A central serotonin regulating gene polymorphism (TPH2) determines vulnerability to acute tryptophan depletion-induced anxiety and ventromedial ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
A central serotonin regulating gene polymorphism (TPH2)
determines vulnerability to acute tryptophan depletion-induced anxiety
and ventromedial prefrontal threat reactivity
Congcong Liu1,2,3, Keshuang Li1,3,4, Meina Fu3, YingYing Zhang3, Cornelia
Sindermann5,6, Christian Montag5, Xiaoxiao Zheng3,7, Hongxing Zhang2, Yao Shuxia1,3,
Zheng Wang8, Bo Zhou1, Keith M. Kendrick1,3, Benjamin Becker1,3*
1
The Center of Psychosomatic Medicine, Sichuan Provincial Center for Mental
Health, Provincial People's Hospital, University of Electronic Science and
Technology of China, Chengdu, China
2
School of Psychology, Xinxiang Medical University, Xinxiang, China
3
MOE Key Laboratory for Neuroinformation, School of Life Science and
Technology, University of Electronic Science and Technology of China, Chengdu,
China
4
School of Psychology and Cognitive Science, East China Normal University,
Shanghai, China
5
Department of Molecular Psychology, Institute of Psychology and Education, Ulm
University, Ulm, Germany
6
Interchange Forum for Reflecting on Intelligent Systems, University of Stuttgart,
Stuttgart, Germany
7
Brain Cognition and Brain Disease Institute (BCBDI), Shenzhen Institute of
Advanced Technology, Chinese Academy of Sciences, Shenzhen, China
8
School of Psychological and Cognitive Sciences, Beijing Key Laboratory of
Behavior and Mental Health, IDG/McGovern Institute for Brain Research, Peking.
Tsinghua Center for Life Sciences, Peking University, Beijing, China
*Correspondence to: Benjamin Becker (ben_becker@gmx.de)
NOTE: This preprint reports new research that has not been certified by peer review and should not be used to guide clinical practice.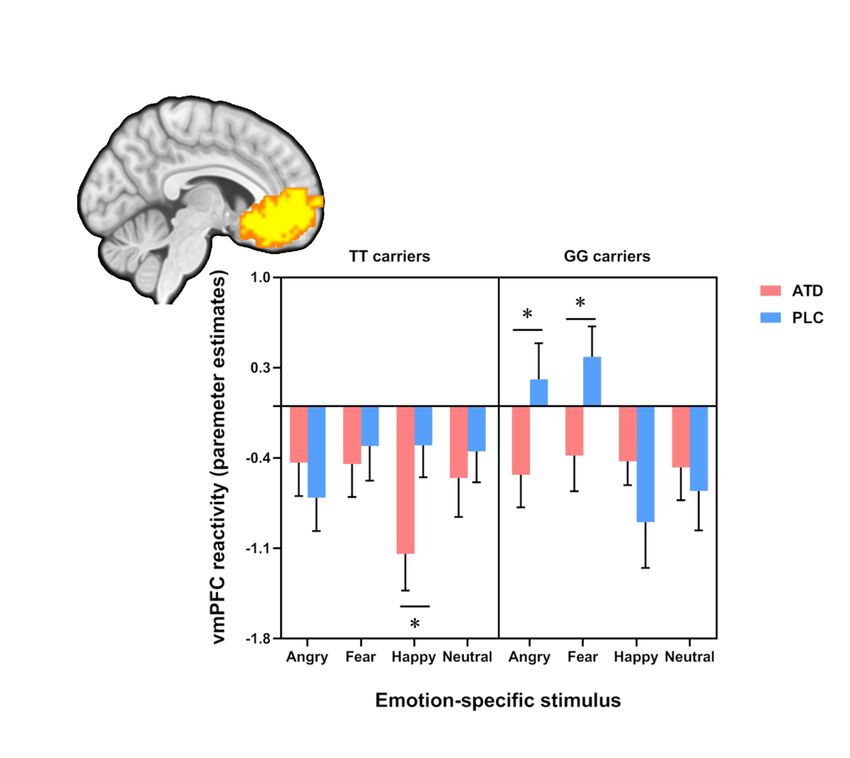
medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
Abstract
Serotonin (5-HT) has long been implicated in adaptive emotion regulation as well as
the development and treatment of emotional dysregulations in mental disorders.
Accumulating evidence suggests that a genetic vulnerability may render some
individuals at a greater risk for the detrimental effects of transient variations in 5-HT
signaling. The present study aimed to investigate whether individual variations in the
Tryptophan hydroxylase 2 (TPH2) genetics influence susceptibility for behavioral and
neural threat reactivity dysregulations during transiently decreased 5-HT signaling. To
this end, interactive effects between TPH2 (rs4570625) genotype and acute tryptophan
depletion (ATD) on reactivity towards angry, neutral and happy faces were examined
in a within-subject placebo-controlled pharmacological fMRI trial (n = 51). An a priori
genotype stratification approach of extreme groups (GG vs. TT) allowed balanced
sampling. While no main effects of ATD on neural reactivity to threat-related stimuli
and mood state were observed in the entire sample, accounting for TPH2 genotype
revealed an ATD-induced increase in subjective anxious arousal in the GG but not the
TT carriers. The effects were mirrored on the neural level, such that ATD specifically
reduced ventromedial prefrontal cortex (vmPFC) reactivity towards threat-related
stimuli in the GG carriers. Furthermore, the ATD-induced increase in subjective anxiety
positively associated with the extent of ATD-induced changes in vmPFC activity in
response to threat-related stimuli in GG carriers. Together the present findings suggest
for the first time that individual variations in TPH2 genetics render individuals
susceptible to the anxiogenic and neural effects of a transient decrease in 5-HT
signaling.
Keywords: serotonin; tryptophan; threat; anxiety; fear; depression; amygdala; PFC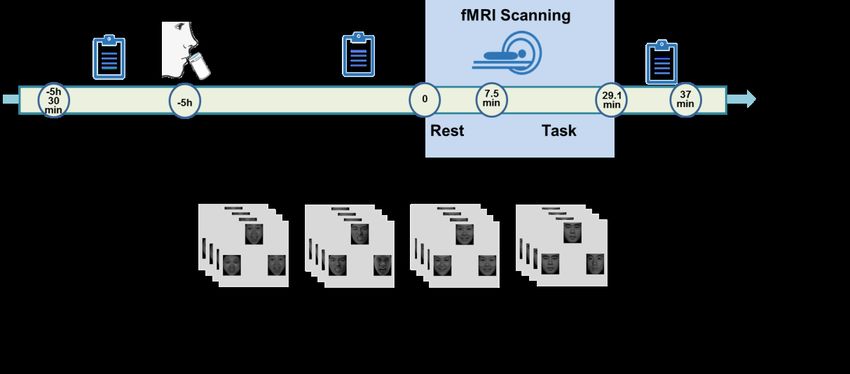
medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
Introduction
Serotonin (5-HT) has long been implicated in adaptive emotion regulation as well as
the development and treatment of emotional dysregulations in mental disorders, in
particular anxiety and mood disorders [1-4]. Most human research on the role of 5-HT
in these domains is based on determining behavioral and neural variations associated
with individual differences in central 5-HT signaling, including stable (genetic, trait-
like) variations as well as transient variations e.g. induced by a rapid decrease in central
serotonergic signaling via acute tryptophan depletion (ATD) procedures [5-7]. However,
recent quantitative and qualitative reviews challenge both the role of 5-HT genetics in
the etiology of emotional dysregulations in mood and anxiety disorders [4, 8] as well
as the anxiogenic effects of ATD and associated neural processes [9-11].
Mounting evidence suggests considerable individual variations in the emotional
effects of an ATD-induced transient reduction of central serotonergic signaling.
Individual variations in the effects of ATD have been particularly described in the
domains of fear/anxiety and executive control and linked to individual differences in 5-
HT genetics and associated personality traits [12-18]. These findings may point to an
underlying genetic vulnerability that renders some individuals at a greater risk for the
detrimental effects of transient variations in 5-HT signaling. However, the few previous
studies exclusively focused on genetic variations in the 5-HT transporter-linked
promoter region (5-HTTLPR), a polymorphism with unclear relevance for both, central
serotonergic signaling and psychopathological phenotypes [19, 20].
Central serotonergic signaling is directly regulated by the 5-HT biosynthesis ratemedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
limiting enzyme tryptophan hydroxylase 2 (TPH2) and variations in TPH2 genetics
have been robustly linked to serotonin synthesis rates in limbic and prefrontal systems
[21-23] and behavioral variations in emotional reactivity and regulation [24-26]. Initial
studies employing genetic imaging approaches furthermore reported that TPH2
rs4570625 single nucleotide polymorphism (SNP) variants affect emotional reactivity
in limbic regions and regulatory control in frontal regions [27-30]. The TPH2
rs4570625 SNP biases the reactivity of the amygdala in response to threatening stimuli
[31] and attenuates top-down regulatory control of the ventromedial prefrontal cortex
(vmPFC) [17]. Dysregulations in these regions may thus represent a candidate marker
that neurally reflects a TPH-2 variation-related vulnerability for the detrimental effects
of transient disruptions in 5-HT signaling.
Previous studies have revealed some - yet also inconsistent - evidence for effects
of transiently reduced 5-HT signaling induced by ATD on the amygdala-vmPFC
circuitry. ATD modulated communication in this circuitry and biased processing
towards aversive emotional stimuli [32, 33]. ATD moreover affected the activity of the
amygdala and the vmPFC during aversive stimuli processing [11, 34, 35], such that
emotionally salient stimuli yielded higher responses in the mPFC as a result of impaired
emotional evaluation following ATD [36, 37].
Despite accumulating evidence that interactions between genetic and
pharmacological variations in 5-HT signaling affect the neural mechanisms of emotion
processing, it remains unknown if individual variations the TPH2 gene polymorphisms
can render individuals vulnerable for the effects of a transient variation in 5-HTmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
signaling. The present study therefore examined whether the TPH2 SNP rs4570625
polymorphism determines the impact of an ATD-induced transient dysfunction in
serotonergic signaling on subjective anxiety and on threat-specific neural reactivity of
the vmPFC-amygdala circuitry (e.g. in responses to angry or fearful, but not happy
faces) using a pre-registered within-subject randomized placebo-controlled double-
blind fMRI experiment during which healthy male participants (n=51) either underwent
transient decreases in 5-HT signaling (via a previously validated orally administered
acute tryptophan depletion protocol, ATD) or a matched placebo protocol (PLC).
Effects on threat-related amygdala-vmPFC reactivity were determined using functional
magnetic resonance imaging (fMRI) during which angry faces and fearful faces (threat
condition) as well as neutral, happy and non-emotional control stimuli were presented.
First, we aimed to test whether the effect of diminished central 5-HT on threat-related
amygdala-vmPFC reactivity varied as a function of TPH2 gene polymorphisms. Second,
the relationship between ATD-induced effects on subjective anxious arousal and brain
activity changes induced by ATD was explored. In particular, we hypothesized an
interaction between the TPH2 genotype and ATD on subjective anxiety and amygdala-
vmPFC activity, which together may reflect a genetic vulnerability factor for the
development of emotional dysregulations in response to transient 5-HT changes.
Method
Participants
A total of 51 right-handed, healthy male participants (age M = 22.02 ± 2.48, 27 TT
carriers, 24 GG carriers) were enrolled. Please note that TT and GG carriers weremedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
invited after a priori genotyping to increase statistical power for the present experiment.
To reduce variance related to sex differences in 5-HT synthesis rates [38], only male
subjects were included. In the context of problematic effect size determination in
genetic imaging studies [39, 40] sample size was based on previous studies [41].
Individuals with a current or a history of medical, neurological/psychiatric disorders,
regular use of psychotropic substances or MRI contraindications were excluded. The
study was approved by the local ethics committee and in accordance with the latest
revision of the Declaration of Helsinki. Written informed consent was obtained and the
study was pre-registered on clinicaltrials.gov (https://clinicaltrials.gov/ct2/
show/NCT03549182, ID NCT03549182).
Genotyping
Genomic DNA was purified from buccal cells using a MagNA Pure96 robot (Roche
Diagnostics, Mannheim), sample probes were designed by TIB MolBiol (Berlin,
Germany). Genotyping was conducted by real-time polymerase chain reaction (RT-
PCR) and subsequent high-resolution melting on a Cobas Z480 Light Cycler (Roche
Diagnostics, Mannheim, Germany) according to the manufacturer’s instructions.
Serotonergic Challenge
Previous studies have demonstrated that after administration of ATD, tryptophan levels
continuously decrease until they reach a plateau after about 5 hours and that the robust
decrease lasts around two hours [6, 7, 42]. Given that tryptophan is the amino acid
precursor of serotonin, the ATD procedure induces a transient and selective reduction
in central serotonergic neurotransmission [5]. The contents of the amino-acid mixturesmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
were based on previously validated proportions [43]. The amino acid mixture (ATD)
consisted of 5.5g L-alanine, 4.9 g L-arginine, 2.7 g L-cystine, 3.2 g glycine, 3.2g L-
histidine, 8g L-isoleucine, 13.5 g L-leucine, 8.9 g L-lysine, 3.0 g L-methionine, 12.2 g
L-proline, 5.7 g L-phenylalanine, 6.9 g L-serine, 6.5 g L-threonine, 6.9 g L-tyrosine,
8.9 g L-valine, (total: 100 g). The control drink (PLC) contained identical ingredients
plus 2.3 g of L-tryptophan (total: 102.3 g). The drinks were prepared by stirring the
mixture into 200-ml water and lemon-lime flavor was added to mask the taste of the
mixture.
Control variables
To control for between-group differences (TT, GG) in anxiety, current stress, and early
life stress, the State-Trait Anxiety Inventory (STAI) [44], Perceived Stress Scale (PSS)
[45] and Childhood Trauma Questionnaire (CTQ) [46] were administered before
treatment administration. To assess effects of treatment on subjective anxiety levels the
State Anxiety Inventory component of the STAI (SAI) was repeatedly administered
before administration of the amino acid drink (T1), during transient 5-HT decrease
(immediately before MRI acquisition, T2) and at the end of the experiment (T3).
Procedure
The present study employed a with-subject randomized double-blind placebo-
controlled design. Participants were instructed to abstain from alcohol and caffeine for
24h and from food and drinks (except water) for 12h prior to the experiment. To adhere
to the pharmacodynamic profile of treatment, participants arrived between 7:30 to
10:00 AM and underwent fMRI acquisition between 13:00 to 15:30 PM. Upon arrival,medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
participants received a standardized protein-poor diet for breakfast. Following the
assessment of pre-treatment control variables, participants underwent a previously
validated tryptophan depletion protocol with ATD which has been demonstrated to lead
to a robust transient reduction in central 5HT signaling [7, 43, 47, 48] or PLC. The
ingestion of the amino acid mixtures was followed by a resting period of 5 hours to
achieve a robust reduction in tryptophan levels. During the resting period participants
were asked to relax and magazines were provided. Subsequently, participants
underwent the fMRI paradigm. Control variables were re-assessed immediately before
fMRI and after the emotion processing paradigms (at the end of the fMRI acquisition)
(schematic outline of the experimental protocols see Figure 1). All subjects were
administered both treatments (ATD, PLC) separated by approx. 5 weeks of wash-out
period.
Experimental paradigm
A validated blocked-design emotional face-matching paradigm was administered
during the two experimental sessions [49]. The paradigm consisted of 6 runs and every
run comprised 4 blocks of facial stimuli as well as 1 block of non-facial stimuli serving
as non-social control stimuli. During the face-processing blocks, a trio of condition-
specific (angry, fearful, happy or neutral expressions) facial stimuli was presented and
subjects were required to select one of the two faces (lower panel) that was identical to
a target face (upper panel). Each block comprised four condition-specific trials,
balanced for face gender. Asian facial stimuli were selected from a standardized Asian
facial expression database [50]. During the non-social control blocks a trio of simplemedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
geometric shapes (circles and ellipses) was presented and subjects were required to
select one of two shapes (lower panel) that matched the target shape presented in upper
panel (Figure 1B). Each control block comprised four different shape trios. All blocks
were preceded by a brief instruction (‘Face match’ or ‘Shape match’) that lasted 2s.
Within each block, each trial was presented for 4s with a variable interstimulus interval
(ISI) of 1-3 s (mean, 2s). The total block length was 24s and the total paradigm lasted
21min 36s.
MRI data acquisition and processing
MRI data were acquired on a 3.0 Tesla GE MR750 system (General Electric Medical
System, Milwaukee, WI, USA). To exclude subjects with apparent brain pathologies
and improve normalization of the functional time series, T1-weighted high-resolution
anatomical images were acquired with a spoiled gradient echo pulse sequence:
repetition time (TR), 5.9ms; echo time (TE), minimum; flip angle, 9°; field of view
(FOV), 256 × 256mm; acquisition matrix, 256 × 256; thickness, 1mm; number of slices,
156. For the functional MRI timeseries a total of 648 functional volumes were acquired
using a T2*-weighted Echo Planar Imaging (EPI) sequence (TR, 2000ms; TE, 30ms;
FOV, 220 × 220mm; flip angle, 90°; image matrix, 64 × 64; thickness/gap, 3.2/0mm;
43 axial slices with an interleaved ascending order).
Functional time-series were pre-processed using statistical parametric mapping
(SPM12; Wellcome Department of Cognitive Neurology, Institute of Neurology,
London, United Kingdom). For each subject and run the first seven volumes were
discarded to allow magnet-steady images. The remaining functional images were slice-medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
time corrected and realigned to the first image to correct for head motion. The
functional images were next co-registered to the T1-weighted structural images and
normalized to Montreal Neurological Institute (MNI) standard space using the
segmentation parameters obtained from segmenting the structural images and
interpolated at 3×3×3mm voxel size. Finally, the normalized images were spatially
smoothed with an 8-mm full-width at half maximum (FWHM) Gaussian filter. For
statistical analysis a two-step general linear model (GLM) approach was employed. At
the single subject level an event-related general linear model (GLM) was employed
including condition-specific regressors modelling the five experimental conditions, the
instruction period and the six head motion parameters. Regressors for the experimental
conditions and instruction were convolved with the default SPM hemodynamic
response function (HRF). The design matrices additionally included a high pass filter
to control for low frequency components and a first-order autoregressive model (AR[1])
to account for autocorrelation in the time-series.
Region of Interest (ROI) selection and statistical thresholding
Based on our regional specific hypotheses derived from previous work [51-55], the
analyses specifically focused on the amygdala and vmPFC. The amygdala ROIs were
derived from Automatic Anatomical Labelling (AAL) [56]. The vmPFC ROI was
defined based on an automated meta-analysis of 199 studies using NeuroSynth
(www.neurosynth.org) with the search term “vmPFC” and thresholded at pFDRmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
extracted beta estimates from the vmPFC, left and right amygdala was employed. To
determine emotion-specific interactions between genotype and treatment the extracted
estimates were subjected to three-way ANOVAs (genotype×treatment×emotion)
examining the effect for each ROI separately. The p values for the analysis were
correspondingly adapted to p < 0.017 (0.05/3, corrected for three regions): (2) To
facilitate a more specific localization of the effects, a voxel-wise small volume
corrected analysis for the vmPFC and entire bilateral amygdala was additionally
employed. To this end, a peak-level family-wise error (FWE) small volume correction
was applied to a single mask encompassing the vmPFC and bilateral amygdala (total
volume of the mask, 33912 mm3) (FWE, p < 0.05). Behavioral and neural (extracted
parameter estimates) indices were further analyzed using SPSS 22.0 with appropriate
ANOVA models and Bonferroni corrected post-hoc tests. P < 0.05 two-tailed was
considered significant. Partial eta squared (h2p) and Cohen’s d were computed for as
measures of effect size.
Results
Sample characteristics, confounders, genetic and treatment effects on subjective
anxiety levels
The genotype groups did not differ with respect to pre-treatment control variables
including age, trait anxiety, current stress, childhood trauma exposure and pre-treatment
anxiety levels (all ps > 0.24, details see Table 1). Examining effects of treatment and
genotype on state anxiety during the course of the experiment by means of mixed
ANOVA models with genotype (TT vs. GG) as between-subject factor, and treatmentmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
(ATD vs. PLC) and timepoint (T1-T3; pre-oral administration, pre-fMRI, post-fMRI,
T1-T3) as within-subject factors revealed a interaction effect between genotype and
amino acid mixture (F1,46 = 4.67, p = 0.036, h2p = 0.092), with post-hoc analyses
suggesting that in GG carriers, participants reported higher anxious arousal levels
during transient 5-HT decreases as compared to the PLC session (p = 0.013, Figure 2).
Further exploratory analysis showed that the interaction effect was driven by a
significant increase in state anxiety levels in the GG group during transient 5-HT
decrease as compared to the PLC session following the threat exposure paradigm (T3;
t = 2.83, p = 0.01). No other significant main and interaction effects were observed.
Behavioral results
Examining accuracy (ACC) and response times (RT) during the fMRI paradigm by
means of mixed ANOVAs with emotion (angry vs. fearful vs. happy vs. neutral) and
treatment (ATD vs. PLC) as within-subject factors and genotype (TT vs. GG) as
between-subject factor revealed a significant main effect of emotion on both ACC (F =
15.93, p < 0.001, h2p = 0.245) and RT (F = 115.58, p < 0.001, h2p = 0.702). For the
ACC, post-hoc analysis revealed that, all participants responded more accurately for
angry (0.984±0.004) and fearful faces (0.980±0.003) as compared to happy
(0.959±0.006) and neutral faces (0.956±0.007, all ps < 0.001), reflecting attentive task
engagement irrespective of treatment. In addition, post-hoc analyses revealed that RTs
for fearful face (1038.24 ± 23.77) were faster compared to angry faces (1120.72 ±
26.57), angry faces compared to happy faces (1186.84 ± 34.23) and slower RT for
neutral faces (1272.39 ± 33.75) compared to all other conditions (all ps < 0.001).medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
Importantly, we observed a significant main effect of treatment (F = 5.81, p = 0.02, h2p
= 0.106) suggesting that participants made generally slower responses in the ATD
session as compared to PLC sessions (p = 0.02).
Neural activity: ROI analysis - three-way interaction in vmPFC and amygdala to face
emotions
Examination of the extracted parameter estimates from the vmPFC, left and right
amygdala revealed a significant three-way interaction in the ROI-specific ANOVAs for
both the vmPFC (F3,147 = 4.58; p = 0.007; h2p = 0.085; power (1 -β) = 0.838) and right
amygdala (F3,147 = 4.40; p = 0.006; h2p = 0.082; power (1 - β) = 0.85), but not the left
amygdala (p = 0.087). Post-hoc analyses on the extracted beta values from the vmPFC
mask revealed that in the GG carriers, ATD treatment decreased vmPFC reactivity to
fearful and angry face relative to the corresponding conditions under PLC treatment
(fearful: p = 0.034, angry: p = 0.043), while for the TT carriers, ATD treatment
suppressed vmPFC reactivity to happy face relative to the PLC treatment. Post-hoc
analyses on the extracted beta values from the right amygdala mask revealed that during
the PLC sessions, TT carriers showed higher reactivity both to happy face (p = 0.039)
and neutral face (p = 0.037) as compared to GG carriers, but when participants received
ATD treatment, there was no significant difference between GG and TT carriers (see
Figure 3).
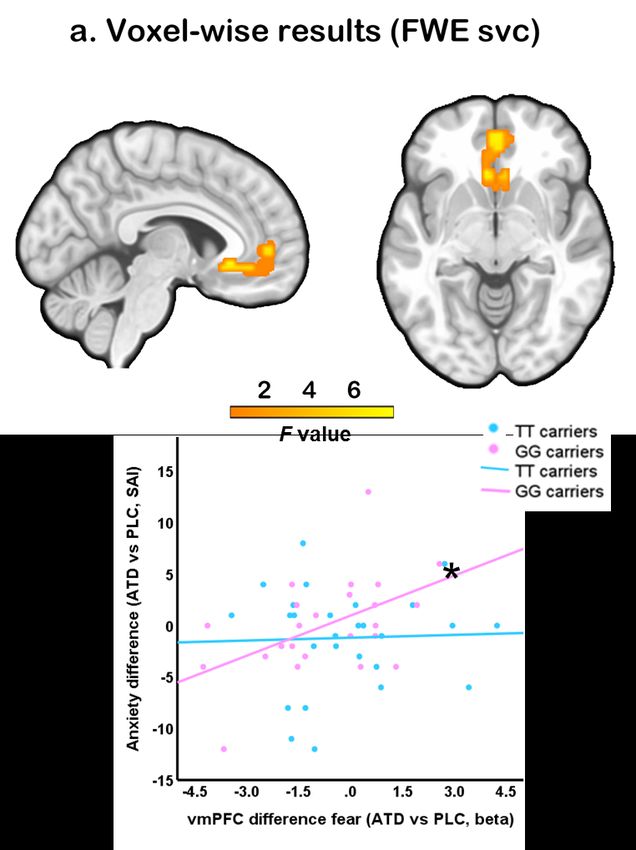
To facilitate a more specific localization of the effects, a voxel-wise small volume
corrected analysis for the vmPFC and entire bilateral amygdala was additionally
employed. This analysis revealed a significant three-way interaction effect in themedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
vmPFC after small volume correction for the mask encompassing the vmPFC and entire
amygdala (k = 103, pFWE = 0.031, F3, 343 = 7.47, peak MNI co-ordinate: x = -6, y = 27,
z = −6) (Fig. 4a), while no significant interaction effect was found in the right amygdala
(pFWE = 0.200). Thus, the interaction effect between genotype, treatment and emotion
in the vmPFC appears to be robust, while interaction in right amygdala was less robust.
Associations between subjective anxious arousal and neural activity
To explore the relationship between treatment-induced changes on the level of
subjective experience and neural activity (T2: pre-fMRI) an exploratory correlation
analysis was conducted. Results indicated a significant positive correlation between
state anxiety level changes and vmPFC activity changes induced by treatment (r = 0.502,
p = 0.013; Figure 4b) in GG carriers, while not in TT carriers (r = 0.035, p = 0.860,
between group correlation difference t = 1.85, p = 0.065). No significant associations
were observed for the other face conditions (all ps>0.05).
Discussion
The present study aimed at determining whether individual differences in a genetic
polymorphism with a regulatory role over central serotonin synthesis rates (TPH2)
influence susceptibility to the effects of a transient decrease in 5-HT signaling on
subjective anxiety and threat-related neural activity. To this end interactions between
TPH2 genotype and the effects of ATD were examined in a pre-registered within-
subject placebo-controlled pharmacological fMRI trial. Partly resembling previous
findings, the present study did not reveal significant effects of ATD on subjective
anxiety or neural reactivity to threat-related stimuli in the entire (pooled) sample [10,medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
57, 58]. However, when accounting for genetic differences significant interaction
effects between TPH2 genotype and ATD were observed, such that specifically the GG
carriers reported increased anxious arousal after ATD, whereas TT carriers reported no
significant changes during transiently decreased 5-HT signaling. The genotype-specific
anxiogenic effects were mirrored on the neural level, such that ATD specifically
reduced vmPFC reactivity towards threat-related stimuli in the GG carriers. In contrast,
in TT carriers ATD – relative to PLC – induced a reduced reactivity to happy faces in
the vmPFC. Finally, in GG carriers the ATD-induced increase in subjective anxiety was
positively associated with the extent of ATD-induced changes in vmPFC activity in
response to fearful faces following ATD. Together the present findings demonstrate for
the first time that individual variations in TPH2 genetics render healthy participants
susceptible to the anxiogenic and neural effects of a transient decrease in 5-HT
signaling.
The relevance of the serotonergic system for anxiety-related processes has long
been proposed and has been supported by several findings [59-62]. Several of these
studies have capitalized on a validated ATD procedure which allows a reliable and
transient decrease in central serotonin synthesis rates. While ATD reliably decreases the
availability of tryptophan - the precursor of serotonin - its anxiogenic effects have been
recently questioned in a comprehensive meta-analysis [10] and accumulating evidence
suggests strong individual differences [12-14, 16, 18, 63]. In line with our hypothesis
for a genetically determined susceptibility to the subjective emotional effects of ATD
we found that specifically GG carriers experienced anxiogenic effects. Although themedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
neurobiological implications of the TPH2 rs4570625 polymorphism are not fully
understood, the G-allele has been suggested to be related to a hypofunction of
tryptophan hydroxylase [64, 65] and in turn lower 5-HT synthesis rates which may
promote a stronger impact of a transient decrease in tryptophan.
Consistent with our hypothesis, the susceptibility on the behavioral level was
paralleled by a threat-specific interaction effect in the vmPFC-amygdala circuits.
Specifically in GG carriers ATD induced a decreased vmPFC-response to threat-related
stimuli, i.e. fearful and angry faces. Animal model and human studies demonstrated that
serotonergic fibers project into the limbic system and cortical regions, including the
vmPFC [52, 66], suggesting that 5-HT regulates neural activity in these circuits.
In the context of anxiety-related domains the vmPFC plays a critical role in both
motivational processes to minimize threat [67-69] and implicit emotion regulation of
threat, i.e. during extinction [53, 70]. Previous studies in individuals with pathological
anxiety have reported deficient engagement of the vmPFC, including decreased vmPFC
engagement accompanied by deficient differentiation of safety from threat [71] in
generalized anxiety disorder or deficient extinction recall across anxiety disorders [72].
The associated deficits in safety-threat differentiation may in turn bias both the
perception of information as threatening [73] and an adaptive regulatory control over
threat. In line with this proposed role of the vmPFC, non-invasive stimulation of this
region attenuates the attentional bias to threat-related stimuli in the dot-probe test [67]
and facilitates adaptive threat extinction [74]. The observation of ATD-induced
decreased vmPFC reactivity to threat-related stimuli associated with the extent ofmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
treatment-induced increased anxiety levels in the GG carriers may thus reflect either an
enhanced detection of threat or a reduced engagement of implicit top-down fear
regulation, both of which have been associated with higher anxious arousal [53, 75].
In contrast to our hypothesis, no robust treatment and genotype interaction effects
were observed on the mean amplitude of amygdala threat reactivity. These findings may
underscore that the genetic vulnerability to ATD renders prefrontal regulatory regions
but not the amygdala - which is more involved in bottom-up threat reactivity –
susceptible to transient serotonergic hypoactivity. Moreover, the amygdala shows a
rapid adaptation to repeated presentations with both reduced [76-78] and increased
reactivity [54, 79-81] being reported, which may have overshadowed determination of
robust effect. In addition, previous evidence between amygdala reactivity and 5-HT
functioning is inconsistent, withsome studies failing to find associations between
general amygdala reactivity and 5-HT genetics as well as robust effects of ATD on
amygdala activity [9, 82, 83].
Given the inconsistent results on associations between serotonergic functioning
and disorders characterized by emotional regulation deficits the present findings have
additional clinical implications. First, transient changes in tryptophan and serotonin
levels have been associated with different environmental factors such as psychosocial
stress, inflammation, and dietary habits [84], and the present results indicate that
particular individuals with a specific TPH2 genotype are vulnerable to the detrimental
effects on anxiety and neural activity. The underlying changes may represent a core
pathological mechanism for the development and maintenance of exaggerated anxiousmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
arousal which can promote emotional dysregulations and mental disorders. Second,
serotonin dysfunction has been suggested as a candidate biomarker for exaggerated
anxiety [85] and promising treatment target for anxiety disorders. The present results
indicate that effects of serotonin-modulation on anxiety and associated vmPFC threat
responses depend on individual variation in the TPH2 gene. This can lead to individual
variations in the initial response to treatments targeting serotonin transmission,
including selective serotonin reuptake inhibitors (SSRIs) frequently prescribed for
anxiety and mood disorders.
Findings of the present study need to be considered in the context of the following
limitations. First, only male subjects were enrolled to reduce variance in the data related
to sex differences in 5-HT synthesis [38]. Future studies need to determine whether the
observed effects generalize to women. Second, considering that drawing blood samples
might increase anxiety levels and thus can confound our primary outcomes tryptophan
blood levels were not assessed. However, previous studies reported robust and selective
decreases in 5-HT signaling following similar the ATD treatment protocols as used in
the present study [43, 86], the additional examination of blood-level measures
particularly in the combined treatment group may reveal important additional
information on the relevance of serotonergic system to anxiety-relevant processes and
should be included in future studies. The present study matched the groups for trait
anxiety, while a number of studies reported associations with trait anxiety impact
cognitive and emotional processes as well as pharmacological effects [58, 87, 88].
Finally, while the study involved a total of 51 participants in a complex pharmaco-fMRImedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
design we did not include an a priori sample size calculation. The priori calculation for
sample size and power in complex genetics-pharmaco-fMRI design is still limited
although our sample size was based on previous studies [41]. Although the ROI analysis
may have facilitated a more robust determination of effects validation and replication
designs are needed [89, see also 23].
Together, the present findings provide the first evidence that individual variations
in a TPH2 polymorphism – a genetic variation functionally related to central 5-HT
signaling – render individuals susceptible for the anxiogenic and threat-specific neural
effects of transient variations in serotonergic signaling. They may also explain the
previous inconsistent findings on the effects of acute tryptophan depletion in healthy
individuals and point to a vulnerability marker for conditions characterized by
excessive anxiety.medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
Acknowledgements
This work was supported by the China MOST2030 Brain Project (Grant No.
2022ZD0208500), National Key Research and Development Program of China (Grant
No. 2018YFA0701400) and the National Natural Science Foundation of China
(Grants No. 32250610208, 82271583)
Author contributions
CL and BB designed the study. CL, KL, XZ, MF, CS, YZ conducted the experiment
and collected the data. CL performed data analyses. CL and BB wrote the manuscript.
HZ, YS, BZ, CM, ZW and KK revised the manuscript draft.
Data availability
Unthresholded group-level statistical maps are available on NeuroVault
(https://neurovault.org/images/794826/), additional data related to study is available
from the corresponding author upon reasonable request. A preprint of the manuscript
had been archived on the biorxiv.org repository.medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
References
1. Cools R, Roberts AC, Robbins TW. Serotoninergic regulation of emotional and behavioural control
processes. Trends Cogn Sci. 2008;12(1):31-40. doi: 10.1016/j.tics.2007.10.011.
2. Hood SD, Bell CJ, Argyropoulos SV, Nutt DJ. Don't panic. A guide to tryptophan depletion with
disorder-specific anxiety provocation. J Psychopharmacol. 2016;30(11):1137-1140. doi:
10.1177/0269881116655321.
3. Kraus C, Castrén E, Kasper S, Lanzenberger R. Serotonin and neuroplasticity - Links between
molecular, functional and structural pathophysiology in depression. Neurosci Biobehav Rev.
2017;77:317-326. doi: 10.1016/j.neubiorev.2017.03.007.
4. Moncrieff J, Cooper RE, Stockmann T, Amendola S, Hengartner MP, Horowitz MA. The serotonin
theory of depression: a systematic umbrella review of the evidence. Mol Psychiatry. 2022. doi:
10.1038/s41380-022-01661-0.
5. Crockett MJ, Clark L, Roiser JP, Robinson OJ, Cools R, Chase HW, et al. Converging evidence for
central 5-HT effects in acute tryptophan depletion. Mol Psychiatry. 2012;17(2):121-123. doi:
10.1038/mp.2011.106.
6. Young SN, Smith SE, Pihl RO, Ervin FR. Tryptophan depletion causes a rapid lowering of mood in
normal males. Psychopharmacology (Berl). 1985;87(2):173-177. doi: 10.1007/bf00431803.
7. Williams WA, Shoaf SE, Hommer D, Rawlings R, Linnoila M. Effects of acute tryptophan depletion
on plasma and cerebrospinal fluid tryptophan and 5-hydroxyindoleacetic acid in normal volunteers. J
Neurochem. 1999;72(4):1641-1647. doi: 10.1046/j.1471-4159.1999.721641.x.
8. Ottenhof KW, Sild M, Lévesque ML, Ruhé HG, Booij L. TPH2 polymorphisms across the spectrum
of psychiatric morbidity: A systematic review and meta-analysis. Neurosci Biobehav Rev. 2018;92:29-42.
doi: 10.1016/j.neubiorev.2018.05.018.
9. Raab K, Kirsch P, Mier D. Understanding the impact of 5-HTTLPR, antidepressants, and acute
tryptophan depletion on brain activation during facial emotion processing: A review of the imaging
literature. Neurosci Biobehav Rev. 2016;71:176-197. doi: 10.1016/j.neubiorev.2016.08.031.
10. Schopman SME, Bosman RC, Muntingh ADT, van Balkom A, Batelaan NM. Effects of tryptophan
depletion on anxiety, a systematic review. Transl Psychiatry. 2021;11(1):118. doi: 10.1038/s41398-021-
01219-8.
11. Fusar-Poli P, Allen P, McGuire P, Placentino A, Cortesi M, Perez J. Neuroimaging and
electrophysiological studies of the effects of acute tryptophan depletion: a systematic review of the
literature. Psychopharmacology (Berl). 2006;188(2):131-143. doi: 10.1007/s00213-006-0493-1.
12. Kanen JW, Arntz FE, Yellowlees R, Cardinal RN, Price A, Christmas DM, et al. Serotonin depletion
amplifies distinct human social emotions as a function of individual differences in personality. Transl
Psychiatry. 2021;11(1):81. doi: 10.1038/s41398-020-00880-9.
13. Kanen JW, Arntz FE, Yellowlees R, Christmas DM, Price A, Apergis-Schoute AM, et al. Effect of
Tryptophan Depletion on Conditioned Threat Memory Expression: Role of Intolerance of Uncertainty.
Biol Psychiatry Cogn Neurosci Neuroimaging. 2021;6(5):590-598. doi: 10.1016/j.bpsc.2020.12.012.
14. Kanen JW, Apergis-Schoute AM, Yellowlees R, Arntz FE, van der Flier FE, Price A, et al. Serotonin
depletion impairs both Pavlovian and instrumental reversal learning in healthy humans. Mol Psychiatry.
2021;26(12):7200-7210. doi: 10.1038/s41380-021-01240-9.
15. Finger EC, Marsh AA, Buzas B, Kamel N, Rhodes R, Vythilingham M, et al. The impact of tryptophan
depletion and 5-HTTLPR genotype on passive avoidance and response reversal instrumental learningmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
tasks. Neuropsychopharmacology. 2007;32(1):206-215. doi: 10.1038/sj.npp.1301182.
16. Marsh AA, Finger EC, Buzas B, Soliman N, Richell RA, Vythilingham M, et al. Impaired recognition
of fear facial expressions in 5-HTTLPR S-polymorphism carriers following tryptophan depletion.
Psychopharmacology (Berl). 2006;189(3):387-394. doi: 10.1007/s00213-006-0581-2.
17. Neufang S, Akhrif A, Herrmann CG, Drepper C, Homola GA, Nowak J, et al. Serotonergic modulation
of 'waiting impulsivity' is mediated by the impulsivity phenotype in humans. Transl Psychiatry.
2016;6(11):e940. doi: 10.1038/tp.2016.210.
18. Walderhaug E, Magnusson A, Neumeister A, Lappalainen J, Lunde H, Refsum H, et al. Interactive
effects of sex and 5-HTTLPR on mood and impulsivity during tryptophan depletion in healthy people.
Biol Psychiatry. 2007;62(6):593-599. doi: 10.1016/j.biopsych.2007.02.012.
19. Munafò MR, Freimer NB, Ng W, Ophoff R, Veijola J, Miettunen J, et al. 5-HTTLPR genotype and
anxiety-related personality traits: a meta-analysis and new data. Am J Med Genet B Neuropsychiatr
Genet. 2009;150b(2):271-281. doi: 10.1002/ajmg.b.30808.
20. Culverhouse RC, Saccone NL, Horton AC, Ma Y, Anstey KJ, Banaschewski T, et al. Collaborative meta-
analysis finds no evidence of a strong interaction between stress and 5-HTTLPR genotype contributing
to the development of depression. Mol Psychiatry. 2018;23(1):133-142. doi: 10.1038/mp.2017.44.
21. Booij L, Turecki G, Leyton M, Gravel P, Lopez De Lara C, Diksic M, et al. Tryptophan hydroxylase(2)
gene polymorphisms predict brain serotonin synthesis in the orbitofrontal cortex in humans. Mol
Psychiatry. 2012;17(8):809-817. doi: 10.1038/mp.2011.79.
22. Furmark T, Marteinsdottir I, Frick A, Heurling K, Tillfors M, Appel L, et al. Serotonin synthesis rate
and the tryptophan hydroxylase-2: G-703T polymorphism in social anxiety disorder. J Psychopharmacol.
2016;30(10):1028-1035. doi: 10.1177/0269881116648317.
23. Liu C, Xu L, Li J, Zhou F, Yang X, Zheng X, et al. Serotonin and early life stress interact to shape brain
architecture and anxious avoidant behavior - a TPH2 imaging genetics approach. Psychol Med.
2021;51(14):2476-2484. doi: 10.1017/s0033291720002809.
24. Gutknecht L, Jacob C, Strobel A, Kriegebaum C, Müller J, Zeng Y, et al. Tryptophan hydroxylase-2
gene variation influences personality traits and disorders related to emotional dysregulation. Int J
Neuropsychopharmacol. 2007;10(3):309-320. doi: 10.1017/s1461145706007437.
25. Laas K, Kiive E, Mäestu J, Vaht M, Veidebaum T, Harro J. Nice guys: Homozygocity for the TPH2 -
703G/T (rs4570625) minor allele promotes low aggressiveness and low anxiety. J Affect Disord.
2017;215:230-236. doi: 10.1016/j.jad.2017.03.045.
26. Reuter M, Kuepper Y, Hennig J. Association between a polymorphism in the promoter region of
the TPH2 gene and the personality trait of harm avoidance. Int J Neuropsychopharmacol.
2007;10(3):401-404. doi: 10.1017/s1461145706007073.
27. Reuter M, Esslinger C, Montag C, Lis S, Gallhofer B, Kirsch P. A functional variant of the tryptophan
hydroxylase 2 gene impacts working memory: a genetic imaging study. Biol Psychol. 2008;79(1):111-
117. doi: 10.1016/j.biopsycho.2007.12.002.
28. Kennedy AP, Binder EB, Bowman D, Harenski K, Ely T, Cisler JM, et al. A common TPH2 haplotype
regulates the neural processing of a cognitive control demand. Am J Med Genet B Neuropsychiatr Genet.
2012;159b(7):829-840. doi: 10.1002/ajmg.b.32090.
29. Canli T, Congdon E, Gutknecht L, Constable RT, Lesch KP. Amygdala responsiveness is modulated by
tryptophan hydroxylase-2 gene variation. J Neural Transm (Vienna). 2005;112(11):1479-1485. doi:
10.1007/s00702-005-0391-4.
30. Canli T, Congdon E, Todd Constable R, Lesch KP. Additive effects of serotonin transporter andmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
tryptophan hydroxylase-2 gene variation on neural correlates of affective processing. Biol Psychol.
2008;79(1):118-125. doi: 10.1016/j.biopsycho.2008.01.004.
31. Brown SM, Peet E, Manuck SB, Williamson DE, Dahl RE, Ferrell RE, et al. A regulatory variant of the
human tryptophan hydroxylase-2 gene biases amygdala reactivity. Mol Psychiatry. 2005;10(9):884-888,
805. doi: 10.1038/sj.mp.4001716.
32. Passamonti L, Crockett MJ, Apergis-Schoute AM, Clark L, Rowe JB, Calder AJ, et al. Effects of acute
tryptophan depletion on prefrontal-amygdala connectivity while viewing facial signals of aggression.
Biol Psychiatry. 2012;71(1):36-43. doi: 10.1016/j.biopsych.2011.07.033.
33. Klaassen T, Riedel WJ, Deutz NE, Van Praag HM. Mood congruent memory bias induced by
tryptophan depletion. Psychol Med. 2002;32(1):167-172. doi: 10.1017/s003329170100438x.
34. Daly E, Deeley Q, Hallahan B, Craig M, Brammer M, Lamar M, et al. Effects of acute tryptophan
depletion on neural processing of facial expressions of emotion in humans. Psychopharmacology (Berl).
2010;210(4):499-510. doi: 10.1007/s00213-010-1850-7.
35. Helmbold K, Zvyagintsev M, Dahmen B, Bubenzer-Busch S, Gaber TJ, Crockett MJ, et al. Effects of
serotonin depletion on punishment processing in the orbitofrontal and anterior cingulate cortices of
healthy women. Eur Neuropsychopharmacol. 2015;25(6):846-856. doi:
10.1016/j.euroneuro.2015.02.007.
36. Roiser JP, Levy J, Fromm SJ, Nugent AC, Talagala SL, Hasler G, et al. The effects of tryptophan
depletion on neural responses to emotional words in remitted depression. Biol Psychiatry.
2009;66(5):441-450. doi: 10.1016/j.biopsych.2009.05.002.
37. Robinson OJ, Overstreet C, Allen PS, Letkiewicz A, Vytal K, Pine DS, et al. The role of serotonin in
the neurocircuitry of negative affective bias: serotonergic modulation of the dorsal medial prefrontal-
amygdala 'aversive amplification' circuit. Neuroimage. 2013;78:217-223. doi:
10.1016/j.neuroimage.2013.03.075.
38. Nishizawa S, Benkelfat C, Young SN, Leyton M, Mzengeza S, de Montigny C, et al. Differences
between males and females in rates of serotonin synthesis in human brain. Proc Natl Acad Sci U S A.
1997;94(10):5308-5313. doi: 10.1073/pnas.94.10.5308.
39. Carter CS, Bearden CE, Bullmore ET, Geschwind DH, Glahn DC, Gur RE, et al. Enhancing the
Informativeness and Replicability of Imaging Genomics Studies. Biol Psychiatry. 2017;82(3):157-164. doi:
10.1016/j.biopsych.2016.08.019.
40. Avinun R, Nevo A, Knodt AR, Elliott ML, Hariri AR. Replication in Imaging Genetics: The Case of
Threat-Related Amygdala Reactivity. Biol Psychiatry. 2018;84(2):148-159. doi:
10.1016/j.biopsych.2017.11.010.
41. Eisner P, Klasen M, Wolf D, Zerres K, Eggermann T, Eisert A, et al. Cortico-limbic connectivity in
MAOA-L carriers is vulnerable to acute tryptophan depletion. Hum Brain Mapp. 2017;38(3):1622-1635.
doi: 10.1002/hbm.23475.
42. Carpenter LL, Anderson GM, Pelton GH, Gudin JA, Kirwin PD, Price LH, et al. Tryptophan depletion
during continuous CSF sampling in healthy human subjects. Neuropsychopharmacology. 1998;19(1):26-
35. doi: 10.1016/s0893-133x(97)00198-x.
43. Moreno FA, Parkinson D, Palmer C, Castro WL, Misiaszek J, El Khoury A, et al. CSF neurochemicals
during tryptophan depletion in individuals with remitted depression and healthy controls. Eur
Neuropsychopharmacol. 2010;20(1):18-24. doi: 10.1016/j.euroneuro.2009.10.003.
44. Spielberger CD. Manual for the State-Trait Anxiety Inventory. Self Evaluation Questionnaire. 1970.
45. Cohen S, Kamarck T, Mermelstein R. A global measure of perceived stress. J Health Soc Behav.medRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
1983;24(4):385-396.
46. Bernstein DP, Stein JA, Newcomb MD, Walker E, Pogge D, Ahluvalia T, et al. Development and
validation of a brief screening version of the Childhood Trauma Questionnaire. Child Abuse Negl.
2003;27(2):169-190. doi: 10.1016/s0145-2134(02)00541-0.
47. Salomon RM, Cowan RL, Rogers BP, Dietrich MS, Bauernfeind AL, Kessler RM, et al. Time series
fMRI measures detect changes in pontine raphé following acute tryptophan depletion. Psychiatry Res.
2011;191(2):112-121. doi: 10.1016/j.pscychresns.2010.10.007.
48. Yatham LN, Liddle PF, Sossi V, Erez J, Vafai N, Lam RW, et al. Positron emission tomography study
of the effects of tryptophan depletion on brain serotonin(2) receptors in subjects recently remitted from
major depression. Arch Gen Psychiatry. 2012;69(6):601-609. doi: 10.1001/archgenpsychiatry.2011.1493.
49. Hariri AR, Tessitore A, Mattay VS, Fera F, Weinberger DR. The amygdala response to emotional
stimuli: a comparison of faces and scenes. Neuroimage. 2002;17(1):317-323. doi:
10.1006/nimg.2002.1179.
50. Gong X, Huang YX, Wang Y, Luo YJ. Revision of the Chinese facial affective picture system. Chinese
Mental Health Journal. 2011.
51. Jackson ME, Moghaddam B. Amygdala regulation of nucleus accumbens dopamine output is
governed by the prefrontal cortex. J Neurosci. 2001;21(2):676-681. doi: 10.1523/jneurosci.21-02-
00676.2001.
52. Challis C, Berton O. Top-Down Control of Serotonin Systems by the Prefrontal Cortex: A Path toward
Restored Socioemotional Function in Depression. ACS Chem Neurosci. 2015;6(7):1040-1054. doi:
10.1021/acschemneuro.5b00007.
53. Zhou F, Geng Y, Xin F, Li J, Feng P, Liu C, et al. Human Extinction Learning Is Accelerated by an
Angiotensin Antagonist via Ventromedial Prefrontal Cortex and Its Connections With Basolateral
Amygdala. Biol Psychiatry. 2019;86(12):910-920. doi: 10.1016/j.biopsych.2019.07.007.
54. Liu C, Lan C, Li K, Zhou F, Yao S, Xu L, et al. Oxytocinergic Modulation of Threat-Specific Amygdala
Sensitization in Humans Is Critically Mediated by Serotonergic Mechanisms. Biol Psychiatry Cogn
Neurosci Neuroimaging. 2021;6(11):1081-1089. doi: 10.1016/j.bpsc.2021.04.009.
55. Mihov Y, Kendrick KM, Becker B, Zschernack J, Reich H, Maier W, et al. Mirroring fear in the absence
of a functional amygdala. Biol Psychiatry. 2013;73(7):e9-11. doi: 10.1016/j.biopsych.2012.10.029.
56. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N, et al. Automated
anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI
single-subject brain. Neuroimage. 2002;15(1):273-289. doi: 10.1006/nimg.2001.0978.
57. Cools R, Calder AJ, Lawrence AD, Clark L, Bullmore E, Robbins TW. Individual differences in threat
sensitivity predict serotonergic modulation of amygdala response to fearful faces. Psychopharmacology
(Berl). 2005;180(4):670-679. doi: 10.1007/s00213-005-2215-5.
58. Hornboll B, Macoveanu J, Nejad A, Rowe J, Elliott R, Knudsen GM, et al. Neuroticism predicts the
impact of serotonin challenges on fear processing in subgenual anterior cingulate cortex. Sci Rep.
2018;8(1):17889. doi: 10.1038/s41598-018-36350-y.
59. Gordon JA, Hen R. The serotonergic system and anxiety. Neuromolecular Med. 2004;5(1):27-40.
doi: 10.1385/nmm:5:1:027.
60. Frick A, Åhs F, Engman J, Jonasson M, Alaie I, Björkstrand J, et al. Serotonin Synthesis and Reuptake
in Social Anxiety Disorder: A Positron Emission Tomography Study. JAMA Psychiatry. 2015;72(8):794-
802. doi: 10.1001/jamapsychiatry.2015.0125.
61. Baldwin DS, Anderson IM, Nutt DJ, Allgulander C, Bandelow B, den Boer JA, et al. Evidence-basedmedRxiv preprint doi: https://doi.org/10.1101/2023.03.18.23287402; this version posted March 20, 2023. The copyright holder for this
preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in
perpetuity.
It is made available under a CC-BY-NC 4.0 International license .
pharmacological treatment of anxiety disorders, post-traumatic stress disorder and obsessive-
compulsive disorder: a revision of the 2005 guidelines from the British Association for
Psychopharmacology. J Psychopharmacol. 2014;28(5):403-439. doi: 10.1177/0269881114525674.
62. Bandelow B, Sher L, Bunevicius R, Hollander E, Kasper S, Zohar J, et al. Guidelines for the
pharmacological treatment of anxiety disorders, obsessive-compulsive disorder and posttraumatic
stress disorder in primary care. Int J Psychiatry Clin Pract. 2012;16(2):77-84. doi:
10.3109/13651501.2012.667114.
63. Neumeister A, Hu XZ, Luckenbaugh DA, Schwarz M, Nugent AC, Bonne O, et al. Differential effects
of 5-HTTLPR genotypes on the behavioral and neural responses to tryptophan depletion in patients with
major depression and controls. Arch Gen Psychiatry. 2006;63(9):978-986. doi:
10.1001/archpsyc.63.9.978.
64. Lin YM, Chao SC, Chen TM, Lai TJ, Chen JS, Sun HS. Association of functional polymorphisms of the
human tryptophan hydroxylase 2 gene with risk for bipolar disorder in Han Chinese. Arch Gen Psychiatry.
2007;64(9):1015-1024. doi: 10.1001/archpsyc.64.9.1015.
65. Chen GL, Vallender EJ, Miller GM. Functional characterization of the human TPH2 5' regulatory
region: untranslated region and polymorphisms modulate gene expression in vitro. Hum Genet.
2008;122(6):645-657. doi: 10.1007/s00439-007-0443-y.
66. Azmitia EC, Segal M. An autoradiographic analysis of the differential ascending projections of the
dorsal and median raphe nuclei in the rat. J Comp Neurol. 1978;179(3):641-667. doi:
10.1002/cne.901790311.
67. Nejati V, Khalaji S, Goodarzi H, Nitsche M. The role of ventromedial and dorsolateral prefrontal
cortex in attention and interpretation biases in individuals with general anxiety disorder (GAD): A tDCS
study. J Psychiatr Res. 2021;144:269-277. doi: 10.1016/j.jpsychires.2021.10.034.
68. Motzkin JC, Philippi CL, Wolf RC, Baskaya MK, Koenigs M. Ventromedial prefrontal cortex is critical
for the regulation of amygdala activity in humans. Biol Psychiatry. 2015;77(3):276-284. doi:
10.1016/j.biopsych.2014.02.014.
69. Williams LM. An integrative neuroscience model of "significance" processing. J Integr Neurosci.
2006;5(1):1-47. doi: 10.1142/s0219635206001082.
70. Etkin A, Büchel C, Gross JJ. The neural bases of emotion regulation. Nat Rev Neurosci.
2015;16(11):693-700. doi: 10.1038/nrn4044.
71. Via E, Fullana MA, Goldberg X, Tinoco-González D, Martínez-Zalacaín I, Soriano-Mas C, et al.
Ventromedial prefrontal cortex activity and pathological worry in generalised anxiety disorder. Br J
Psychiatry. 2018;213(1):437-443. doi: 10.1192/bjp.2018.65.
72. Marin MF, Zsido RG, Song H, Lasko NB, Killgore WDS, Rauch SL, et al. Skin Conductance Responses
and Neural Activations During Fear Conditioning and Extinction Recall Across Anxiety Disorders. JAMA
Psychiatry. 2017;74(6):622-631. doi: 10.1001/jamapsychiatry.2017.0329.
73. Hazlett-Stevens H, Borkovec TD. Interpretive cues and ambiguity in generalized anxiety disorder.
Behav Res Ther. 2004;42(8):881-892. doi: 10.1016/s0005-7967(03)00204-3.
74. Raij T, Nummenmaa A, Marin MF, Porter D, Furtak S, Setsompop K, et al. Prefrontal Cortex
Stimulation Enhances Fear Extinction Memory in Humans. Biol Psychiatry. 2018;84(2):129-137. doi:
10.1016/j.biopsych.2017.10.022.
75. Bar-Haim Y, Lamy D, Pergamin L, Bakermans-Kranenburg MJ, van IMH. Threat-related attentional
bias in anxious and nonanxious individuals: a meta-analytic study. Psychol Bull. 2007;133(1):1-24. doi:
10.1037/0033-2909.133.1.1.You can also read

















































