus)
The number of valid taxa in this genus is a matter of long-term debates (Pokrovski et al. 1984; Abramson et
al. 2008; Abramson & Petrova 2018; Abramson et al. 2018; Lissovsky et al. 2019). Recently, a group of
researchers lumped all these taxa except for one (Lemmus ‘trimucronatus’) into a single species, L. lemmus,
which is thought to be widespread throughout Northern Eurasia from the Atlantic Coast to the Chukotka and
Kamchatka peninsulas and Wrangel Island (Lissovsky et al. 2019). Similar concepts with one or two
polymorphic and widespread Lemmus species were proposed repeatedly by earlier scholars (see review
Abramson et al. 2008). However, this ‘lumping’ approach does not consider multiple morphological,
genetic, and biogeographic evidences that strongly indicate the distinctiveness of these taxa and their origin
in separate Pleistocene refugia (Fedorov et al. 1999, 2003; Fedorov 1999; Fedorov & Stenseth 2001; Wilson
& Reeder 2005; Abramson et al. 2008, 2018; Lagerholm et al. 2014; Wilson et al. 2017; Abramson &
Petrova 2018). It was shown that L. lemmus, L. sibiricus, and L. amurensis can hybridize with each other and
that these three taxa shared a strong reproductive isolation from the Beringian species Lemmus nigripes
(True, 1894) (Pokrovski et al. 1984). Several recent studies provided DNA sequences of topotypes for most
nominal taxa described from Northern Eurasia and North America that can serve as the robust molecular
basis for an integrative taxonomic revision (Fedorov et al. 2003; Abramson & Petrova 2018; Abramson et al.
2018). Based on these earlier works (Fedorov et al. 2003; Abramson et al. 2008, 2018; Abramson & Petrova
2018) and our own data, we separate six prospective species of the true lemmings, which are characterized
here (see Appendix 1). Our view on the genus should be considered as a preliminary taxonomic hypothesis
that needs future research efforts using nuclear DNA gene sequences and genomic datasets.
Acknowledgements
The collection of lemmings from Novaya Zemlya in 2015 was performed during the ‘Floating University’
Scientific Expedition of the Northern Arctic Federal University. Russian Foundation for Basic Research
supported I.N.B. and V.M.S. under grant No. 19-34-90012, and A.V.K. under grant No. 18-44-292001. The
Ministry of Science and Higher Education of the Russian Federation supported this work as follows: project
АААА-А17-117033010132-2 to V.M.S. and N.A.Z., and project АААА-А18-118012390161-9 to M.Y.G.
and A.A.T. The Russian Science Fundation supported A.V.K. and I.N.B. under project No. 19-04-00066. We
are grateful to the reviewers who helped us to improve earlier versions of this paper.
110SPITSYN ET AL.
References
Abramson, N., Dokuchaev, N. & Petrova, T. (2018) Long-standing taxonomic and nomenclature issue of
Lemmus obensis chrysogaster Allen, 1903 (Rodentia, Cricetidae) resolved. Mammalia, 82, 167–172.
https://doi.org/10.1515/mammalia-2016-0081
Abramson, N. I., Kostygov, A. Y. & Rodchenkova, E. N. (2008) The taxonomy and phylogeography of
Palaearctic true lemmings (Lemmus, Cricetidae, Rodentia): New insights from cyt b data. Russian
Journal of Theriology, 7(1), 17–23. https://doi.org/10.15298/rusjtheriol.7.1.03
Abramson, N. I. & Petrova, T. V. (2018) Genetic analysis of type material of the Amur lemming resolves
nomenclature issues and creates challenges for the taxonomy of true lemmings (Lemmus, Rodentia:
Cricetidae) in the eastern Palearctic. Zoological Journal of the Linnean Society, 182(2), 465–477.
https://doi.org/10.1093/zoolinnean/zlx044
Adkins, R. M., Gelke, E. L., Rowe, D. & Honeycutt, R. L. (2001) Molecular phylogeny and divergence time
estimates for major rodent groups: evidence from multiple genes. Molecular Biology and Evolution,
18, 777–791. https://doi.org/10.1093/oxfordjournals.molbev.a003860
Allen, G. M. (1914) Mammals. Notes on the birds and mammals of the Arctic coast of east Siberia.
Proceedings of the New England Zoölogical Club, 5, 49–66.
Andersson, M. (2015) Aposematism and crypsis in a rodent: antipredator defence of the Norwegian
lemming. Behavioral Ecology and Sociobiology, 69, 571–581. https://doi.org/10.1007/s00265-014-
1868-7
Bandelt, H. J., Forster, P. & Röhl, A. (1999) Median-joining networks for inferring intraspecific phylogenies.
Molecular Biology and Evolution, 16(1), 37–48.
https://doi.org/10.1093/oxfordjournals.molbev.a026036
Baranova, G. I. & Gromov, I. M. (2003) Rodentia. Catalogue of Type Specimens in the Collections of the
Zoological Institute, 4, 1–100.
Berger, A., Crucifix, M., Hodell, D. A., Mangili, C., McManus, J. F., Otto Bliesner, B., Pol, K., Raynaud, D.,
Skinner, L. C., Tzedakis, P. C., Wolff, E. W., Yin, Q. Z., Abe-Ouchi, A., Barbante, C., Brovkin, V.,
Cacho, I., Capron, E., Ferretti, P., Ganopolski, A., Grimalt, J. O., Hönisch, B., Kawamura, K.,
Landais, A., Margari, V., Martrat, B., Masson-Delmotte, V., Mokeddem, Z., Parrenin, F.,
Prokopenko, A. A., Rashid, H., Schulz, M. & Vazquez Riveiros, N. (2016) Interglacials of the last
800,000 years. Reviews of Geophysics, 54, 162–219. https://doi.org/10.1002/2015RG000482
Christoff, A. U., Vieira, E. M., Oliveira, L. R., Gonçalves, J. W., Valiati, V. H. & Tomasi, P. S. (2016) A
new species of Juliomys (Rodentia, Cricetidae, Sigmodontinae) from the Atlantic Forest of Southern
Brazil. Journal of Mammalogy, 97, 1469–1482. https://doi.org/10.1093/jmammal/gyw082
Conroy, C. J. & Cook, J. A. (1999) MtDNA evidence for repeated pulses of speciation within arvicoline and
murid rodents. Journal of Mammalian Evolution, 6, 221–245.
https://doi.org/10.1023/A:1020561623890
Coulson, S. J., Convey, P., Aakra, K., Aarvik, L., Ávila-Jiménez, M. L., Babenko, A., Biersma, E. M.,
Boström, S., Brittain, J. E., Carlsson, A. M., Christoffersen, K., De Smet, W. H., Ekrem, T.,
Fjellberg, A., Füreder, L., Gustafsson, D., Gwiazdowicz, D. J., Hansen, L. O., Holmstrup, M., Hullé,
M., Kaczmarek, Ł., Kolicka, M., Kuklin, V., Lakka, H.-K., Lebedeva, N., Makarova, O., Maraldo,
K., Melekhina, E., Ødegaard, F., Pilskog, H. E., Simon, J. C., Sohlenius, B., Solhøy, T., Søli, G.,
Stur, E., Tanasevitch, A., Taskaeva, A., Velle, G., Zawierucha, K. & Zmudczyńska-Skarbek, K.
(2014) The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents
Sea; Svalbard, Franz Josef Land and Novaya Zemlya. Soil Biology and Biochemistry, 68, 440–470.
https://doi.org/10.1016/j.soilbio.2013.10.006
Desmarest, A. G. (1822) Mammalogie ou description des espèces des Mammifères. Seconde partie,
contenant les ordres des rongeurs, des edentes, des pachydermes, des ruminans et des cretaces.
Chez Mme veuve Agasse, Imprimeur-Libraire, Paris, 556 pp.
Drummond, A. J. & Rambaut, A. (2007) BEAST: Bayesian evolutionary analysis by sampling trees. BMC
Evolutionary Biology, 7(214). https://doi.org/10.1186/1471-2148-7-214
Drummond, A. J., Suchard, M. A., Xie, D. & Rambaut, A. (2012) Bayesian phylogenetics with BEAUti and
the BEAST 1.7. Molecular Biology and Evolution, 29, 1969–1973.
https://doi.org/10.1093/molbev/mss075
Elias, S. A. & Brigham-Grette, J. (2013) Late Pleistocene Glacial Events in Beringia. In: Elias, S. A. &
Ecologica Montenegrina, 40, 2021, 93-117 111You can also read

















































